ВЉЮФ
ВњЧтСІдМЕШгкФъЧсЬЌЁО20еТЁП
||
Ек20еТ ВњЧтСІвтЮЖзХФъЧсЬЌ
БОбаОПРДздЖэТоЫЙЃЌЭЈЙ§БШНЯСНжжВЛЭЌРДдДЕФЖЏЮяФкдДадВњЧтЦјКЭВњМзЭщЕФВЛЭЌЃЌЗжЮіСЫСНжжЖЏЮядкЗўгУФбЯћЛЏЬМЫЎЛЏКЯЮяКѓЧтЦјКЭМзЭщЕФВњЩњВювьЃЌНсЙћЗЂЯжЃЌИпЧтЦјЖЏЮяФмВњЩњИќЖрЧтЦјЃЌВњМзЭщЖЏЮяВњЩњИќЖрМзЭщЁЃ
етвЛбаОПНсЙћжаЃЌИпЧтЦјзщЖЏЮяЕФЬиЕуИќЗћКЯФъЧсШЫЕФЬиЕуЃЌЖјИпМзЭщзщЖЏЮяИќНгНќРЯФъШЫЕФЬиЕуЁЃетвЛЯжЯѓШчЙћдкШЫРрШЗЪЕШчДЫЃЌОпгаЗЧГЃживЊЕФМлжЕЁЃЪзЯШЃЌбаОПЫЕУїГІЕРОњШКВњЩњЧтЦјЕФФмСІКЭФъСфЯрЙиЃЌФъСфдНДѓВњЧтЦјФмСІдНЕЭЃЌВњМзЭщФмСІдНИпЁЃЫфШЛМзЭщКЭЧтЦјЖМЗЂЯжОпгаПЙбѕЛЏФмСІЃЌЕЋЪЧЖдгкГІЕРШфЖЏЕФзїгУЪЧЭъШЋВЛЭЌЃЌЧтЦјДйНјГІЕРШфЖЏЃЌЖјМзЭщдђвжжЦГІЕРШфЖЏЁЃЫљвдЃЌЖдгквцЩњдЊРДЫЕЃЌРЯФъШЫвђЮЊЛсВњЩњИќЖрМзЭщЃЌЦфзїгУОЭБШНЯгаЯоЁЃЗўгУЧтЫЎдђВЛДцдкетжжгАЯьЃЌРЯФъШЫДйНјГІЕРШфЖЏЕФаЇЙћЛсИќКУЁЃЦфДЮЃЌЧтЦјВњЩњФмСІПЩКЭФъСфИКЯрЙиЃЌЕБШЛДцдкИіЬхВювьЃЌЛђепЫЕВњЧтЦјФмСІЧПЕФИќФъЧсЛђИќФъЧсЛЏЁЃЖдвЛаЉФкдДадВњЧтЦјФмСІВЛзуЕФРЯФъШЫРДЫЕЃЌЮЊФъЧсЛЏВЙГфЧтЦјИќгаМлжЕЁЃетаЉПДЗЈФПЧАШдШЛДцдкжїЙлвђЫиЃЌашвЊИќЖрбаОПжЄОнЁЃ
ЮЊСЫбщжЄЕЭМзЭщКЭИпМзЭщВњЩњепЖдВЛПЩЯћЛЏЬМЫЎЛЏКЯЮяЕФЗДгІЪЧЗёВЛЭЌЕФМйЩшЃЌНјааСЫвЛЯюбаОПЁЃдкжЎЧАЕФбаОПжаЃЌЮвУЧЗЂЯжPuschinoЗБжГГЁЃЈSPFзДЬЌЃЉЕФWistarДѓЪѓЪЧЕЭМзЭщВњЩњепЃЌЖјStolbovayaЗБжГГЁЕФГЃЙцДѓЪѓЪЧИпМзЭщВњЩњепЁЃдкУПИіЗБжГГЁЕФ25жЛДѓЪѓжаЃЌЦРЙРСЫЧтЦјКЭМзЭщКєЮќВтЪдвдМАГІЕРОњШКЕФЗжРрФкШнЁЃЪЙгУНіБЧВПзАжУДгЧхабЕФДѓЪѓФЧРяВЩМЏКєГіЕФПеЦјбљБОЃЌВЂЭЈЙ§ЦјЯрЩЋЦзЗЈНјааЗжЮіЁЃУПжЛДѓЪѓЕФГІЕРОњШКЗжРрФкШнЭЈЙ§16S rRNAЗНЗЈНјааЦРЙРЁЃШщЙћЬЧЁЂЙЯЖћНККЭОеЗлвдУПжмЪБМфМфИєЭЈЙ§ЙрЮИЗЈИјгшУПжЛДѓЪѓЁЃдкЬМЫЎЛЏКЯЮяИјвЉКѓЕФ8аЁЪБФкВтСПКєГіПеЦјбљБОжаЕФЧтЦјКЭМзЭщЫЎЦНЁЃВЛЭЌзщжЎМфЕФЗжРрЮЂЩњЮязщЙЙГЩВювьКмДѓЁЃЕЭМзЭщВњЩњДѓЪѓОпгаНЯЕЭЕФІСКЭІТЖрбљадЃЌChristensenellaceaeКЭAkkermansiaЯИОњЗсЖШНЯИпЃЌHelicobacteraceaeЗсЖШНЯЕЭЃЌЧвУЛгаMethanobacteriaceaeЃЌетгыаТЩњЖљКЭЖљЭЏЕФЮЂЩњЮязщЯрЫЦЁЃИпМзЭщВњЩњДѓЪѓЃЈРДздStobovayaЗБжГГЁЃЉЕФЮЂЩњЮяОњШКЖрбљадвЊИпЕУЖрЃЌЯёHelicobacteraceaeКЭMethanobacteriaceaeетбљЕФКФЧтЮЂЩњЮяЗсЖШИќИпЃЌНгНќРЯФъШЫЕФЮЂЩњЮязщГЩЁЃЕЭМзЭщВњЩњДѓЪѓЙрЮИЬМЫЎЛЏКЯЮяКѓЃЌжЛгаКєГіПеЦјжаЕФЧтЦјЫЎЦНдіМгЃЌЖјЯрЭЌЕФЬМЫЎЛЏКЯЮяжЛдкМзЭщЫЎЦНЩЯв§Ц№діМгЁЃЮвУЧЭЦВтЃЌЭтдДадЧтЦјЃЈИЛКЌЧтЦјЕФЫЎЃЉЕФИјвЉНЋИќгааЇЕиЬсИпРЯФъШЫЕФПЙбѕЛЏЗРгљФмСІЃЌвђЮЊЪГгУЪГЮяЯЫЮЌВЂВЛЛсЕМжТбЊвКжаЧтЦјЫЎЦНЕФдіМгЁЃ
НщЩм
дкПЦбЇНчЙуЗКЬжТлСЫГІЕРОњШКВЛНіВЮгыЮИГІМВВЁЕФЗЂВЁЛњжЦЁЊЁЊПЫТоЖїВЁЁЂГІвзМЄзлКЯжЂЁЂИЙаКЕШЃЌЛЙЩцМАЗЪХжжЂ[1]ЁЂЖЏТіжрбљгВЛЏ[2]ЁЂ2аЭЬЧФђВЁ[3]ЁЂаФСІЫЅНп[4]ЁЂЖЏТіадИпбЊбЙ[5]ЕШМВВЁЕФЗЂеЙжаЁЃКъЛљвђзщбЇбаОППЩвдМьВтЕНГІЕРЮЂЩњЮязщжаЕФДѓСПЮЂЩњЮяЃЌЕЋЫќУЧВЛФмЦРЙРУПИіМьВтЕНЕФЮЂЩњЮяЛђЦфШКЬхКЭШКТфЕФЙІФмЛђДњаЛЬиеїЁЃДњаЛзщбЇбаОПЃЌМДгЩОњШКВњЩњЕФЕЭЗжзгСПЩњЮяБъжОЮяЕФбаОПЃЌЮќв§СЫПЦбЇНчЕФДѓСПЙизЂЁЃетбљЕФЩњЮяБъжОЮяАќРЈЦјЬхЃЌШчЧтЦјЁЂМзЭщЁЂСђЛЏЧтЃЌвдМАЖЬСДжЌЗОЫсЃЈSCFAЃЉКЭЦфЫћЕЭЗжзгСПЛгЗЂадЮяжЪ[6]ЁЃвбОБэУїЃЌЧтЦјЪЧгЩГІЕРОњШКзЈУХВњЩњЕФЃЌЖјВЛЪЧЫожїЕФЬхЯИАћ[7, 8]ЁЃГЄЪБМфвдРДЃЌЩњЮяМзЭщЕФаЮГЩБЛШЯЮЊЪЧжЛгаВњМзЭщЙХОњВХФмВњЩњЕФЁЃШЛЖјЃЌНќФъРДдкЬхЭтЪЕбщжаБэУїЃЌдкШБбѕЬѕМўЯТЃЌШЫРрЯИАћЯЕвВФмВњЩњМзЭщ[9]ЁЃМИКѕ70%СаШыШЫЬхЮЂЩњЮязщЯюФПЮИГІЕРЃЈHMP GIЃЉВЮПМЛљвђзщЪ§ОнПтЕФЮЂЩњЮяЮяжжБрТыСЫЧтЛЏУИЃЌетЖдгкеЙЪОВњЧтЃЈВњЩњЧтЦјЃЉЛђКФЧтЃЈЯћКФЧтЦјЃЉЛюадЪЧБиашЕФ[10]ЁЃМьВщЮИГІЕРжаЧтЦјВњЩњЕФзюГѕВтЪджЎвЛЪЧШщЙћЬЧКєЮќВтЪдЁЃШщЙћЬЧЪЧШЫРрВЛЯћЛЏЕФЬМЫЎЛЏКЯЮяЁЃЕБИјгшШЫРрЛђФіГнРрЖЏЮяЪБЃЌШщЙћЬЧНЋДЉЙ§ГІЕРЃЌжБЕНЕНДяФмЙЛЯћЛЏИУЕзЮяЕФГІЕРОњШКПтЁЃзїЮЊЗЂНЭЕФИБВњЮяЃЌВњЩњСЫЧтЦјЁЃЧтЦјЃЈвдМАЦфЫћГІЕРВњЩњЕФЦјЬхЃЉНјШыбЊвКЃЌбЛЗЃЌРЉЩЂЕНЗЮХнПеМфЃЌВЂдкКєЮќжаКєГі[11]ЁЃ
ИљОнЖде§ГЃЪмЪдепЕФбаОПЃЌЦЈЕФжївЊГЩЗжАќРЈЃКЧтЦјЃЈ3-20%ЃЉЁЂЖўбѕЛЏЬМЃЈ9-14%ЃЉЁЂМзЭщЃЈ7.2%ЃЉЁЂбѕЦјЃЈ2-4%ЃЉКЭСђЛЏЧтЃЈ0.00028%ЃЉ[12, 13]ЁЃГІЕРЮЂЩњЮяШКжаЕФЙХОњЛсДњаЛЧтЦјЁЃСНжжжївЊЕФМзЭщВњЩњЮяжжЁЊЁЊMethanobrevibacter smithiiКЭMethanosphaera stadtmanaeдкШЫРрНсГІжаЪЧжївЊВњМзЭщОњЁЃЫќУЧЕФЪ§СПбизХНсГІдіМгЃЌжБЕНдкжБГІДяЕНзюДѓжЕЁЃMethanobrevibacter smithiiРћгУЧтЦјНЋЖўбѕЛЏЬМЛЙдЮЊМзЭщЃЌЖјMethanosphaera stadtmanaeРћгУЧтЦјНЋМзДМЛЙдЮЊМзЭщЁЃКЯГЩвЛЗжзгМзЭщашвЊЫФИіЧтЗжзгКЭвЛИіЖўбѕЛЏЬМЗжзгЃКCO2 + 4H2 Ёњ CH4 + 2H2O [14, 15]ЁЃ
жБЕН2007ФъЃЌЧтЦј(ЧтЦј)БЛШЯЮЊЪЧЩњРэЩЯЖшадЕФЦјЬхЁЃЬЋЬяГЩФаНЬЪкМАЦфЭЌЪТдкЁЖздШЛЁЗдгжОЩЯЗЂБэСЫЙиМќадЮФеТЃЌжЄУїСЫЧтЦјЕФПЙбѕЛЏЛюад[16]ЁЃдкЙ§ШЅЕФЖўЪЎФъРяЃЌРДздСйДВЧАКЭСйДВбаОПЕФЛ§РлжЄОнБэУїЃЌЧтЦјПЩФмзїЮЊвЛжжПЙбѕЛЏМСЃЌЖдАќРЈДњаЛадМВВЁдкФкЕФИїжжМВВЁЗЂЛгжЮСЦКЭдЄЗРзїгУ[17]ЁЃДЫЭтЃЌЛёЕУЕФжЄОнЯдЪОЃЌЧтЦјгАЯьаХКХЭЈТЗЃЌЭЈЙ§етаЉЭЈТЗаХЯЂДЋЕнДЉЙ§ЯИАћФЄЃЌВЂЧвЛЙОпгаЯИАћБЃЛЄзїгУЃЌМѕЩйДйбзЯИАћвђзгЕФКЯГЩКЭЯИАћЕђЭіЁЃЫцКѓНјааЕФСйДВЪдбщдкКмДѓГЬЖШЩЯжЄЪЕСЫжЎЧАдкЖЏЮяЩэЩЯЪЕбщбаОПЕФНсЙћЃЌДгЖјШЗШЯСЫЧтЦјЕФаФдрБЃЛЄКЭЩёОБЃЛЄаЇЙћЃЌвдМАЖдФкЦЄЙІФмеЯАЕФЛ§МЋгАЯь[18, 19]ЁЃ
вЛЗНУцЃЌЧтЦјЛ§МЋаЇгІЕФЗЂЯжЕМжТСЫбаОПЭтдДадЧтЦјаЇгІЕФбаОПЪ§СПдіМгЃЛСэвЛЗНУцЃЌвВжиаТМЄЗЂСЫЖдГІЕРЮЂЩњЮяШКВњЩњЕФФкдДадЧтЦјаЇгІЗжЮіЕФаЫШЄЁЃ
МИЯюбаОПБэУїЃЌНЁПЕИіЬхПЩвдИљОнЬМЫЎЛЏКЯЮяв§ЗЂЕФЧтЦјКЭМзЭщВњСПБЛЗжЮЊШ§ИіЬѕМўбЧзщЁЃЕквЛзщдкШщЙћЬЧИјвЉКѓКєГіЕФПеЦјжаЧтЦјКЭМзЭщЫЎЦННЯЕЭЁЃЕкЖўзщЃЌМзЭщбєадЃЌМзЭщдіМгЖјЧтЦјМИКѕУЛгадіМгЁЃЕкШ§зщжЛЙлВьЕНЧтЦјЫЎЦНЕФдіМг[20, 21]ЁЃЪЙгУВЛЭЌНсЙЙЕФВЛПЩЯћЛЏЩХЪГЯЫЮЌзїЮЊвцЩњдЊЪЧвЛжжАВШЋгааЇЕФЗНЗЈЃЌПЩвдИФЩЦЮЂЩњЮяШКЕФЙІФмзДЬЌЃЌвђЮЊЫќУЧЪЧВњЩњЧтЦјЮЂЩњЮяЕФжївЊЕзЮя[22,23,24]ЁЃ
вђДЫЃЌЮвУЧЕФбаОПжМдкЕїВщЪЧЗёПЩвдЭЈЙ§ЪЙгУВЛЭЌЕФВЛПЩЯћЛЏЬМЫЎЛЏКЯЮяРДдіМгЕЭВњМзЭщКЭИпВњМзЭщДѓЪѓЕФФкдДадЧтЦјВњСПЃЌДгЖјЬсИпЛњЬхЕФПЙбѕЛЏЗРгљФмСІЁЃ
ВФСЯКЭЗНЗЈ
ЪЕбщЖЏЮя
ИљОнХЗжовщЛсКЭХЗжоСЊУЫРэЪТЛсЙигкБЃЛЄгУгкПЦбЇФПЕФЖЏЮяЕФ2010/63/EUжИСюЃЌЖдЖЏЮяЕФЫЧбјКЭЫљгаВйзїОљАДЙцЖЈжДааЁЃЪЕбщЗНАИвбгЩФЊЫЙПЦЙњСЂДѓбЇТзРэЮЏдБЛсХњзМЃЈЩъЧыБрКХ129-ZhЃЌШеЦкЮЊ2021Фъ5дТ31ШеЃЉЁЃЖЏЮяРДздСНИіВЛЭЌЕФбјжГГЁЁЃЕквЛХњЃЌ25жЛДѓЪѓЁЊЁЊРДздЁАЦеЯЃХЕЁБЪЕбщЪвЖЏЮябјжГГЁЃЈЩњЮягаЛњЛЏбЇПЦбЇбаОПЫљЃЌФЊЫЙПЦЕиЧјЃЉЁЃИљОнжЄЪщЃЌWistarДѓЪѓОпгаSPFзДЬЌЁЃЕкЖўХњЃЌ25жЛДѓЪѓЁЊЁЊРДздЪЕбщЪвЖЏЮяЗжВПЁАЫЙЭаЖћВЉЭпбЧЁБЃЈЖэТоЫЙСЊАювНСЦЩњЮябЇОжЃЌФЊЫЙПЦЧјЃЉЁЃетаЉДѓЪѓЗћКЯвХДЋБъзМЃЌетвЛЕуЭЈЙ§жЪСПжЄЪщЕУЕНСЫШЗШЯЁЃ
ЖЏЮяЕНДяКѓСЂМДБЛЗХжУдкЖЏЮяЗПФкЃЌМьВщЪЧЗёгаЩЫПкЁЂЬлЭДКЭ/ЛђВЛЪЪЕФМЃЯѓЃЌВЂБЛАВжУдкT3С§жаЃЌУПС§2жЛДѓЪѓЁЃЖЏЮядкБъзМЬѕМўЯТЫЧбјЃЌЙтееЪБМфЮЊ12аЁЪБЃЌПЩЮоЯоСПЛёШЁЫЎКЭЪГЮяЃЌВЂЧвЖЈЦкПижЦЪвФкЮТЖШКЭЪЊЖШЁЃЪЪгІЖЏЮяЗПЬѕМўЕФЪБМфжСЩйЮЊ7ЬьЁЃЮЊСЫзюМбЪЕЪЉЪЕбщВйзїВЂБмУтбЙСІЖдбаОПНсЙћЕФгАЯьЃЌЫљгаЖЏЮяЖМОЙ§СЫДІРэГЬађЁЃ
ЮИФкИјвЉ
ЫљгаЕїВщЮяжЪОљЭЈЙ§ЙрЮИЗЈЪЙгУЮоОњЮИФкЙмЃЈГпДч16ЃЌЪЪгУгк200-300ПЫжиЕФДѓЪѓЃЉИјвЉЁЃЖдгкУПИіЖЏЮязщЃЌЁАЦеЯЃХЕЁБКЭЁАЫЙЭаЖћВЉЭпбЧЁБЃЌЖМЪЙгУСЫЕЅЖРЕФЮИФкЙмЃЌвдОЁСПМѕЩйЮЂЩњЮяШКдкзщМфЕФзЊвЦЁЃ
гУгкЧтЦј/МзЭщКєЦјЪдбщЕФвћЪГЛЏКЯЮя
ШщЙћЬЧЃЈFresenius Kabi IPSUM S.r.l.ЃЌвтДѓРћЃЉЁЃШщЙћЬЧЃЈ2ПЫ/ЙЋНяЬхжиЃЌЭЈЙ§ЙрЮИЃЉЪЧвЛжжЖўЬЧЃЌгЩАыШщЬЧКЭЙћЬЧВаЛљзщГЩЁЃ
PHGGЃЈВПЗжЫЎНтЙЯЖћНКЃЌ4ПЫ/ЙЋНяЃЌOptifiber ®ЃЌШИГВНЁПЕПЦбЇЃЌЕТЙњЃЉЁЃЦфжївЊГЩЗжЪЧДЬЮрЭЉЙћЪЕЬсШЁЮяЃЌМДЙЯЖћНКЁЃЛЏбЇНсЙЙЪЧ100%ЙЯЖћАыШщИЪТЖОлЬЧЃЌгЩИЪТЖЬЧКЭАыШщЬЧвдДѓдМ2:1ЕФБШР§зщГЩЁЃ
ОеЗлЃЈЕААзЙЋЫОЃЌЖэТоЫЙЃЉЁЃОеЗлЪЧвЛжжD-ЙћЬЧЕФОлКЯЮяЃЌгЩ30-35ИіпЛрЋЬЧаЮЪНЕФЙћЬЧВаЛљзщГЩЁЃОеЗлвд4ПЫ/ЙЋНяМССПИјвЉЁЃ
ИјвЉЮяжЪЕФжиСПЪЧИљОнЖЏЮяЕФЬхжиМЦЫуЕФЁЃЫљгаЮяжЪдкИјвЉЧАЖМШмНтдк2-3КСЩ§ДПОЛЫЎжаЁЃУПжжРраЭЕФВЛПЩЯћЛЏЬМЫЎЛЏКЯЮяЖМдкСНзщЕФУПжЛДѓЪѓЩЯНјааСЫВтЪдЁЃУПжЛДѓЪѓЕФСЌајВтСПЪБМфМфИєЮЊвЛжмЃЌвдХХГ§ЯШЧАВтЪдЕФгАЯьЁЃ
КєГіЦјЬхбљЦЗВЩМЏЪЕбщЩшМЦ
ЪЕбщзАжУВЩгУСЫЩњУќжЇГжЯЕЭГЕФИФНјАцБОЃЌгУгкЧтЦјЕФЖЈСПЪеМЏ[25]ЃЌЕЋЮвУЧУЛгаНЋДѓЪѓЗХжУдкУмЗтЕФЖЏЮяЪвжаЃЌЖјЪЧЪЙгУСЫвЛжжДјгаAllay™дМЪјСьЕФRat BreathalyzerЯЕЭГ[26]ЕФАцБОЃЌвдБугкВЩМЏКєГіЦјЬхбљБОЃЌВтСПЧтЦјКЭМзЭщЕФЫЎЦН[27]ЃЈМћЭМ20.1ЃЉЁЃдкБОбаОПжаЃЌAllay™дМЪјСьгыЭЈЗчЕФНіЯоБЧзгЕФЧЛЪвНсКЯЪЙгУЁЃИУзАжУЪЧгЩКЯжјепжЎвЛЃЈBondarenko G.ЃЉЙЙНЈЕФЁЃ
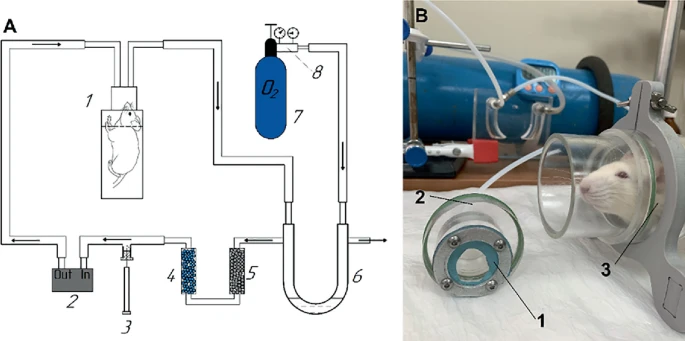
ЭМ20.1ЪЕбщзАжУЪОвтЭМЃЌгУгкЪеМЏЪЕбщжаШЋВППеЦјбљБОЁЃ1ЁЊЦјЦПЯфЃЌ2ЁЊПеЦјБУЃЌ3ЁЊВЃСЇзЂЩфЦїЃЌ4ЁЊЮќЪЊМСЃЈЫЎФ§НКЃЉЃЌ5ЁЊЖўбѕЛЏЬМЮќЪеМСЃЈЫеДђЪЏЛвЃЉЃЌ6ЁЊбЙСІМЦЃЌ7ЁЊбѕЦјЦПЃЌ8ЁЊЦјЬхМѕбЙЗЇЃЛB ЖЏЮяС§ЕФееЦЌЃК1ЁЊСНИіЧЛЪвЗжИєЦНУцЩЯЕФЯ№НКФЄПзЃЌ2ЁЊЖЏЮяЭЗВПЗХжУЧЛЪвЃЌгУгкЖЏЮяжиИДКєЮќЃЌ3ЁЊЖЏЮяЩэЬхЗХжУЧЛЪвЃЌгУгкЙЬЖЈЖЏЮяЕФОБВПЧјгђ
ЦјЬхЪЕбщзАжУгЩШ§ИіЙиМќВПЗжзщГЩЁЊЁЊЦјЦПЯфЁЂЩњУќжЇГжЯЕЭГКЭбЛЗЭЈЗчЯЕЭГЁЃЖЏЮяЕФЭЗВПгУОБВПЫмСЯМаЙЬЖЈЃЌНіДгЩЯЗНКЭВрУцНєУмЬљКЯОБВПЧјгђЃЌетдЪаэЖЏЮяздгЩКєЮќЁЃДѓЪѓЕФБЧзгЮЛгквЛИіЬиЪтЕФЭЗВПИєМфФкЃЌжмЮЇНєУмЬљКЯЯ№НКФЄЁЃетбљЃЌзАжУЕФУмЗтадЕУЕНБЃжЄЃЌКєГіЕФПеЦјдкЯЕЭГФкВПбЛЗЃЌВЛЛсвнГіЁЃЭЗВПИєМфФкгаЪфШыКЭЪфГіСЌНгЦїЁЃ
ЭЈЙ§Й§ТЫЦїбЛЗЕФКєГіПеЦјБЛЧхГ§СЫЫЎЗжКЭЖўбѕЛЏЬМЁЃбѕЦјЫЎЦНЮЌГждк20-21%ЁЃетбљЃЌЧтЦјКЭМзЭщЕФЛ§РлОЭЗЂЩњСЫЃЌетЖдгкВтСПЦјЬхзАжУЯЕЭГжаЕФЦјЬхХЈЖШЪЧБивЊЕФЁЃНјЦјЭЈЙ§УмЗтЗЇНјааЃЌЪЙгУЬиЪтЕФЦјУмаЭHamiltonзЂЩфЦїНјааЦјЯрЩЋЦзЗжЮіЁЃЮвУЧЯЕЭГЕФгХЪЦдкгкЃЌЮвУЧФмЙЛЪеМЏЕНЮДЪмеЭЦјИЩШХЕФНіКєГіЕФПеЦјЃЌетЪЙЮвУЧЕФЯЕЭГРрЫЦгкШЫРрКєЮќВтЪджаЕФВЩбљПеЦјЁЃ
ЪзДЮПеЦјбљБОЪЧдкЖЏЮяНћЪГ12аЁЪБКѓВЩМЏЕФЃЌЕБЪБДѓЪѓПЩвдздгЩвћЫЎЁЃШЛКѓЃЌНЋЖЏЮяДгС§згжаШЁГіЃЌВЂЭЈЙ§ЙрЮИЗЈИјгшЬиЖЈЬМЫЎЛЏКЯЮяШмвКЁЃдкУПжЛДѓЪѓжаЃЌКєГіПеЦјбљБОЙВЪеМЏЮхДЮЃКЛљЯпЁЂЬМЫЎЛЏКЯЮяИјвЉКѓ2ЁЂ4ЁЂ6КЭ8аЁЪБЁЃУПДЮВЩбљКѓЃЌДѓЪѓБЛЫЭЛиЦфС§жаЃЌжБЕНЯТвЛИіЪБМфЕуЁЃЮЊСЫХХГ§ВЛЭЌФбЯћЛЏЬМЫЎЛЏКЯЮядкГІЕРЮЂЩњЮяШКжаЕФНЛВцзїгУЃЌДѓЪѓдкгУСэвЛжжФбЯћЛЏЬМЫЎЛЏКЯЮяНјааВтСПЧАЃЌЛсдкБъзМвћЪГжаБЃГжМИЬьЁЃ
бљБОЗжЮі
дкЦјЬхЩЋЦзвЧЃЈTRILyzer mBA-3000ЃЌШеБОЃЉЩЯНјааСЫКєГіПеЦјжаЧтЦјЁЂМзЭщКЭвЛбѕЛЏЬМЕФВтЖЈЃЌЪЙгУКЌгаЕЭХЈЖШЃЈПеЦјжаЧтЦјЁЂМзЭщКЭвЛбѕЛЏЬМЃЌУПжж5ppmЃЉКЭИпХЈЖШЃЈЧтЦјЁЂМзЭщКЭвЛбѕЛЏЬМЁЊПеЦјжаУПжж50ppmЃЉЕФЦјЬхНјаааЃзМЁЃ
ЛцжЦСЫЧтЦјКЭМзЭщЕФХЈЖШ-ЪБМфЧњЯпЃЌВЂМЦЫуСЫAUC 0-8аЁЪБЧњЯпЯТЕФУцЛ§ЃЈppm*hЃЉЃЌетЗДгГСЫЪЕбщ8аЁЪБФкЦјЬхЩњГЩЕФЫйТЪЁЃ
ЗрБубљБОЪеМЏ
ЗрБубљБОЪеМЏдкЕЅИіEppendorfЙмжаЁЃEppendorfЙмСЂМДИЧЩЯИЧзгЃЌБъМЧВЂРфЖГдк-80ЁуCЁЃЖдгкУПжЛЖЏЮяЃЌвЛДЮШЁСНИіЗрБубљБОЁЃШЁбљЪЧдкКєЮќВтЪдЧАЯІНјааЕФЁЃ
ЮЂЩњЮяШКТфзщГЩЗжЮі
ЪЙгУQIAampPowerFecal Pro DNA KitЃЈQiagenЃЌHildenЃЌЕТЙњЃЉАДеежЦдьЩЬЕФЫЕУїДгЗрБужаЗжРыDNAЁЃЭЈЙ§гЋЙтМЦЃЈThermo Fisher ScientificЃЌMAЃЌУРЙњЃЉЦРЙРDNAЕФЪ§СПКЭХЈЖШЁЃЪЙгУ1-2 ngЕФDNAНјааЮФПтзМБИЁЃЪЙгУв§Юя515F [28]КЭPro-mod-805R [29]ЭЈЙ§СНВНPCRжЦБИ16S rRNA V4ИпБфЧјРЉдізгЮФПтЁЃ
УПИіDNAбљБОЕФPCRЖМЪЧИљОнжЎЧАЕФавщ[30]НјааЕФЁЃдкVeritiШШбЛЗвЧЃЈApplied BiosystemsЃЌMAЃЌУРЙњЃЉЩЯНјааРЉдіЁЃЕУЕНЕФЮФПтдкЧэжЌЬЧФ§НКЩЯМьВщВЂЕШФІЖћЛьКЯЁЃзюжеЛьКЯЮяЪЙгУQIAquick Gel Extraction KitЃЈQiagenЃЌЕТЙњЃЉАДеежЦдьЩЬЕФавщДПЛЏЁЃ
ЪЙгУИіШЫВтађЯЕЭГЃЈIlluminaЃЌSan DiegoЃЌCAЃЌУРЙњЃЉНјааВтађЃЌ2ЁС156bpХфЖдФЉЖЫЁЃ
ЭГМЦбЇЗжЮі
ЪЕбщНсЙћЕФЭГМЦЗжЮіКЭЫцЛњЗжзщЪЧЪЙгУGraphPad Prism 8ШэМўЭъГЩЕФЁЃЪЙгУShapiro-WilkБъзММьбщЗжВМЕФе§ЬЌадЁЃЖдгкЦЕТЪГЌЙ§3ИіВтСПЕувдЩЯЕФКєЮќВтЪдНсЙћЃЌВЩгУЧњЯпЯТУцЛ§ЃЈAUCЃЉМЦЫуНјааБШНЯЁЃЪЙгУЕЅвђЫиЕЅЗНЯђЗНВюЗжЮіЃЈANOVAЃЉБШНЯЖргкСНИібљБОжаЕЅИіжИЪ§ЕФЦНОљжЕЁЃЮЊСЫШЗЖЈзщБ№КЭБЉТЖГжајЪБМфЕФЭЌЪБаЇгІЃЌвдМАетаЉвђЫижЎМфЕФЯрЛЅзїгУЃЌЪЙгУЫЋвђЫиANOVAЗНВюЗжЮіРДЪЖБ№зщМфВювьЃЌЭЈЙ§ХфЖдКЭЗЧХфЖдtМьбщЗжБ№ЗжЮіЯрЙибљБОКЭЖРСЂбљБОЁЃЖдгкЗЧе§ЬЌЗжВМЕФзщМфГЩЖдБШНЯЃЌЪЙгУWilcoxonМьбщЗжЮіЯрЙибљБОЃЌMann-WhitneyМьбщЗжЮіЖРСЂбљБОЁЃЪЙгУSpearmanЕШМЖЯрЙиЯЕЪ§МЦЫуЯрЙиадЁЃЪЙгУROUTБъзМХХГ§ЭГМЦвьГЃжЕЃЌQжЕВЛГЌЙ§1%ЁЃдкp < 0.05ЪБЃЌШЯЮЊВювьОпгаЭГМЦбЇвтвхЁЃЫљгаЪ§ОнвдОљжЕЁРБъзМВюЃЈMeanЁРSDЃЉГЪЯжЁЃЮЂЩњЮяШКТфзщГЩЁЂalphaКЭbetaЖрбљаджИБъКЭЭГМЦЪЙгУRЃЈАцБО4.2.0ЃЉНјааЗжЮіЁЃИпжЪСПЖСШЁЖдЪЙгУDADA2ЙмЕР[29]ДІРэЃЌИљОнвбЗЂВМЕФавщ[31]жДааЁЃЪЙгУSilva138Ъ§ОнПт[32]ЃЌЭЈЙ§ЦгЫиБДвЖЫЙЗжРрЦїШЗЖЈРЉдіађСаБфЬхЃЈASVsЃЉЕФЗжРрЁЃЮЊСЫНјвЛВНЗжЮіЃЌЪЙгУPhyloseqАќ[33]ЖдЖСШЁНјааЯЁБЁЛЏДІРэЁЃЛёЕУЕФASVВЮПМађСаЁЂбљБОдЊЪ§ОнЁЂЗсЖШБэКЭЗжРрЕМШыЕНPhyloseqАќжаЃЌЫљгаКѓајВйзїЖМдкPhyloseqЖдЯѓЩЯжДааЁЃЪЙгУmicroecoАќЃЈЛљгкОрРыОиеѓЕФЖрБфСПЗНВюЗжЮіЃЉ[34]КЭveganАќ[35]НјааЮЂЩњЮяШКТфзщГЩЕФПЩЪгЛЏЁЃ
НсЙћ
ВЛЭЌНсЙЙЕЅвЛЬМЫЎЛЏКЯЮяИКдиЃЈШщЙћЬЧЁЂОеЗлЁЂPHGGЯЫЮЌЃЉЖдДѓЪѓКєГіЦјжаЦјЬхБъМЧЮяЃЈЧтЦјКЭМзЭщЃЉКЌСПЕФгАЯь
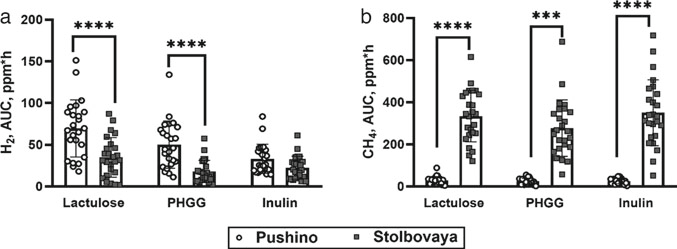
дкЁАPuschinoЁБЪЕбщзщжаЃЌЪЙгУШщЙћЬЧШмвККЭPHGGЯЫЮЌЪБЃЌЧтЦјКЭМзЭщЫЎЦНгаЯджјВювьЃЌp < 0.0001КЭp = 0.001ЃЌЗжБ№ЃЈЭМ20.2aЃЉЁЃдкОеЗлзщжаЃЌЧтЦјКЭМзЭщЕФЫЎЦНДѓжТЯрЭЌЃЈдМ35-40 ppm*hЃЉЃЌВювьВЛЯджјЃЌp = 0.6ЃЈЭМ20.2aЃЉЁЃдкЁАStolbovayaЁБЪЕбщзщжаБШНЯЧтЦјКЭМзЭщЫЎЦНЪБЃЌЫљгаЪЕбщзщжаЦјЬхДњаЛЮяЫЎЦНЕФВювьОљДяЕНИпЖШЯджјадЃЌp < 0.0001ЃЈЭМ20.2bЃЉЁЃЦНОљЖјбдЃЌЫљгаЪЕбщзщжаМзЭщЫЎЦНБШЧтЦјЫЎЦНИпГі12БЖЃЌЧтЦјЕФзмЦНОљЫЎЦНЮЊ25 ЁР 8 ppm*hЃЈШщЙћЬЧ35 ЁР 23 ppm*hЃЌPHGG 18 ЁР 13 ppm*hЃЌОеЗл22 ЁР 13 ppm*hЃЉЃЌМзЭщЕФЦНОљЫЎЦНЮЊ320 ЁР 38 ppm*hЃЈШщЙћЬЧ334 ЁР 122 ppm*hЃЌPHGG 276 ЁР 134 ppm*hЃЌОеЗл350 ЁР 156 ppm*hЃЉЃЈЭМ20.2bЃЉЁЃ

ЭМ20.2 ЯдЪОСЫдкЁАPuschinoЁБзщКЭЁАStolbovayaЁБзщЕФДѓЪѓжаЃЌЕЅДЮЙрЮИВЛЭЌНсЙЙЗЧЯћЛЏадЬМЫЎЛЏКЯЮяЃЈШщЙћЬЧЁЂPHGGКЭОеЗлЃЉКѓКєГіЦјжаЧтЦјКЭМзЭщЕФБШНЯЁЃ
ЕББШНЯзюГѕОпгаВЛЭЌЧтЦјВњЩњЛюадЮЂЩњЮяШКЕФДѓЪѓЪБЃЌЗЂЯжетаЉДѓЪѓЖдФГаЉЬМЫЎЛЏКЯЮяЕФЧтЦјЗДгІВЛЭЌЁЃвђДЫЃЌдкЁАPuschinoЁБзщЕФДѓЪѓжаЃЌгыЁАStolbovayaЁБзщЕФДѓЪѓЯрБШЃЌШщЙћЬЧКЭPHGGзщжаЧтЦјВњЩњЕФЯджјдіМгЗжБ№ЮЊ1.9БЖКЭ2.7БЖЃЌp < 0.0001ЃЈЭМ20.3aЃЉЁЃЖдгкзюГѕОпгаИпЧтЧБСІЕФДѓЪѓРДЫЕЃЌОеЗлЕФЗДгІБШЁАStolbovayaЁБзщЕФДѓЪѓИпГі1.4БЖЃЌЕЋетИіВювьУЛгаЭГМЦбЇвтвхЃЌp = 0.2ЃЈЭМ20.3aЃЉЁЃ
ЖдгкзюГѕОпгаВЛЭЌМзЭщЩњГЩЛюадЛљЯпЕФДѓЪѓзщЃЌЕЅДЮЙрЮИЗЧЯћЛЏадЬМЫЎЛЏКЯЮяЕФМзЭщЗДгІвВгаЫљВЛЭЌЁЃдкЁАStolbovayaЁБзщжаЃЌгыЁАPuschinoЁБзщЕФДѓЪѓЯрБШЃЌЕЅДЮгІгУЗЧЯћЛЏадЬМЫЎЛЏКЯЮяКѓЕФМзЭщЫЎЦНИпГі11.5БЖЛђИќЖрЃЈЪЙгУШщЙћЬЧИпГі12БЖЃЌPHGGИпГі11.5БЖЃЌОеЗлИпГі14БЖЃЉЃЌp < 0.0001ЃЈЭМ20.3bЃЉЁЃ
ЭМ20.3 БШНЯСЫЁАPuschinoЁБКЭЁАStolbovayaЁБзщЕФДѓЪѓдкЕЅДЮЙрЮИВЛЭЌЗЧЯћЛЏадЬМЫЎЛЏКЯЮяКѓКєГіЦјжаЕФa ЧтЦјКЭb МзЭщЫЎЦНЁЃ
ЫљгазюГѕОпгаЛљДЁЧтЧБСІЧвУЛгаУїЯдМзЭщЩњГЩЛюадЕФЁАPuschinoЁБзщДѓЪѓЃЌЖдЫљгаРраЭЖюЭтвћЪГЛЏКЯЮяЕФЕЅДЮгІгУЖМгаЧтЦјЗДгІЃЈЭМ20.4ЃЉЁЃЧтЦјВњЩњЕФдіМгАДвдЯТЫГађЃКОеЗл Ёњ PHGG Ёњ ШщЙћЬЧЁЃМЧТМЕНЕФзюДѓЧтЦјВњЩњЪЧдкзЂЩфШщЙћЬЧЕФЧщПіЯТЁЃЫљгазщжЎМфЕФВювьОљОпгаЭГМЦбЇвтвхЃЌPHGGгыОеЗлзщжЎМфЕФЯджјадЫЎЦНЮЊp = 0.03ЃЌШщЙћЬЧгыPHGGзщжЎМфЮЊp = 0.009ЃЌШщЙћЬЧгыОеЗлзщжЎМфЮЊp < 0.0001ЃЈЭМ20.4aЃЉЁЃЭЌЪБЃЌдкЪЙгУЫљгаЗЧЯћЛЏадЬМЫЎЛЏКЯЮяЪБЃЌМШУЛгаМЧТМЕНЯрЖдгкЁАПеИЙЁБзДЬЌЃЈСуВтСПЕуЃЉЕФМзЭщЫЎЦНЕФЯджјдіМгЃЌвВУЛгаМЧТМЕНКєГіЦјжаМзЭщЫЎЦНжЎМфЕФзщМфВювьЃЌЫљгазщжЎМфЕФЯджјадЫЎЦНp > 0.9ЃЈЭМ20.4bЃЉЁЃ
дкЁАStolbovayaЁБзщЕФДѓЪѓжаЃЌМЧТМЕНСЫИпЕФМзЭщЩњГЩЛюадЃЈЭМ20.4bЃЉКЭЕЭЕФЧтЧБСІЃЈЭМ20.4aЃЉЁЃЕБЪЙгУгыЁАPuschinoЁБзщДѓЪѓЯрЭЌЕФЗЧЯћЛЏадЬМЫЎЛЏКЯЮяЪБЃЌДѓЪѓЕФЗДгІЭъШЋЯрЗДЁЃдкШЮКЮЪЕбщзщжаЖМУЛгаМЧТМЕНЧтЦјЕФдіМгЃЌЖјЧвЧтЦјЕФЛљЯпЫЎЦНвВЗЧГЃЕЭЃЌp > 0.9ЃЈЭМ20.4aЃЉЁЃ
ЖдЫљгаШ§ИізщЕФЯЫЮЌгІгУЕФМзЭщЩњГЩЗДгІЖМКмИпЃЌВЂЧвдкPHGG Ёњ ШщЙћЬЧ Ёњ ОеЗлЯЕСажавЛжТдіМгЁЃжЛдкPHGGКЭОеЗлзщжЎМфЗЂЯжСЫЭГМЦбЇЩЯЕФЯджјВювьЃЌp = 0.03ЁЃШщЙћЬЧгыPHGGзщжЎМфвдМАШщЙћЬЧгыОеЗлзщжЎМфЕФВювьЗжБ№ЮЊp = 0.1КЭp = 0.9ЃЈЭМ20.4bЃЉЁЃ

ЭМ20.4 ЯдЪОСЫдкЁАPuschinoЁБКЭЁАStolbovayaЁБзщЕФДѓЪѓжаЃЌЕЅДЮЪЙгУВЛЭЌЗЧЯћЛЏадЬМЫЎЛЏКЯЮяКѓКєГіЦјжаЕФa ЧтЦјКЭb МзЭщЫЎЦНБШНЯЁЃ
ГІЕРЮЂЩњЮяШКзщГЩЗжЮі
дкШЅИДгУКЭЖСШЁдЄДІРэжЎКѓЃЌУПИібљБОжиИДЦНОљЛёЕУСЫ19,201ИіЖСШЁЖдЁЃОЙ§DADA2ДІРэВНжшЃЈЖюЭтЙ§ТЫЁЂШЅдыЁЂКЯВЂКЭЧЖКЯЬхвЦГ§ЃЉКѓЃЌУПИіжиИДЕФЦНОљЖСШЁМЦЪ§ЮЊ15,017ЁЃЯЁЛЏЗжЮіЯдЪОЫљгаВтађбљБОЖМОпгаСМКУЕФБЅКЭЖШЃЌетБэУїВтађЩюЖШзуЙЛЁЃDADA2СїГЬВњЩњСЫ1854ИіРЉдіађСаБфЬхЃЈASVsЃЉЁЃ
ЮЂЩњЮяЖрбљадЗжЮі
ЭЈЙ§ЮЂЩњЮязщбЇАќНјааЕФalphaЖрбљаджИБъЗжЮіЯдЪОЃЌЙлВьЕНЕФASVЪ§СПЃЌвдМАShannonКЭФцЯђSimpsonжИЪ§ЃЌдкЁАStolbovayaЁБзщжаЯджјИпгкЁАPuschinoЁБзщЃЈp < 0.001ЃЉЃЈЭМ20.5AЃЉЁЃЭЈЙ§veganАќЕФadonis2КЏЪ§ЪЙгУBray-CurtisОрРыНјааЕФbetaЖрбљадЗжЮіЯдЪОЃЌЁАPuschinoЁБКЭЁАStolbovayaЁБзщЕФЮЂЩњЮяШКТфБЫДЫЯджјВЛЭЌЃЈ10,000ДЮжУЛЛЃЛF1, 96 = 88.36ЃЌp = 0.001ЃЉЃЌетЭЈЙ§PCoAЗжЮівВЕУЕНСЫЧхЮњЕФжЇГжЃЈЭМ20.5BЃЉЁЃдк1854ИіASVsжаЃЌга717ИіЪЧЁАPuschinoЁБзщЬигаЕФЃЌ843ИіЪЧЁАStolbovayaЁБзщЬигаЕФЃЌЖј294ИіASVsдкСНИіЪЕбщзщжаЖМБЛМьВтЕНЃЈЭМ20.5CЃЉЁЃ
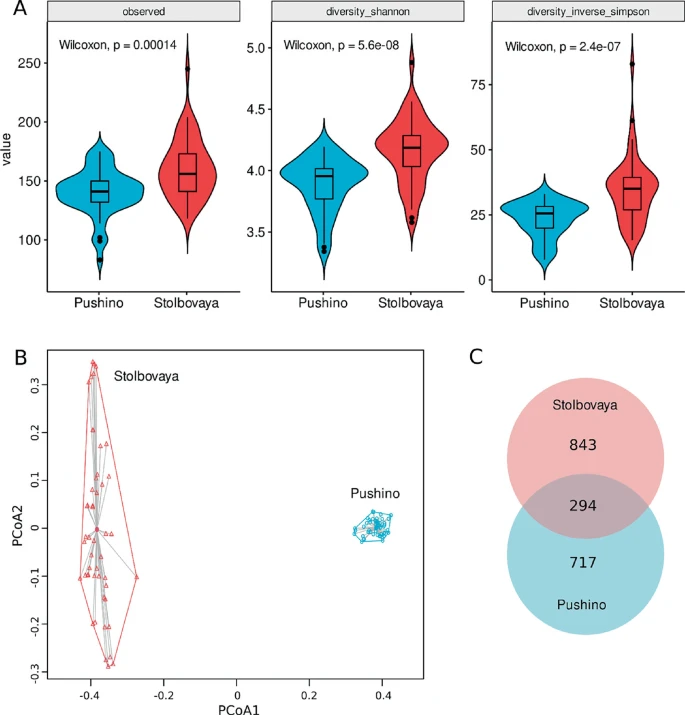
ЭМ20.5 ЪЕбщзщЕФЖрбљадЗжЮіЁЃA ЪЧalphaЖрбљаджИБъБШНЯЗжЮіЕФПЩЪгЛЏЃЛB ЪЧЁАPuschinoЁБКЭЁАStolbovayaЁБзщДѓЪѓШКЬхжаЯИОњШКТфЕФжїзјБъЗжЮіЃЈPcoAЃЉЃЛC ЪЧдкЪЕбщзщжаМьВтЕНЕФASVsЕФЮЌЖїЭМЁЃ
ЮЂЩњЮяШКТфзщГЩ
дкЫљгаЪЕбщзщжаЃЌДѓЪѓГІЕРОњШКЕФШКТфзщГЩжївЊгЩКёБкОњУХЃЈFirmicutesЃЉКЭФтИЫОњУХЃЈBacteroidotaЃЉЫљжїЕМЁЃдкРДздЁАStolbovayaЁБзщЕФЗрБубљБОжаЃЌCampylobacterotaЕФЪ§СПЯджјЃЌЖјдкЁАPuschinoЁБзщжаМИКѕЮДМьГіЁЃдкСНИізщжаЃЌФтИЫОњФПЃЈBacteroidiaЃЉКЭЫѓОњФПЃЈClostridiiЃЉНЯЮЊЦеБщЁЃдкФПМЖБ№ЩЯЃЌФтИЫОњФПЃЈBacteroidalesЃЉЁЂТна§ЬхФПЃЈOscillospiralesЃЉКЭLachnospiralesдкСНзщжаОљеМжїЕМЕиЮЛЁЃдкЦфЫћЯИОњФПжаЃЌШщИЫОњФПЃЈLactobacillalesЃЉдкЁАStolbovayaЁБзщжаеМгХЪЦЃЌЖјЫѓОњUCG-014ШКЃЈClostridia UCG-014ЃЉдкЁАPuschinoЁБзщжаИќЮЊЦеБщЁЃ
ГІЕРОњШКЦјЬЌДњаЛВњЮяЃЈЧтЦјКЭМзЭщЃЉЫЎЦНгыЗжРрзщГЩЕФЯрЙиадЗжЮі
ЭЈЙ§ШпгрЗжЮіКЭPearsonЯрЙиадЗжЮіЃЈдкmicroecoАќжаНјааЃЉРДЗжЮіЦјЬЌДњаЛЮяЃЈЧтЦјКЭМзЭщЃЉЫЎЦНгыЮЂЩњЮяШКТфзщГЩжЎМфЕФЯрЙиадЁЃНсЙћБэУїЃЌЁАStolbovayaЁБзщЗрБубљБОжаЕФЮЂЩњЮяШКТфгыМзЭщЫЎЦНЯрЙиЃЌЖјЁАPuschinoЁБзщЗрБубљБОжаИпЧтЦјЫЎЦНгыЮЂЩњЮязщГЩЧПСвЯрЙиЃЈЭМ20.6ЃЉЁЃдкЯИОњРрШКжаЃЌгыЧтЦјГЪзюЧПе§ЯрЙиЕФЪЧФтИЫОњЪєЃЈBacteroidesЃЉЁЂAlistipesКЭEnterococcusЃЌЖјМзЭщЫЎЦНгыЦеРзЮжЪЯОњЪєЃЈPrevotellaЃЉЁЂPrevotellaceae NK3B31ШККЭRickenellaceae RC9ГІЕРШКЕФДњБэГЪе§ЯрЙиЁЃ

ЭМ20.6 ЯрЙиадЗжЮіЁЃRDAХХађЭМеЙЪОСЫКєГіЦјжаЦјЬЌДњаЛЮяЃЈЧтЦјКЭМзЭщЃЉЫЎЦНгыЪЕбщзщжаЯИОњЪєЯрЖдЗсЖШжЎМфЕФЙиЯЕЁЃ
ЬжТл
ЮвУЧЕФбаОПЯдЪОЃЌРДздСНИіВЛЭЌбјжГГЁЕФДѓЪѓдкГІЕРОњШКЕФЗжРрзщГЩКЭЧтЦјгыМзЭщВњЩњБШР§ЗНУцПЩФмДцдкЯджјВювьЁЃ
вЛзщДѓЪѓЙКздPuschinoбјжГГЁЃЌОпгаSPFЃЈЮоЬиЖЈВЁдЬхЃЉзДЬЌЁЃетаЉДѓЪѓЕФЮЂЩњЮяШКТфЖрбљадНЯЕЭЃЌЭЈЙ§ЮЂЩњЮязщНјааЕФalphaЖрбљаджИБъЗжЮіЯдЪОЃЌShannonКЭФцЯђSimpsonжИЪ§гыСэвЛзщЃЈStolbovayaЃЉЯрБШУїЯдНЯЕЭЃЈp < 0.001ЃЉЁЃСНзщбаОПжаЕФЮЂЩњЮяШКТфЦзЯЕВювьКмДѓЁЃдкPuschinoзщЕФ1011ИіРЉдіађСаБфЬхЃЈASVsЃЉжаЃЌжЛга294ИігыStolbovayaзщЕФ1137ИіASVsЙВгаЁЃ
ХМШЛЗЂЯжСНзщДѓЪѓжЎМфДцдкЕФЯджјВювьЃЌПЩФмФЃФтСЫШЫРржаИпЧтзщКЭИпМзЭщзщЕФЧщПі[20, 21, 36]ЃЌЮЊВтЪдФкдДадЧтЦјПЩгУадПЩФмШЁОігкЫљЩуШЁЕФВЛвзЯћЛЏЬМЫЎЛЏКЯЮяЕФРраЭвдМАЧтВњЩњОњКЭМзЭщВњЩњОњЕФБШР§етвЛМйЩшЬсЙЉСЫЛљДЁЁЃ
ЮвУЧЕФбаОПНсЙћЯдЪОЃЌдкВњЧтДѓЪѓзщжаЃЌЭЈЙ§ЙрЮИИјгшШщЙћЬЧЁЂЙЯЖћНКЛђОеЗлКѓЃЌНіЙлВьЕНЯЫЮЌвРРЕадЕФЧтЦјЪЭЗХдіМгЁЃЯрЗДЃЌНЋЯрЭЌЕФЬМЫЎЛЏКЯЮяЙрЮИИјИпВњМзЭщДѓЪѓЃЈStolbovayaбјжГГЁЃЉКѓЃЌЙлВьЕНКєГіЦјжаМзЭщЕФдіМгЃЌЖјЗЧЧтЦјЁЃ
ОнЭЦВтЃЌГІЕРжаЕФЧтЦјжївЊгЩЪєгкFirmicutesУХЕФRuminococcus spp.ЁЂRoseburia spp.ЁЂClostridium spp.ЃЛЪєгкBacteroidetesУХЕФBacteroides spp.ЕШЯИОњВњЩњ[10, 37]ЁЃДЫЭтЃЌГЌЙ§200жжжТВЁОњПЩвдВњЩњЧтЦј[38]ЁЃвћЪГжаАќКЌВЛПЩЯћЛЏЕФЬМЫЎЛЏКЯЮяЭЈГЃЛсЕМжТГІЕРОњШКВњЩњЕФЧтЦјдіМг[7, 10, 23, 37, 39]ЁЃ
ФіГнЖЏЮябаОПЕФНсЙћНвЪОСЫВЛЭЌШКЬхЪЕбщДѓЪѓЕФГІЕРОњШКВњЩњЕФЧтЦјСПдкЩуШыЯрЭЌЩХЪГЯЫЮЌЕФБГОАЯТДцдкЯджјВювьЁЃвђДЫЃЌЕквЛзщДѓЪѓЕФУХОВТібЊЧтЦјХЈЖШЮЊ1.54 ІЬmol/lЃЌЖјЕкЖўзщЮЊ17.4 ІЬmol/lЁЃ
НЋВњЧтЦјИпЕФДѓЪѓНсГІОњШКЭЈЙ§ПкЗўвЦжВЕНЕквЛзщЕЭЧтЦјВњЩњЕФЖЏЮяжаЃЌЕМжТУХОВТіжаЧтЦјХЈЖШДг3.07діМгЕН9.95 ІЬmol/lЃЌвдМАЪєгкActinobacteriaУХЕФBifidobacteriumЪєЁЂFirmicutesУХЕФAllobaculumЪєКЭBacteroidetesУХЕФParabacteroidesЪєЯИОњЪ§СПЕФдіМгЁЃЭЌЪБЃЌЪєгкBacteroidetesУХЕФBacteroidesЪєЁЂFirmicutesУХЕФRuminococcusЪєКЭProteobacteriaУХЕФEscherichiaЪєЕФЫЎЦННЕЕЭ[40]ЁЃ
БШНЯВњЧтЦјКЭВњМзЭщДѓЪѓзщжЎМфЕФЗжРрбЇВювьБэУїЃЌЧтЦјВњЩњепЕФОњШКзщГЩНгНќгкаТЩњЖљКЭгзЖљЕФОњШКзщГЩЁЃЧтЦјВњЩњепЕФChristensenellaceaeЯИОњЗсЖШЪЧвбжЊЕФЧтЦјВњЩњСьЕМепЕФМИБЖЁЃ
ШЫРрЕФФъСфгыChristensenellaceaeКЭChristensenellaЕФЗсЖШГЪИКЯрЙиЃЌБэУїФъЧсЪмЪдепаЏДјИќИпЯрЖдЗсЖШЕФChristensenellaceaeКЭChristensenella[41]ЁЃGoodrichЕШШЫЗжЮіСЫ416ЖдЫЋАћЬЅЕФГІЕРОњШКЃЌЗЂЯжChristensenellaceaeЕФЗсЖШгыЕЭBMIЯрЙиЃЌЧвНЋChristensenella minutaвЦжВЕНЮоОњаЁЪѓжаПЩМѕЩйЬхжидіМг[42]ЁЃдкВњЧтЦјЕФДѓЪѓжаЗЂЯжСЫAkkermansiaЛљвђЃЌЕЋдкВњМзЭщЕФДѓЪѓжаУЛгаЗЂЯжЁЃVerrucomicrobiaУХЕФЗсЖШКЭAkkermansiaЪєAkkermansiaceae muciniphilaдкНЁПЕШЫРржадМеМ90%ЃЌеМЗрБуОњШКЕФдМ1%жС3%ЃЌВЂдкЩњУќЕквЛФъФкЖЈжГГІЕРЁЃЫќЕФСїааТЪПЩФмЛсЫцзХФъСфЕФдіГЄЛђМВВЁзДЬЌЖјНЕЕЭ[43]ЁЃ
AkkermansiaЕФЛљвђзщжаКЌгаЗсИЛЕФЧтЛЏУИЃЌШчHypEЁЂHypDЁЂHypAЁЂHypBЁЂHypFЁЂHypCЁЂHybGКЭHupFЃЌвђДЫAkkermansiaФмЙЛДпЛЏЗжНтКЭРћгУЧтЦјЁЃвђДЫЃЌГжајКЭГфзуЕФЧтЦјЙЉгІПЩФмДйНјетжжЯИОњзїЮЊгЊбјЮяжЪЕФДЋВЅ[44]ЁЃEverardЕШШЫеЙЪОСЫA. muciniphilaЕФЯрЖдЗсЖШгыЦЯЬбЬЧДњаЛЕФгаРћаЇЙћЯрЙиСЊЃЌЖјдкЗЪХжКЭЬЧФђВЁаЁЪѓМАШЫРржаМѕЩй[45]ЁЃ
ЪЕбщБэУїЃЌВњЧтЕФДѓЪѓЬхФкЕФHelicobacteraceaeЯИОњЃЈАќРЈHelicobacter pyloriКЭHelicobacter ssp.ЃЉЕФЪ§СПБШВњМзЭщЕФДѓЪѓЩйЫФБЖЁЃHelicobacteraceaeФмЙЛбѕЛЏЧтЦјЃЌЕМжТГІЕРжаЧтЕФМѕЩй[38]ЁЃдкВњЧтЕФДѓЪѓзщжаЃЌЮвУЧЮДФмМьВтЕНMethanobacteriaceaeЕФЗсЖШЃЈПЩФмЪЧЫќУЧЕФЫЎЦНЕЭгкЮвУЧЗНЗЈЕФМьВтЯоЃЉЃЌЕЋетаЉЙХОњдкВњМзЭщЕФзщжаБЛЗЂЯжЁЃ
вђДЫЃЌЛљгквдЯТЙлВьНсЙћЃЈЕЭalphaКЭbetaЖрбљадЁЂChristensenellaceaeКЭAkkermansiaЯИОњЕФИпЗсЖШЁЂHelicobacteraceaeЕФЕЭЗсЖШвдМАMethanobacteriaceaeЕФШБЪЇЃЉЃЌЮвУЧПЩвдМйЩшИпВњЧтЕФДѓЪѓзщПЩвдзїЮЊФъЧсШЫРрЮЂЩњЮяШКЕФФЃаЭЁЃ
ЮоОњДѓЪѓЕФЪЕбщЮЊГЄОУвдРДЕФЙлЕуЬсЙЉСЫЛљДЁЃЌМДМзЭщКЭЧтЦјвЛбљЃЌЭъШЋгЩГІЕРЮЂЩњЮяШКВњЩњ[46]ЁЃвЛаЉбаОПЕУГіНсТлЃЌШЫРрЕФЯИАћвВФмВњЩњМзЭщЃЌЕЋЫљгажЇГжетвЛНсТлЕФжЄОнЖМЪЧдкЬхЭтЛёЕУЕФЃЌВЂЧвЕНФПЧАЮЊжЙЛЙУЛгаЭЈЙ§ЬхФкЪЕбщЕУЕНжЄЪЕЁЃЛљгкетаЉЪ§ОнЃЌгаШЫЬсГіЃЌГ§СЫЮЂЩњЮяРДдДЭтЃЌПЩФмЛЙгаЦфЫћЩаЮДШЗЖЈЕФФкдДадМзЭщВњЩњЕФРДдД[47]ЁЃMethanobrevibacter smithiiКЭMethanosphaera stadtmanaeЪЧШЫРрНсГІжажївЊЕФМзЭщЩњВњепЁЃЫќУЧЕФЪ§СПбизХНсГІдіМгЃЌжБЕНдкжБГІДяЕНзюДѓжЕ[48]ЁЃMethanobrevibacter smithiiРћгУЧтЦјНЋЖўбѕЛЏЬМЛЙдЮЊМзЭщЃЌЖјMethanosphaera stadtmanaeдђРћгУЧтЦјНЋМзДМЛЙдЮЊМзЭщ[49, 50]ЁЃКЯГЩвЛИіМзЭщЗжзгашвЊЫФИіЧтЗжзгКЭвЛИіЖўбѕЛЏЬМЗжзгЃКЖўбѕЛЏЬМ + 4ЧтЦј Ёњ МзЭщ + 2H2OЁЃГІЕРЮЂЩњЮяШКжаВњМзЭщОњЕФЪ§СПЫцзХФъСфЕФБфЛЏЖјБфЛЏЁЃЭЈГЃЃЌаТЩњЖљЕФГІЕРЮЂЩњЮяШКВЛКЌгаВњМзЭщОњ[46]ЃЌвдМАШ§ЫъвдЯТЕФЖљЭЏ[51]ЁЃЫцзХФъСфЕФдіГЄЃЌВњМзЭщЕФШЫРрБШР§діМгЃЌ80-90ЫъФъСфзщЕФБШР§ДяЕН40-77%[52, 53]ЁЃЮвУЧЭЦВтЃЌИпВњМзЭщЕФДѓЪѓПЩвдзїЮЊРЯФъШЫЮЂЩњЮяШКЕФФЃаЭЁЃ
ЭЈЙ§ЪЙгУВЛПЩЯћЛЏЕФЬМЫЎЛЏКЯЮяШщЙћЬЧЁЂЙЯЖћНКЃЈЛЏбЇНсЙЙЮЊ100%ЙЯЖћЖЙАыШщИЪТЖОлЬЧЃЌгЩТќХЕЬЧКЭАыШщЬЧвдДѓдМ2:1ЕФБШР§зщГЩЃЉКЭОеЗлЃЈD-ЙћЬЧЕФОлКЯЮяЃЉИККЩЕФКєЮќВтЪдЃЌНідкВњЧтДѓЪѓжав§ЗЂСЫВЛЭЌЪ§СПЕФЧтЦјдіМгЁЃдкИјгшШщЙћЬЧКѓЃЌЧтЦјЗДгІЕФзюДѓдіМгЃЌОЁЙмЪГЮяЯЫЮЌЃЈЙЯЖћНККЭОеЗлЃЉЕФМССПЪЧШщЙћЬЧЕФСНБЖЃЌЕЋЪЧЙЯЖћНККЭОеЗлв§Ц№ЕФЧтЦјЗДгІЕФЗљЖШНЯЕЭЁЃBondКЭLevittдкШЫРрЩЯЕФЪЕбщжавВЯдЪОСЫРрЫЦЕФНсЙћЁЃетжжИіЬхЬМЫЎЛЏКЯЮядкВњЧтаЇЙћЩЯЕФВювьвбдкЬхЭт[24, 54]КЭЬхФкЪЕбщ[22]жаЯдЪОЃЌВЂПЩвдЭЈЙ§ЫожїЮЂЩњЮяШКЕФЗжРрзщГЩВювьРДНтЪЭЁЃР§ШчЃЌАыШщИЪТЖОлЬЧжївЊгЩB. ovatusНЕНтЃЌЖјОеЗлдђгЩBacteroides ovatusКЭBacteroides caccaeНЕНт[55]ЁЃШщЙћЬЧзюДѓЕФЧтЦјДЬМЄаЇЙћПЩвдНтЪЭЮЊЦфМђЕЅЕФНсЙЙЃЈШЫдьЖўЬЧЃЌКЌгаЙћЬЧКЭАыШщЬЧЃЉЃЌВЂЧвдкШЫРрЗрБужаЩИбЁГі35жжФмЙЛРћгУШщЙћЬЧЕФЮяжж[56]ЁЃ
дкВњМзЭщЕФДѓЪѓзщжаЙлВьЕНЫљгабаОПЕФЬМЫЎЛЏКЯЮяЕФвтЭтаЇЙћЁЃИјгшШЮКЮвЛжжШ§жжбаОПЕФЬМЫЎЛЏКЯЮяКѓЃЌНіАщЫцзХМзЭщВњСПЕФдіМгЁЃдкШЫРрбаОПжавВеЙЪОСЫРрЫЦЕФНсЙћ[21, 23]ЁЃГІЕРЮЂЩњЮяШКВњЩњЕФЧтЦјПЩвдБЛМИжжЧтгЊбјЬхЯћКФЃЌШчВњМзЭщЕФЙХОњЁЂВњЩњСђЛЏЧтЕФСђЫсЛЙдЯИОњКЭВњДзОњ[57]ЁЃВЛЭЌжжРрЕФВњМзЭщЙХОњРћгУЧтЦјгыЖўбѕЛЏЬМЁЂМзДМЛђШ§МзАЗЃЈTMAЃЉКЯГЩМзЭщ[10, 58]ЁЃгЩгкКЯГЩМзЭщашвЊЧтЦјЕФДцдкЃЌвђДЫВЛФмЫЕВњМзЭщЕФШЫРрЛђЖЏЮяВЛВњЩњЧтЦјЁЃЮвУЧМйЩшЃЌЕМжТНіВњЩњМзЭщЕФЬМЫЎЛЏКЯЮяИККЩЪЕбщЕФНсЙћПЩвдЭЈЙ§ДцдкИпЛюадЕФЧтгЊбјЬхЃЈБОР§жаЮЊМзЭщОњЃЉРДНтЪЭЃЌЫќУЧЯћКФСЫШЮКЮПЩгУЕФЧтЦјЁЃRuaudЕШШЫНјааЕФгХбХбаОПНсЙћЮЊЮвУЧЕФМйЩшЬсЙЉСЫЧПгаСІЕФжЇГжЃЌЫћУЧФмЙЛХрбјГіетбљЕФбсбѕОњЃЌШчВњЧтЕФChristensenellaКЭВњМзЭщЕФMethanobrevibacterЁЃдкЕЅЖРХрбјЪБЃЌChristensenellaЯИОњВњЩњЧтЦјЃЌЖјMethanobrevibacterЙХОњВњЩњМзЭщЁЃЕБСНжжЮЂЩњЮядкЭЌвЛЙмжаХрбјЪБЃЌЫќУЧаЮГЩСЫНєУмНгДЅЃЌГЦЮЊЭХПщЃЌChristensenellaВњЩњЕФШЋВПЧтЦјБЛMethanobrevibacterЯћКФ[41]ЁЃЧтЦјВњЩњОњЃЈBacteroides thetaiotaomicronЃЉКЭЧтгЊбјЬхЃЈDesulfovibrio vulgarisЃЉВњЩњСђЛЏЧтЕФХфЖдвВЯдЪОСЫРрЫЦЕФаЇЙћ[59]ЁЃ
ЮЊСЫЦРЙРГІЕРОњШКЕФзмЬхЗЂНЭФмСІЃЌЮвУЧШЯЮЊзюМбЗНЪНЪЧЦРЙРдЪМЕФЁЂГѕМЖВњЩњЕФЧтЦј(ЧтЦј)змСПЃЌетаЉЧтЦјПЩФмЫцКѓЭЈЙ§ЗЮВПКєГіЃЌБЛВЛЭЌЕФЧтгЊбјОњЯћКФЁЃГѕМЖВњЩњЕФЧтЦјзмСППЩвдМЦЫуЮЊКєГіЕФЧтЦјЃЈЕЅЮЛЮЊppmЃЉКЭКєГіЕФМзЭщЃЈЕЅЮЛЮЊppmЃЉЕФзмКЭГЫвд4ЕФЯЕЪ§ЁЃетжжМЦЫуЗНЗЈдкжЎЧАЕФбаОП[60]жавбБЛВЩгУЁЃЮвУЧМйЩшетЯюбаОПЕФНсЙћЖдгкНЁПЕвћЪГНЈвщвдМАЬсИпЛњЬхЕФПЙбѕЛЏФмСІОпгаживЊвтвхЁЃдіМгЪГЮяЯЫЮЌЕФЩуШыЪЧжкЫљжмжЊЕФНЈвщЃЌЕЋШчЙћГІЕРОњШКЕФИпВњМзЭщЛюадКмИпЃЌетНЋЪЧЮоаЇЕФЁЃЮвУЧШЯЮЊвћгУИЛКЌЧтЕФЫЎНЋЕМжТЧтЦјдкЮИГІЕРЩЯВПБЛЮќЪеЃЌРЉЩЂЕНбЊвКжаЃЌВЂЭЈЙ§УХОВТіЯЕЭГЕНДяИЮдрЃЌЦ№ЕНПЙбѕЛЏМСЕФзїгУЁЃ
ЙигкОњШКЗЂНЭЛюадбаОПЗНЗЈЕФМђБЈЁЃдкСйДВбаОПжаЃЌгУгкеяЖЯаЁГІЯИОњЙ§ЖШЩњГЄЃЈSIBOЃЉЁЂЮќЪеВЛСМЁЂВЛЭЌЬМЫЎЛЏКЯЮяЕФФЭЪмадЁЂПкУЄГІДЋЪфЪБМфвдМАЦфЫћвЛаЉВЁзДЃЌЧтЦјКєЮќВтЪдеМжїЕМЕиЮЛЁЃдкДѓЖрЪ§ЙигкФіГнРрЖЏЮяЃЈаЁЪѓКЭДѓЪѓЃЉЕФЪЕбщбаОПжаЃЌЖЏЮяБЛЗХжУдкУмЗтЕФВЃСЇЦПЛђДњаЛС§жа[25, 61,62,63,64]ЁЃ
етжжЗНЗЈгыШЫРрЕФКєЮќВтЪдВЛЭЌЃЌвђЮЊдкУмЗтПеМфжаХМЖћХХЦјЃЈЗХЦЈЃЉЪБЃЌИпХЈЖШЃЈвд%МЦЃЌЖјЗЧЯёКєГіПеЦјжаЕФppmЃЉЕФГІЕРЦјЬхЛсВњЩњЫљбаОПЦјЬхХЈЖШЕФЗхжЕЃЌДгЖјдіМгСЫНсЙћЕФРыЩЂЖШЁЃдкетЯюЙЄзїжаЃЌЮвУЧЪЙгУСЫвЛИіИФСМЕФНіБЧЯЕЭГНсКЯAllay™дМЪјЯюШІ[26, 27]РДМђЛЏКєЮќВтЪдГЬађЃЌВЂНіЪеМЏДѓЪѓКєГіЦјжаЕФЧтЦјЃЌетЪЙЕУИУЗНЗЈНгНќШЫРрЕФКєЮќВтЪдЃЌВЂБугкНЋЪЕбщНсЙћгыСйДВНсЙћНјааБШНЯЁЃ
НсТл
ВЛЭЌЪГЮяЯЫЮЌдкдіМгФкдДадЧтЦјЪЭЗХЗНУцЕФгааЇадШЁОігкОњШКжаЧтЦјВњЩњОњКЭМзЭщВњЩњОњЕФЗсЖШБШЁЃМзЭщВњЩњОњЕФГЪЯждНИпЃЌЪГЮяЯЫЮЌЕФЧтЦјДЬМЄаЇЙћОЭдНЕЭЁЃгЩгкЫцзХФъСфЕФдіГЄЃЌВњМзЭщОњЕФБШР§ИќИпЃЌЮвУЧЭЦВтЖдЪЉгУЕФЪГЮяЯЫЮЌЕФЛ§МЋЗДгІЛсИќЕЭЁЃЮвУЧШЯЮЊЃЌЖдгкРЯФъШЫШКЬхРДЫЕЃЌвћгУИЛКЌЧтЕФЫЎЛђЮќШыЧтЦјвдЬсИпЛњЬхЕФПЙбѕЛЏБЃЛЄНЋИќЮЊгааЇЁЃ
https://blog.sciencenet.cn/blog-41174-1427039.html
ЩЯвЛЦЊЃКAIШчКЮАяжњЬьЦјдЄБЈ
ЯТвЛЦЊЃКЩчНЛУНЬхаћДЋбаОПТлЮФПЩвдЬсИпВЮгыЖШЃЌЕЋВЛФмЬсИпв§гУТЪ
