博文
355.新型冠状病毒的先天免疫逃逸策略
||
新型冠状病毒的先天免疫逃逸策略
摘要
新型冠状病毒病毒是新冠肺炎疫情的罪魁祸首,与全球的发病率和死亡率密切相关。尽管嗜性主要局限于气道,但新冠肺炎与多器官功能障碍和长期认知病理学相关。这种生物学的主要驱动力来自于病毒介导的对受感染细胞中宿主抗病毒防御的干扰和旁观者细胞对病原体相关物质的感知的综合效应。这种动态导致在感染部位延迟诱导I型和III型干扰素(IFN-I和IFN-III ),但在远端器官和屏障上皮表面分别引发全身性IFN-I和IFN-III。在这篇综述中,作者研究了新型冠状病毒生物学和感染细胞反应之间的关系,详细说明了宿主先天免疫防御的拮抗和失调如何导致新冠肺炎病的严重程度。
介绍
病毒的进化成功取决于其进入细胞并获得原始遗传物质、氨基酸和重新产生子代病毒所需的基本细胞机制的能力。鉴于这些资源的价值,细胞已经进化出监视策略来检测它们的使用情况。在脊椎动物中,通过识别外来RNA或DNA结构来识别活跃复制的病毒。病毒衍生的RNA可以被识别为缺失修饰的结果(例如,2′-末端修饰)O-甲基化)、包含某些病毒区分元件(例如,暴露的5′-三磷酸)和/或广泛的二级结构(例如,双链RNA (dsRNA))的存在。相比之下,病毒来源的DNA可以通过其在细胞内的位置(例如,内体或胞质DNA)或通过宿主DNA中不存在的序列和/或结构的不同方面(例如,未甲基化的CpG基序)来鉴定。在这两个例子中,在被感染的细胞内对这些所谓的病原体相关分子模式(PAMPs)的细胞检测是直接的或通过自噬来辅助的,自噬是细胞吞噬和水解自身内部部分并随后将内容物释放回细胞质的过程。由于这些PAMPs代表着不可避免的复制副产品,病毒处于不断的进化压力下,以最小化它们的水平,防止它们被检测到和/或阻断任何随之而来的下游生物学。
破坏细胞检测的病毒策略,如果成功的话,不仅能够在同源宿主中发生生产性感染,而且还能够促进人畜共患事件,因为许多细胞防御成分在系统发育相关的物种中是保守的。出于这个原因,一个物种侵入另一个物种的生态系统是动物传染病的常见来源,也是人口过度增长的常见动力——许多新出现的病毒都是毁林的产物这一事实就证明了这一点(方框1)。通过畜牧业、活水市场或获取野味与其他脊椎动物物种的密切接触也是人畜共患病传播的主要来源,并被认为是人类免疫缺陷病毒和无数流感病毒在疫情传播的原因。活水市场等人畜共患病环境也导致了两种高致病性冠状病毒的出现,即2003年的严重急性呼吸综合征冠状病毒(SARS-CoV)和2019年的新型冠状病毒,后者导致了2019年的冠状病毒疾病(新冠肺炎)疫情。
尽管疫苗和抗病毒药物的快速开发和部署,但从2020年初到2022年底,新型冠状病毒造成了超过6.5亿例感染和660万例死亡。随着全球传播的继续,新型冠状病毒经历了无数轮的选择:适应人的生物学,绕过人的免疫防御,提高传播效率。由于这种选择而获得竞争性适合度优势的单个毒株被定义为相关变异体(VOC ),并用希腊字母表中的字母进行命名。在这篇综述中,作者总结了新型冠状病毒绕过我们先天抗病毒防御的第一道防线的机制,并讨论了这与新冠肺炎严重程度的关系。对于聚焦于对新型冠状病毒适应性免疫应答的后期阶段的可比较的综述,作者向读者推荐在别处可获得的几篇写得很好的和全面的摘要。
方框1 .新型冠状病毒是一座警示灯塔
人类对全球生态系统的影响如此深远,以至于许多人认为我们已经进入了一个被称为人类世的新时代。不管这一地质时代的划分是否得到官方认可,人类对地球健康的净负面影响是不可否认的。尽管有近40亿年的自我维持生命,但人类在短短几千年内以无数方式破坏了这种平衡。不断增长的人口导致了森林砍伐、污染和全球气温升高——在全球旅行不受限制的时候破坏了生态系统。这些变化迫使物种寻找新的栖息地,使它们和它们的病原体更接近潜在的新宿主。随着暴露频率的增加,病毒和其他微生物病原体有机会适应这些免疫天真的宿主,进入一个新的生态位,在那里它们可以获得疫情潜力。仅仅在过去的一个世纪里,人类已经观察到数十种新的病毒流行病的出现,其中一半已经成为流行病。人类应该认识到这些事件发生频率的增加,因为它们代表了什么——这个星球被忽视的副产品。此外,我们对新型冠状病毒病毒等病毒的免疫逃逸策略的了解越来越多,这可以在为监测和测试措施提供信息方面发挥重要作用,有助于优先关注各种新出现的威胁。展望未来,作者应该像致力于开发新技术和新疗法以最大限度地减少未来的流行病一样,致力于减少对环境的影响,理解人类行为与新出现的传染病之间的联系。
细胞对新型冠状病毒感染的反应
新型冠状病毒的基因组,像其属的所有成员一样,长度约为30,000个核苷酸,具有不变的基因序列。从5’端开始是复制酶和其他非结构成分的基因(ORF1a和ORF1ab),随后是刺突(S)、包膜(E)、膜(M)和核衣壳(N)蛋白的结构基因,以及许多基因间辅助因子。新型冠状病毒的正义,单链RNA基因组编码约30种蛋白质。这些蛋白包括ORF1a和ORF1ab多蛋白(pp1a和pp1ab),它们被病毒蛋白酶(PLpro和3CLpro)切割成16种非结构蛋白(Nsps ),主要形成RNA依赖性病毒复制酶,4种结构蛋白形成物理病毒体,9种辅助蛋白不是病毒复制所必需的,但对细胞生物学的劫持方面是必需的。
与大多数病毒感染一样,宿主对冠状病毒感染的先天免疫反应的关键第一步是产生I型和III型干扰素(分别为IFN-I和IFN-III)以及促炎细胞因子和趋化因子。IFN-I和IFN-III由多种细胞类型在识别病毒PAMPs和/或宿主危险相关分子模式(DAMPs)后产生,例如通过专门的细胞模式识别受体(PRRs)识别暴露的线粒体DNA。对于新型冠状病毒,基于RNA的复制中间体被认为是主要的病毒PAMP,被RIG-I样和Toll样受体(分别为RLRs和TLRs)识别。RLRs包含一个RNA结合解旋酶家族,包括两个中心细胞内哨兵蛋白RIG-1和MDA5,以及第三个LGP2,其功能尚不清楚。尽管它们表现出结构上的相似性,但RLRs具有不同的结合偏好,这使得它们能够组成一个多样化和互补的防御系统。一般来说,RIG-I被认为可以感知具有暴露的5′-三磷酸的RNA,这是负链RNA病毒的共同特征,而MDA5的激活是通过参与dsRNA来介导的,dsRNA是所有病毒复制过程中形成的共同基序。与这些细胞内的哨兵相反,TLRs对细胞外环境进行取样,以确定远端感染引起的PAMPs或DAMPs的存在。由于涉及RNA病毒感染的检测,细胞外dsRNA、单链RNA或DNA可以分别被TLR3、TLR7/TLR8或TLR9检测到。虽然所有这些典型的感应机制,以及一些非典型的系统,都被认为有助于宿主识别新型冠状病毒,但MDA5被认为在这个过程中是最关键的。
新型冠状病毒的细胞识别促进衔接蛋白的募集和IFN-I和IFN-III的产生。在没有干扰的情况下,RLR激活会导致病毒亚基因组RNA (sgRNA)的产生,其可以通过接合基因组模板容易地形成dsRNA结构。MDA5聚集在dsRNA结构上,随后与线粒体抗病毒信号蛋白(MAVS)结合,导致泛素连接酶和丝氨酸/苏氨酸激酶的募集,从而协调两个核心转录因子家族成员核因子-κB (NF-κB,包含亚单位p50和p65)和干扰素调节因子(IRF)如IRF3。类似地,TLR识别合适的病毒配体启动了MyD88或TRIF细胞衔接蛋白的募集,并通过类似的信号级联触发了NF-κB和IRF的激活。除了与细胞应激相关的其他因子之外,这些转录因子的激活导致称为增强体的大复合物的形成,该复合物结合IFN-I成员(特别是IFNβ)和IFN-III成员(如IFNλ-1、IFNλ-2和IFNλ-3)的转录起始位点的上游,从而启动抗病毒反应。尽管诱导相似的转录输出,但几个特征使这两个抗病毒细胞因子家族具有不同的生物学特性,包括IFN-I和IFN-III的细胞表面受体的组成和分布,蛋白质稳定性的变化,以及每个反应的大小、动力学和解剖学位置。尽管IFN-I几乎可以引发任何细胞以诱导抗病毒状态,但IFN-III的活性更具选择性,并被认为仅限于上皮屏障组织,包括呼吸道和胃肠道以及血脑屏障。
IFN-I和IFN-III信号传导以自分泌和旁分泌的方式促进干扰素刺激基因(ISGs)的上调,如IRF7,一种与IRF3相关的转录因子,表现出更混杂的DNA结合活性。IRF7的加入扩大了由病毒识别产生的转录输出,包括更多的IFN-I家族成员,最显著的是IFNα变体。IRF7和其他ISG的诱导是由IFN-I依赖性或IFN-III依赖性受体二聚化介导的,二聚化最终导致称为IFN刺激的基因因子3 (ISGF3)的转录复合物的激活和装配,该转录复合物包括信号转导和转录激活子(STAT1和STAT2)家族成员,以及一个额外的IRF家族成员(IRF9)。活化的ISGF3然后迁移到细胞核,在那里它协调数百个ISG的转录,这些ISG通过多种机制直接或间接发挥抗病毒作用,包括抑制病毒复制、抑制病毒转录/翻译和降解病毒核酸。
在缺乏IFN-1受体表达的动物(IFNAR1−/−),其中通常产生轻微疾病的小鼠肝炎病毒株(MHV)变得致命。这种表型的基础源于冠状病毒在宿主中引发全身性IFN-I反应,从而保护远端器官免受后续感染。在没有这种反应的情况下,远端组织容易受到低水平循环病毒的影响,这通常与疾病加重有关。在患有危及生命的新冠肺炎的年轻患者中观察到了这种动态,他们被发现具有IFNAR1或者TLR基因以及据报道存在IFN-1自身抗体的高龄患者。感染期间IFN-I或IFN-III、ISGs和PRRs的解剖表达差异也以年龄依赖性方式与新冠肺炎患者的疾病严重程度相关。
除了诱导IFN-I或IFN-III,脊椎动物还产生促炎细胞因子和趋化因子来对抗病毒感染。与IFN-I不同,IFN-I代表在感染部位的“战斗号召”,细胞因子和趋化因子有助于协调来自更远部位的“增援号召”。当功能正常时,这两种互补的途径可以减缓感染部位的复制,并为适应性免疫反应提供时间来发展抗原特异性识别能力,从而为任何病毒的进化成功建立了一个可怕的障碍。不幸的是,病毒已经进化出无数的策略来干扰这些宿主防御,导致经常不可预测的疾病结果,这些结果是由病毒直接引起的,由异常的宿主反应引起的,或者由两者的结合引起的。在接下来的部分,作者描述了关于新型冠状病毒介导的宿主生物学操作的知识现状,并提供了关于这种病毒干扰如何导致疾病严重程度的见解。
新型冠状病毒对宿主防御的规避
作为专性细胞内寄生物,所有病毒都依赖宿主获得能量、原材料和进入复杂的生物机器。为了建立生产性感染,病毒必须侵占或灭活广泛的宿主途径。新型冠状病毒投入大量资源阻止抗病毒反应的建立(表1)。这些策略分为五大类,详细描述如下。
表1负责先天免疫干扰的新型冠状病毒蛋白
开放阅读框 | 活性 | 机制 |
Nsp1 | 阻止宿主传感器的识别 | 防止IRF3磷酸化,可能是通过翻译关闭耗尽所需的细胞因子 |
阻止IFN信号a | 耗尽TYK2和STAT2 | |
阻断核运输a | 与mRNA输出受体异二聚体NX f1–NX t1相互作用,并削弱其与mRNA输出因子和参与核输出的核孔蛋白相互作用的能力 | |
关闭翻译 | 促进不含5’病毒前导序列的细胞mRNA的降解 通过其C末端的结构域与40S核糖体亚单位的18S结构RNA成分结合,阻断mRNA进入核糖体的通道 | |
Nsp3 | 最小化或掩盖炎症RNA | 需要形成与ER相关的DMVss |
阻止宿主传感器的识别a | PLpro结构域设计MDA5 劈开IRF3 | |
损害宿主蛋白质功能a | 宏结构域-X与氨基酸链结合并水解ADP-核糖键 PLpro结构域去泛素化和去糖基化宿主信号蛋白底物 | |
Nsp4 | 最小化或掩盖炎症RNA | 需要形成与ER相关的DMVss |
Nsp5 | 阻止宿主传感器的识别a | 抑制应力颗粒的形成 切割RIG-I的N-末端结构域并阻止其与MAVS相互作用 促进泛素化和MAVS的降解 劈开IRF3 防止IRF3的核转移;独立于Nsp5蛋白酶活性或IRF3磷酸化 通过切割TAB1和NEMO来阻止NF-κB的磷酸化和活化 |
Nsp6 | 最小化或掩盖炎症RNAa | 栓 DMVss到ER |
阻止宿主传感器的识别 | 结合并阻止磷酸化介导的TBK1激活 | |
阻止IFN信号 | 防止STAT1和STAT2的磷酸化 | |
Nsp8 | 关闭翻译 | 结合SRP复合物的7SL RNA支架成分,阻断其结合SRP19的能力,这是正确折叠和组装SRP所必需的 |
Nsp9 | 阻断核运输a | 与核转运机制相互作用并削弱核包膜上Nup62的表达 |
关闭翻译 | 结合SRP复合物的7SL RNA支架成分,阻断其结合SRP19的能力,SRP 19是SRP正确折叠和装配所必需的 | |
Nsp10 | 最小化或掩盖炎症RNA | 在病毒加帽期间充当Nsp14和Nsp16的辅因子 |
关闭翻译 | 增强Nsp14介导的翻译抑制 | |
Nsp12 | 最小化或掩盖炎症RNAa | 在病毒mRNA加帽过程中充当鸟苷酸转移酶 |
阻止宿主传感器的识别 | 防止IRF3的核转移;独立于Nsp12聚合酶活性或IRF3磷酸化 | |
Nsp13 | 最小化或掩盖炎症RNA | 病毒mRNA加帽过程中的5′RNA三磷酸酶活性 |
阻止宿主传感器的识别 | 结合并阻止磷酸化介导的TBK1激活 | |
阻止IFN信号 | 降低IFNAR1的内源性水平 防止STAT1和STAT2的磷酸化 | |
Nsp14 | 最小化或掩盖炎症RNA | 病毒mRNA加帽期间的n7-甲基转移酶活性 |
阻止IFN信号 | 靶向IFNAR1进行溶酶体降解 | |
激活NF-κBa | 增加p65的核转位和促炎性趋化因子的上调,包括IL-6和IL-8 | |
关闭翻译 | 以依赖于外显子结构域和与Nsp10相互作用的方式阻断蛋白质合成 | |
Nsp15 | 最小化或掩盖炎症RNA | 内切核糖核酸酶活性从病毒RNA的负链上切割5′-多聚尿苷以减少病毒PAMPs的积累 |
阻断核运输a | 与宿主核转运机制相互作用(核转运因子2) | |
Nsp16 | 最小化或掩盖炎症RNA | 2′-O病毒mRNA加帽期间的-甲基转移酶活性 |
关闭翻译a | 结合剪接体的snRNA U1和U2亚单位的mRNA识别域 | |
ORF3a | 阻止IFN信号 | 防止STAT1的磷酸化 |
ORF3b | 阻止宿主传感器的识别 | 防止IRF3的核转移 |
ORF6 | 阻断核运输a | 结合核外激素-α2 (KPNA2)输入蛋白 与NUP 98–RAE 1复合物结合并阻止其与NPC结合 促进宿主mRNA和mRNA转运蛋白的核积累;依赖于ORF6 C末端 |
ORF7a | 阻止宿主传感器的识别 | 减少TBK1的表达 |
阻止IFN信号 | 阻断STAT1和STAT2的磷酸化 | |
ORF7b | 阻止宿主传感器的识别 | 以依赖于MAVS的方式阻止RIG-1和MDA5信令 |
阻止IFN信号 | 阻断STAT1和STAT2的磷酸化 | |
ORF8 | 激活NF-κBa | 诱导人IL-17受体异二聚化和NF-κB下游激活的IL-17A病毒模拟物 |
ORF9b | 阻止宿主传感器的识别 | 防止RIG-I和MAVS之间的相互作用 与TOM70结合并抑制TOM70/HSP90相互作用,可能导致对TBK1/IRF3信号的干扰 通过阻止TBK1和TRIF之间的相互作用来阻断TBK1磷酸化 |
S | 阻止宿主传感器的识别 | 加强IRF3的蛋白酶体降解 |
阻止IFN信号 | 防止STAT1与JAK1相互作用 | |
激活NF-κB | 促进p65和IκBα的磷酸化;依赖于S1亚单位 | |
M | 阻止宿主传感器的识别 | 通过损害MAVS形成募集信号衔接子所必需的大聚集体的能力来阻断其激活 通过泛素介导的降解减少TBK1的表达 |
阻断核运输 | 结合KPNA6输入蛋白并阻断其与IRF3的相互作用 | |
核衣壳 | 最小化/掩盖炎症RNA | 结合和去稳定dsRNA 由于其在病毒体装配中的作用而具有固有的RNA结合特性 |
阻止宿主传感器的识别a | 通过结合和隔离G3BP1成核蛋白阻断应激颗粒的形成 与RIG-I的DExD/H盒RNA解旋酶结构域结合,并阻断其与TRIM25的相互作用 可能通过LLPS抑制MAVS的多泛素化和聚集 |
00001. DMVs,双膜囊泡;dsRNA,双链RNA;ER,内质网;ExoN,核酸外切酶;IFN,干扰素;IRF,干扰素调节因子;JAK1,Janus激酶1;KPNA6,核蛋白亚基α6;LLPS,液-液相分离;MAVS,线粒体抗病毒信号蛋白;NF-κB,核因子-κB;NPC,核孔复合体;Nsp,非结构蛋白;Nup,核孔蛋白;NXF1,核RNA输出因子1;NXT1,核转运因子2-样输出因子1;PAMP,病原体相关分子模式;PLpro,木瓜蛋白酶样半胱氨酸蛋白酶;Rae1,核糖核酸输出因子1;snRNA,小核RNASRP,信号识别粒子;STAT、信号转导子和转录激活子;TYK2,酪氨酸激酶2。a表示在新型冠状病毒感染背景下完成的研究支持的机制信息。
最小化和掩蔽炎症RNA
冠状病毒感染循环的第一步涉及三聚体S糖蛋白与宿主细胞受体的结合,这导致与细胞质膜直接融合的“早期”进入途径或通过内吞作用的受体介导的摄取的“晚期”进入途径。在新型冠状病毒的情况下,这两种途径都是通过S三聚体与宿主血管紧张素转换酶2 (ACE2)的结合来介导的。为了发生与宿主膜的直接融合,S蛋白必须在病毒体外出过程中被切割,然后在进入过程中再次被切割。对于新型冠状病毒,第一个事件主要由弗林蛋白酶介导,第二个事件由细胞表面的跨膜丝氨酸蛋白酶2 (TMPRSS2)介导。或者,在受体介导的胞吞过程中,组织蛋白酶也可以介导S蛋白的双重切割。在任一种情况下,一旦基因组病毒核糖核蛋白复合物进入细胞质,基因组就从病毒N蛋白上解离下来,此后可被宿主核糖体直接翻译。
病毒基因组RNA的初始翻译必须符合宿主翻译的规则,这既是为了功能性也是为了避免宿主检测。为此,冠状病毒通过加帽和甲基化来修改其基因组RNA和sgRNAs的5′-三磷酸(pppA),以确保核糖体加载并避免宿主检测(图.1)。冠状病毒帽结构的形成需要四个连续的酶促反应。首先,5′RNA三磷酸酶从新生mRNA的pppA上去除γ-磷酸,形成5′-二磷酸(ppA)。接下来,鸟苷酸转移酶通过将一磷酸鸟苷(GMP)转移到ppA来产生帽核心结构。核心结构形成后,鸟嘌呤在N7位被N7-甲基转移酶甲基化。最后,一个2′-O甲基转移酶在2′-甲基化核糖O产生最终帽结构的第一个转录核苷酸的位置:7MeGpppA2′OMe。对新型冠状病毒和其他冠状病毒进行的研究表明,Nsp13、Nsp14和Nsp16在病毒加帽过程中作为5′RNA三磷酸酶、N7-甲基转移酶和2′-甲基转移酶发挥作用O-甲基转移酶。新型冠状病毒Nsp12也被认为作为鸟苷酸转移酶参与了这一过程。其他研究表明,Nsp10可能通过作为Nsp14和Nsp16活性的辅因子参与病毒加帽,NSP 14和NSP 16的功能在冠状病毒中是保守的。
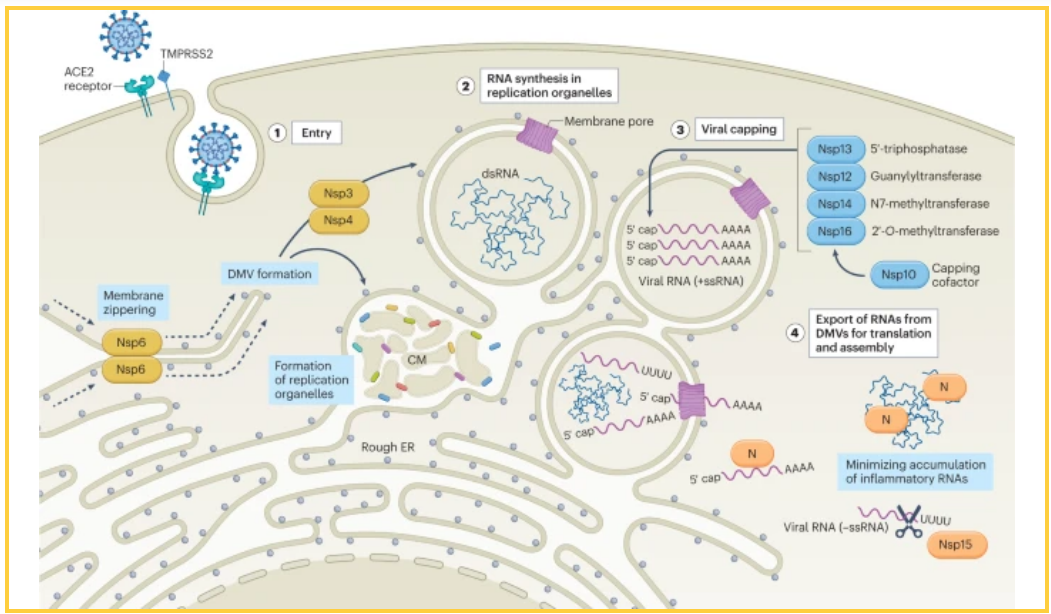
图1:最小化宿主检测的新型冠状病毒策略。新型冠状病毒通过结合细胞表面的血管紧张素转换酶2 (ACE2)进入宿主细胞,这一过程可由跨膜蛋白酶丝氨酸2 (TMPRSS2)促进,它提供病毒刺突蛋白的蛋白水解切割以促进病毒-宿主融合。病毒颗粒内化后,加帽和多聚腺苷酸化的基因组病毒RNA被释放到细胞质中,在那里它可以被直接翻译(阶段1)。病毒基因组RNA的初始翻译导致ORF1a和ORF1ab多蛋白(pp1a和pp1ab)的产生,其随后被病毒蛋白酶加工以形成复制酶和非结构蛋白(Nsps用黄色表示)是建立复制细胞器(ROs)所必需的。Nsp3和Nsp4介导内质网(er)膜修饰为构成ROs的回旋膜(CM)和双膜小泡(DMVs),而Nsp6在ROs和ER之间形成拉链状分子链,使脂质能够流动(阶段2)。新生的病毒RNA被Nsp酶(用蓝色表示)修饰,以模拟宿主转录物,并使细胞诱导防御的能力最小化。首先,Nsp13(一种5’RNA三磷酸酶)从病毒RNA的5’末端去除磷酸。随后通过Nsp12(一种鸟苷酸转移酶)将一磷酸鸟苷转移到5’末端,产生帽核心。随后,Nsp14(一种N7-甲基转移酶)和Nsp16(一种2′-甲基转移酶)O-甲基转移酶)催化完成病毒加帽所必需的最终甲基化步骤(阶段3)。随着病毒复制的进行,反义RNA(ssRNA)和双链RNA (dsRNA)中间体被隔离在ROs内,以防止宿主检测。平行地,翻译病毒蛋白和重新组装病毒粒子所需的正链、单链基因组和亚基因组RNA(+ssRNA)由复制细胞器陪伴。随着复制的加剧,病毒RNA在ROs外积累,并被新型冠状病毒核衣壳蛋白(N)和/或Nsp15掩盖和/或最小化,如橙色所示(阶段4)。
因为病毒mRNA伪装成宿主mRNA,复制酶的翻译和装配最初不应引发细胞反应;然而,随着病毒RNA合成的进行,dsRNA中间体不可避免地开始形成,从而触发宿主免疫激活。其原因是基因组RNA和sgRNAs都必须通过负链中间体。这种具有相反极性和互补序列的RNA转录物的积累增加了dsRNA形成的可能性。对于新型冠状病毒和其他冠状病毒,dsRNA的早期检测决定了病毒的整体适应性。在病毒生命周期完成之前,宿主诱导的抗病毒防御足以将感染限制在最初被感染的细胞中。因此,在dsRNA产生之前,病毒必须确保策略到位,要么最小化其积累,要么将其隐藏起来,以免被宿主传感器发现:新型冠状病毒同时利用了这两种策略。据报道,高度保守的Nsp15通过其靶向反义转录物的核酸内切酶活性,负责最小化负链RNA和dsRNA的积累。与此同时,新型冠状病毒诱导双膜囊泡(DMVs)的组装,这已在其他冠状病毒如MHV中提出,以将病毒复制与细胞传感器分隔开来。这些所谓的复制细胞器(ROs)与内质网相关,并在感染MHV、SARS-CoV、中东呼吸综合征冠状病毒(MERS-CoV)和新型冠状病毒。建立ROs的过程也是通过干扰和/或利用自噬生物学来削弱宿主识别病毒感染存在的能力的一种手段。新型冠状病毒感染期间的RO组装由Nsp3和Nsp4功能介导,Nsp6形成内质网的分子系绳,使脂质能够流动(图.1)。由于积累足够水平的Nsp3、Nsp4、Nsp6和Nsp15需要时间,所以用低的感染多重性实现了最佳感染,从而在DMVs形成或翻译足够水平的Nsp15之前,复制酶装配和早期基因组复制不会产生大量的dsRNA。此外,可以观察到新出现的挥发性有机化合物的先天免疫激活的差异,这是影响参与该过程的病毒蛋白的丰度、表达或功能的非同义变化的结果。
阻止宿主识别
随着病毒复制器的成功建立,新型冠状病毒转录物通过特殊的分子孔离开DMVs并进入胞质溶胶,在那里它们经历翻译并开始病毒体装配。在密封ROs的保护之外,病毒RNA可能被细胞传感器识别(图.2)。随着复制的加剧,通过排除任何炎症病毒RNA来避免宿主检测将很快变得不可能。因此,冠状病毒也投入大量资源来抑制识别后产生的细胞信号通路。
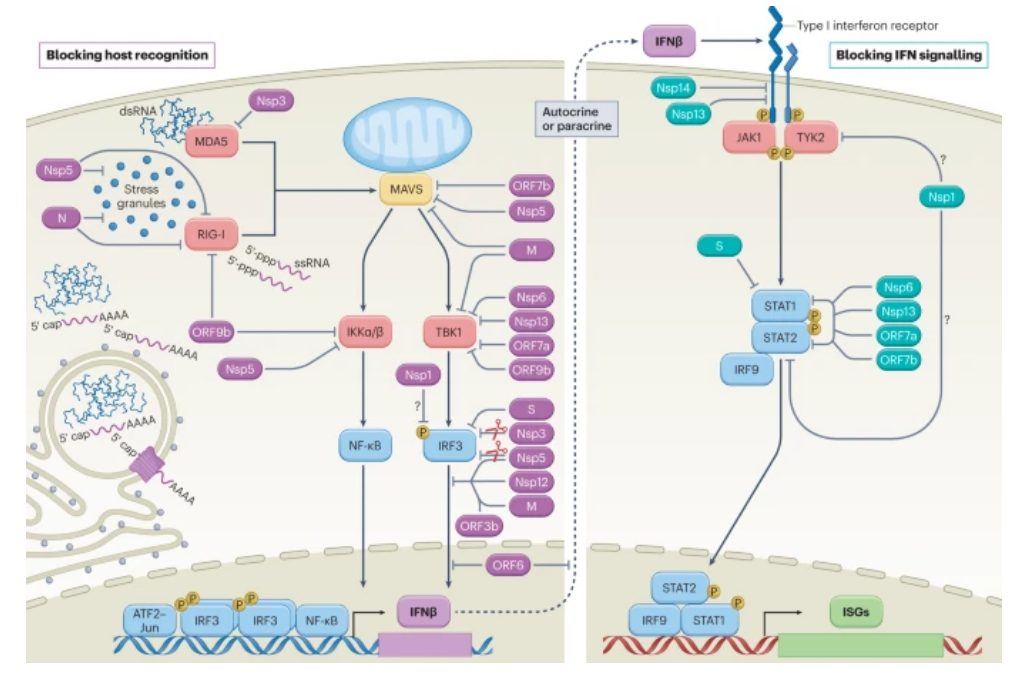
图2:新型冠状病毒介导的细胞先天免疫信号的干扰。病毒感染产生复制中间体和/或诱导应激颗粒的形成,这些颗粒充当RIG-I样受体(RIG-I或MDA5)激活的平台。宿主对病毒病原体相关分子模式的识别,如具有暴露的5′-三磷酸的单链RNA (ssRNA)或双链RNA (dsRNA),促进由线粒体抗病毒信号蛋白(MAVS)协调的线粒体定位信号中枢的组装,并最终激活宿主激酶IKKα、IKKβ和TBK1。激酶激活通过at F2–JUN、干扰素调节因子3 (IRF3)和核因子-κB (NF-κB)转录因子的协同作用诱导干扰素-β (IFNβ)的产生。分泌的IFNβ以自分泌或旁分泌的方式促进细胞的抗病毒状态。在结合IFNβ时,细胞表面的I型IFN受体亚单位二聚化,将受体相关激酶Janus激酶1 (JAK1)和酪氨酸激酶2 (TYK2)聚集在一起,它们随后通过转磷酸作用相互激活,并促进信号转导和转录激活因子(STAT)分子STAT1和STAT2的募集和磷酸化。磷酸化的STAT1和STAT2与干扰素调节因子9 (IRF9)形成稳定的复合物,转运到细胞核中,在那里促进IFN刺激基因(ISGs)的转录。这些过程中的每一个都是新型冠状病毒干扰的目标,如下图所示,并在表1中进一步描述。抑制宿主识别和相关信号通路的病毒蛋白显示为紫色,而阻断IFN信号通路的病毒蛋白显示为青色。由于Nsp1更普遍地抑制蛋白质合成的能力,它在特异性阻断这些途径的元件中的作用仍然不确定(问号)。m,膜蛋白;n,核衣壳蛋白;p,磷酸化;s,刺突蛋白。
为了导航这个先进的监测系统,一些新型冠状病毒蛋白被建议靶向病毒感应过程,并延迟宿主抗病毒防御的产生(图.2)。一个被认为与这一角色有关的核心病毒成分是N蛋白。作为感染过程中产生的最丰富的病毒蛋白之一,这种RNA结合蛋白对于将病毒基因组包装成病毒体至关重要。由于它与免疫拮抗作用有关,N与游离RNA结合的固有能力也可用于掩盖其不被检测或防止其与互补序列形成广泛的碱基对。支持这一假设的是,使用α(b . 1 . 1 . 7)VOC的体外研究表明,与早期的病毒谱系相比,用这种变体观察到的增强的IFN拮抗作用可能与增加的N蛋白表达有关;其他的解释,如dsRNA生产的总体减少或其他非结构基因表达的增加,仍然是可能的。最近的一项研究还表明,宿主胱天蛋白酶6切割后产生的N蛋白片段本身可以抑制宿主的IFN反应,进一步增加了N可以作为IFN拮抗剂的证据。这种由N引起的潜在病毒干扰的高潮是先天免疫激活的延迟,为新型冠状病毒提供了在最佳细胞环境中复制和传播的关键时间。
诱导RLR信号传导的另一个机制涉及应激颗粒的形成(图.2)。这些翻译停滞的 mRNAs 和RNA结合蛋白的无膜细胞质聚集体在各种应激条件下形成,包括病毒感染,并作为RLR信号通路的平台。Nsp5是新型冠状病毒的主要蛋白酶,它可以独立于其酶活性抑制这些应激颗粒的形成。新型冠状病毒N蛋白还可以通过结合和隔离RNA和一种称为G3BP1的特定因子来阻止应激颗粒的形成,G3BP1不仅作为应激颗粒的成核蛋白,而且还作为应激颗粒和RLRs之间的联系。
除了制定隔离病毒PAMPs的机制,新型冠状病毒蛋白还可以阻断PRR的激活。已经证明N蛋白与RIG-I的DExD/H盒RNA解旋酶结构域结合,中断其与TRIM25的相互作用,TRIM25是一种细胞泛素连接酶,通过K63-泛素介导的RIG-I激活增强RLR信号传导。然而,这些研究的结论是基于过表达系统,并没有说明基因组和亚基因组新型冠状病毒RNA都被加帽,或者RIPLET,而不是TRIM25,被认为是RIG-I激活的更相关的衔接子。类似地,Nsp3蛋白,也称为新型冠状病毒木瓜蛋白酶样半胱氨酸蛋白酶(PLpro ),已被报道拮抗称为ISG15的ISG与MDA5的结合,但这些数据也依赖于过表达实验,并集中于具有不清楚生理相关性的激活途径。Nsp3可能在宿主免疫逃逸中发挥更重要的作用,因为它具有干扰其他蛋白质修饰的一般能力,包括泛素和/或ADP-核糖缀合。尽管存在序列差异,但Nsp3改变翻译后修饰的能力在冠状病毒中很大程度上是保守的,这表明该蛋白在宿主逃逸中具有重要的全球作用。
由于细胞对病毒感染的识别不太可能被完全阻止,其他病毒成分在信号级联中更下游靶向宿主因子。这种策略使病毒能够干扰多种抗病毒监视系统,这些系统集中在某些细胞适配器的激活上。例如,对N、M、Nsp5和ORF7b的过表达研究都产生了支持新型冠状病毒干扰MAVS生物学能力的数据(图.2)。尽管对MAVS的作用机制尚未完全阐明,但最近的证据表明N蛋白可能以依赖于N二聚化结构域的方式抑制MAVS多泛素化和聚集。值得注意的是,该结构域也是N与病毒RNA相互作用的液-液相分离(LLPS)所必需的。除了在介导病毒装配中的可能作用外,N的LLPS还与它与应激颗粒和NF-κB信号通路的成分相互作用的能力相关,提供了额外的证据支持它在免疫逃逸和调节中的作用。新型冠状病毒蛋白采用的其他传感器靶向策略包括直接泛素化和蛋白酶体介导的MAVS降解,一种由Nsp5介导的活性。类似地,据报道,新型冠状病毒的M蛋白干扰MAVS建立下游转录因子激活所需的必要支架的能力。新型冠状病毒M、Nsp6、Nsp13、ORF7a和ORF9b蛋白也已被证明靶向并抑制参与MAVS信号传导的其它宿主因子的功能,如负责IRF3激活的激酶TBK1。特别是,Nsp6、Nsp13和ORF9b都被认为与TBK1结合,阻止其磷酸化介导的激活,而据报道M和ORF7a降低了TBK1的表达。
新型冠状病毒与PRR信号传导相关的一个独特方面是,该病毒诱导NF-κB活化,尽管其参与抗病毒信号传导(方框2)。这种活性的基础被认为与NF-κB依赖的转录输出有关,这些转录输出与细胞防御无关,但却是病毒所需要的。此外,由于新型冠状病毒特异性靶向IRF3,激活的NF-κB信号不足以恢复IFN-I诱导。据报道,S、Nsp3和Nsp5病毒蛋白都通过各种提出的模型降低了IRF3的表达。Nsp5、Nsp12、ORF3b和M也阻断PRR信号事件后的IRF3的核转运。对于Nsp5和Nsp12,这种抑制不依赖于IRF3的磷酸化状态以及Nsp5和Nsp12各自的蛋白酶和聚合酶活性。也报道了Nsp1对IRF3磷酸化的抑制,尽管其机制可能涉及更普遍的整体翻译的关闭,这耗尽了该过程发生所必需的细胞因子,如下面进一步讨论的。
方框2新型冠状病毒和NF-κB之间的奇怪关系
细胞的生产性感染是一个决定性的时刻,通常会决定病毒-宿主关系的轨迹。宿主的成功通常等同于该细胞识别感染并启动两种互补策略的能力:由IFN-I或IFN-III介导的“召唤武器”和由促炎细胞因子和趋化因子介导的“召唤增援”。这两种细胞防御的启动需要连接模式识别受体(PRR)传感器,随后激活核因子-κB (NF-κB)和干扰素调节因子3 (IRF3)和/或干扰素调节因子7 (IRF7)。尽管在新型冠状病毒感染期间,各种病毒介导的机制阻断了PRR生物学,但如单细胞ATAC测序所示,NF-κB反应元件仍然活跃。结果,建立了不平衡的免疫反应,其中需要NF-κB和IRF3和/或IRF7的IFN-I或IFN-III信号被削弱,而由NF-κB单独控制的趋化因子介导的“对加强的需求”被维持。这个结果看起来可能会适得其反,因为它与病毒适应性和进化选择有关,但是发现抑制NF-κB不仅阻断趋化因子的产生,而且阻断新型冠状病毒的复制,这表明转录输出肯定也有益于病毒。尽管还不清楚哪些特定的NF-κB基因靶点是确保病毒复制的必要条件,但这些研究确实证明了信号传导是维持的。另一项探索NF-κB和新型冠状病毒之间关系的研究报道,ORF8作为IL-17A的病毒模拟物,结合并诱导人IL-17受体亚单位的异二聚化,并刺激NF-κB的下游活化。ORF8因此能够在感染过程中引发强烈的炎症反应,并被认为有助于新冠肺炎期间的严重炎症。患者感染的新型冠状病毒分离株含有缺失或突变ORF8据报道经历了较轻的疾病。Nsp14也被认为有助于NF-κB的病毒活化,这通过p65的核转位和促炎性趋化因子IL-6和IL-8的上调来测量。类似地,新型冠状病毒的刺突蛋白激活NF-κB的方式依赖于S1亚单位,并与NLRP3炎症小体表达增加相关。炎症小体是细胞内的多蛋白复合物,响应病原体相关的分子模式而聚集,并与炎症级联反应的激活有关。总的来说,新型冠状病毒和NF-κB信号之间的这种奇怪关系是病毒发病机制的独特驱动因素,其中不平衡的免疫反应和相关并发症是大量发病率和死亡率的原因。
阻断干扰素信号
新型冠状病毒感染细胞的单细胞RNA测序显示,在感染细胞中发现的总mRNA的60%以上可以是病毒来源的,这说明了病毒侵占细胞的效率。这一结果是高水平的病毒基因组RNA和sgRNA转录的产物,再加上Nsp1介导的宿主mRNA的抑制。Nsp10和Nsp14介导的宿主翻译和RNA剪接的靶向抑制进一步促成了这种敌意细胞接管。然而,即使有了这些有效的病毒对策,新型冠状病毒最终通过多种机制诱导细胞死亡,使得吞噬细胞能够检测到病毒。正是这种动态可能导致了在对新型冠状病毒感染的反应中观察到的高水平IFN-I和IFN-III,特别是在严重新冠肺炎患者中。由于这种生物学不容易被阻断,新型冠状病毒也受益于靶向负责响应IFN-I和IFN-III并诱导ISG表达的信号级联(图.2)。
新型冠状病毒Nsp13和Nsp14通过降低IFNAR1受体亚单位的表达来干扰IFN-I信号传导。此外,Nsp13和S蛋白都与STAT1相互作用,阻止其与受体对接和随后的磷酸化介导的激活。类似地,由Nsp1、Nsp10和Nsp14介导的宿主转录物的减少也有助于阻断ISG的产生。干扰ISG诱导对病毒来说尤其重要,因为用IFN-I或IFN-III引发的细胞感染是无效的,并且在病毒仍易被检测到时会放大宿主的整体反应。
阻断核运输
无论是通过阻断转录因子的转移还是通过阻止宿主mRNA的输出,干扰核转运机制都可以为细胞质病原体提供直接的选择优势。核孔复合体(NPC)是一个大的结构,它连接核被膜的内膜和外膜,并形成一个水通道,通过该通道调节核质运输。NPC由称为核孔蛋白(Nups)的各种蛋白质亚单位组成,它们与负责在细胞核和细胞质之间穿梭特异性标记蛋白质的核外输蛋白家族(输入蛋白和输出蛋白)的可溶性核转运受体相互作用。来自许多不同家族的病毒利用这种机制来允许特定的病毒蛋白根据需要进入细胞核,和/或阻断宿主成分执行限制病毒复制的功能134。对新型冠状病毒和宿主因子之间发生的蛋白质-蛋白质相互作用的分析表明,Nsp1、Nsp9、Nsp15、ORF6和M都能够与宿主核转运机制相互作用(图.3)。

图3:新型冠状病毒介导的一般宿主细胞生物学的干扰。抗病毒反应的细胞诱导依赖于通过核孔复合体的双向运输(NPC)。在核转运受体(importins)的帮助下,在感染应答中诱导的细胞转录因子通过NPC易位进入细胞核,并结合抗病毒基因内的序列以驱动其表达。新生信使RNA (mRNA)重新转录和加帽后,宿主剪接体在RNA剪接位点组装,促进内含子切除,产生具有翻译能力的成熟转录物。这些转录物与核转运受体(输出蛋白)结合,并通过NPC输出到细胞质中,在细胞质中,它们被宿主核糖体翻译,并根据信号识别颗粒(SRP)对其信号肽的识别进行适当的折叠和细胞定位。新型冠状病毒编码几种阻断核转运的蛋白质(用紫色表示),包括非结构蛋白(Nsps)、开放阅读框6 (ORF6)辅助蛋白和膜结构蛋白(M)。这种抑制作用是通过与所示的宿主蛋白(KPNA2,核转录因子亚单位α2;KPNA6,核蛋白亚基α6;NTF2,核转运因子2;Nup62,核孔蛋白62;Nup98,核孔蛋白98;NXF1,核RNA输出因子1;NXT1,核转运因子2-样输出因子1;Rae1,核糖核酸输出因子1)。新型冠状病毒还编码最终通过抑制宿主RNA剪接、优先阻断宿主RNA进行核输出而支持病毒RNA、干扰核糖体功能和阻止蛋白质运输来关闭翻译的蛋白质(用红色表示)。由于Nsp1也能够更普遍地抑制蛋白质合成,它在病毒RNA优先靶向中的作用仍然不确定(问号)。
许多独立的研究小组已经证明,ORF6通过与IFN诱导的核孔蛋白、Nup98和核糖核酸输出因子1 (Rae1)形成的复合物结合,并使它们远离NPC,从而抑制核质转运。这种相互作用的晶体结构数据分析表明,ORF6比RNA更容易进入NUP 98–RAE 1复合物的mRNA结合沟。通过靶向这种复合物,ORF6阻断转录因子的核转位,包括IRF3和ISGF3的单个组分,并阻止mRNAs,如编码IFN-I、IFN-III或抗病毒ISG的mRNAs进入细胞质进行翻译。值得注意的是,ORF6也可能有助于SARS-CoV和新型冠状病毒之间的可观察到的差异,因为来自新型冠状病毒的ORF6产生与比来自SARS-CoV的ORF6更强的抗IFN活性相关的修饰结构。
关闭翻译
除了阻止被感染的细胞利用自身防御系统的资源,许多病毒还瞄准蛋白质合成方面来增强适应性。在真核翻译过程中,成熟mRNA转录物的5′帽和poly-A结构被40S核糖体亚单位识别,这一过程受真核起始因子调节。在扫描AUG起始密码子时,60S核糖体亚单位加入复合物形成80S核糖体,多肽链开始延伸。由于病毒蛋白的翻译也依赖于宿主机制,病毒介导的对这些过程的干扰必须保持一定的选择性才能成功。
Nsp1是关闭宿主翻译的冠状病毒蛋白的最典型例子,尽管来自不同病毒的等同蛋白似乎使用不同的抑制策略。几个小组已经证明新型冠状病毒Nsp1通过结合40S核糖体亚单位的18S核糖体RNA成分阻断了mRNA进入核糖体的通道(图.3)。事实上,发现Nsp1的C末端与两种已知的核糖体抑制剂SERBP1和Stm1结构相似,这两种抑制剂也阻止mRNA进入40S核糖体的进入通道。此外,当Nsp1的C-末端结构域被突变以消除其与核糖体的相互作用时,观察到翻译阻遏的丧失。Nsp1和核糖体之间的这种相互作用与人类细胞中内源性蛋白质翻译的强烈减少有关,即使在缺乏其他新型冠状病毒蛋白的情况下。尽管一些小组已经报道了含有5’病毒前导序列的mRNAs在很大程度上被保护免受Nsp1介导的翻译抑制,其他人认为这一活动是非歧视性的。在这些研究中,新型冠状病毒的5’和/或3’非翻译区被融合到一个报道基因上,因此病毒mRNA的其他特征可能在逃逸翻译关闭中起作用。另外一项研究表明,包含病毒5’非翻译区导致报道基因mRNA的翻译增加了5倍,表明病毒RNA的翻译可能比宿主mRNA更有效,从而补偿了翻译阻断。或者,Nsp1蛋白可以诱导缺乏5’病毒前导序列的mRNA的降解,从而允许病毒mRNA选择性地翻译超过细胞mRNA。由于Nsp1蛋白与宿主先天免疫逃逸相关的几个过程有关,因此考虑其全面抑制蛋白翻译的能力如何促进其其他已报道的功能是很重要的。
除了充分证明的Nsp1的参与,其他病毒蛋白如Nsp10、Nsp14和Nsp16也在新型冠状病毒感染期间的翻译关闭中发挥作用(图.3)。为了使新生RNA转录物转化为具有翻译能力的成熟mRNA,必须切除插入序列(内含子),以便表达的序列(外显子)可以连接在一起。这一过程被称为RNA剪接,由称为剪接体的RNA-蛋白质复合物介导,剪接体由一组小的非编码核RNA和蛋白质剪接因子组成。由于剪接的破坏通常等同于不需要这种生物学的病毒的选择性优势,这是RNA病毒的另一个常见观察到的目标,包括新型冠状病毒。在这方面,新型冠状病毒Nsp16已被证明能够结合U1和U2的mRNA识别域,这两种主要的小核RNA亚单位组成了剪接体,导致感染期间宿主mRNA剪接的全面抑制。据报道,Nsp10和Nsp14对全局宿主表达的类似干扰也会影响剪接。在来自新冠肺炎患者的受感染肺样本中观察到新型冠状病毒感染导致宿主剪接活性的扰动。
为了确保新形成蛋白质的正确折叠和运输,信号识别颗粒(SRP)不断扫描新生核糖体相关肽链,寻找疏水性信号肽,以识别旨在分泌或插入宿主膜的产物。在信号识别时,SRP触发核糖体转运到内质网,在那里这些蛋白质可以正确地形成和传递。已经发现新型冠状病毒Nsp8和Nsp9各自与SRP复合物的7SL RNA支架成分结合,破坏蛋白质运输并导致新翻译的蛋白质降解。具体来说,Nsp8结合到由SRP54蛋白结合的7SL区域,SRP 54蛋白是SRP负责信号肽识别的成分,而Nsp9结合到与SRP19蛋白相互作用的7SL区域,这是SRP自身正确折叠和装配所必需的。这种对SRP结构和功能的干扰与SRP依赖性膜蛋白整合入细胞膜的蛋白减少有关。由于许多免疫介质,包括IFN-I和IFN-III家族成员,都以这种方式分泌,这项研究表明Nsp8和Nsp9在逃逸宿主免疫中起着重要作用(图.3)。
全身炎症和临床表现
尽管新型冠状病毒在直接感染细胞的情况下具有阻断和逃逸先天免疫的能力,但这些免疫机制在未感染的旁观者细胞中保持完整,这些旁观者细胞能够感应和响应来自死亡或受损细胞的碎片。具有讽刺意味的是,这种动态导致异常强大的先天免疫反应,尽管有所延迟,因为新型冠状病毒复制最初可以在气道中有增无减,直到受感染的细胞开始死亡,并释放出数千个新的子代病毒体。因此,当病毒PAMPs变得易于检测时,宿主反应的初始参与在相对高的病毒载量时开始。这种动态导致IFN-I和IFN-III的产生增加,其扩散到气道之外,并开始在所有远端组织中诱导炎症过程。即使在没有病毒感染的情况下,这种全身性反应也会给身体带来巨大压力,并表现为与新冠肺炎相关的各种临床表现。
可以说,新型冠状病毒生物学的一个悖论是,这种呼吸道病毒能够引起肺外临床表现,特别是胃肠道和/或心血管表现。无数临床案例研究记录了急性感染期间身体每个器官中与病毒相关的转录变化,这种现象可以在新冠肺炎仓鼠模型中复制。此外,相当一部分清除了病毒的新冠肺炎患者可能会保留涉及多个器官的长期疾病症状。尽管对新冠肺炎急性和持续表现的潜在生物学的理解仍不完全,但宿主对新型冠状病毒感染的反应无疑启动了这些过程。
人们很容易推测,新型冠状病毒多样的临床表现可能是该病毒某些独特方面的产物。尽管许多记录在案的系统性疾病的子集可能是真实的,但应该注意的是,在对其他呼吸道感染(如甲型流感病毒(IAV))的反应中,也经常观察到与新冠肺炎相关的几种疾病结果,这表明疾病的程度可能是病毒特异性属性(如病毒保真度或干扰)的副产品,这可能影响炎症物质的产生或可感知该物质的动力学。这一概念得到了最近出版物的支持,这些出版物表明,用新型冠状病毒或H1N1 IAV毒株感染不同的小动物模型可以在远离初始感染位点的器官中产生可比较的转录特征。在感染了任何一种病毒的非免疫缺陷个体中,在气道外分离出传染性病毒是罕见的,这表明病毒介导的气道损伤导致了大量的宿主反应,该反应通过循环并刺激每个器官系统。由于这些器官中的任何一个的潜在状况都可能因这种刺激而恶化,因此可以预见到不同的临床表现。
与观察到的宿主对IAV和新型冠状病毒的免疫反应相似性相反,一些临床表现表明新型冠状病毒确实表现出独特的属性,特别是其诱导急性后遗症(也称为长新冠)的能力。虽然IAV和新型冠状病毒制定了类似的策略来获得细胞通路,但负责这些活动的病毒融合蛋白利用了不同的细胞受体。由此产生的细胞嗜性差异有助于解释新型冠状病毒导致一些与长新冠相关的疾病的倾向,包括嗅觉缺失(嗅觉丧失),这是一种常见的临床表现和新冠肺炎的独特特征。新型冠状病毒有效地感染嗅觉上皮中的支持细胞,导致与在气道中观察到的相同的宿主反应和细胞死亡。病毒感染后,有证据表明,濒死的支持细胞释放载有dsRNA的物质,能够触发小胶质细胞和巨噬细胞的激活以及嗅觉系统中抗病毒防御的有力参与,这些事件在对IAV感染的反应中没有观察到。这些旁观者细胞可能产生的IFN-I、IFN-III和促炎细胞因子随后可以在嗅觉系统中诱导免疫引发,其动态类似于呼吸道感染如何引发远端器官。然而,嗅觉系统中的一个重要区别是,当嗅觉神经元对这些类型的免疫提示做出反应时,它们的正常功能被破坏,这可以解释嗅觉缺失的发展。同样的现象也可能与新型冠状病毒感染后其他神经功能障碍的潜在原因有关。总之,这些数据表明,给定病毒感染的病理生物学可由可用于检测的炎性物质的量,结合该物质产生和/或沉积的物理位置来定义。
新冠肺炎的宿主靶向免疫疗法
RNA病毒种群经历对数扩张,并在此过程中产生小突变景观,使它们能够逃脱作者的免疫防御和/或抗病毒药物施用所施加的选择压力。避免这种逃逸的一个策略是靶向病毒生物学所需的宿主因子,而不是宿主生物学所需的因子。例如,通过全基因组CRISPR-Cas筛选,已经确定参与但不是关键的膜运输、溶酶体调节和染色质重塑的宿主因子是新型冠状病毒感染的关键。因此,设计用来竞争性结合这些因子的小化合物,会对病毒施加强大的选择压力。一种补充方法是直接针对宿主对病毒感染的反应。这种策略的常见例子包括减轻疼痛、发热和/或不适的化合物。此外,使用免疫调节化合物可以减少由宿主对病毒感染的不平衡反应引起的发病机理。下面,作者总结了已经证明在治疗新冠肺炎住院患者中具有临床价值的宿主靶向免疫疗法(表2)。
表2美国国立卫生研究院推荐的用于新冠肺炎治疗的免疫调节小组
种类 | 名字 | 通用机制 | 建议a |
消炎的 | 秋水仙碱 | 降低中性粒细胞的趋化性 抑制炎症体信号传导 减少炎性细胞因子的产生 | 不推荐,除非在临床试验中 |
消炎的 | 氟伏沙明 | 减少炎性细胞因子的产生 | 证据不足 |
抗炎:皮质类固醇 | 地塞米松(全身) | 炎症的全身缓解 | 用于需要机械通气或氧气支持的新冠肺炎住院患者 |
抗炎:皮质类固醇 | 强的松 甲基强的松龙 氢化可的松 | 炎症的全身缓解 | 在无法获得地塞米松的情况下,用作地塞米松的替代品 |
抗炎:皮质类固醇 | 布地奈德(吸入) | 炎症的局部缓解(肺) | 证据不足 |
细胞因子抑制剂 | Canakinumab | 抑制白介素-1 | 证据不足 |
细胞因子抑制剂 | Siltuximab | 抑制白介素-1 | 不推荐,除非在临床试验中 |
细胞因子抑制剂 | Tocilizumab | 抑制白介素-6 | 不推荐,除非在临床试验中 |
细胞因子抑制剂 | Sarilumab | 抑制白介素-6 | 与全身性皮质类固醇联合使用,用于需氧迅速增加且炎症标志物增加的重度新冠肺炎住院患者 |
细胞因子抑制剂 | 萨里卢单抗 | 抑制白介素-6 | 作为tocilizumab的替代药物,以防不可用 |
细胞因子抑制剂 | Gimsilumab Lenzilumab Namilumab Otilimab Mavrilimumab | 抑制粒细胞-巨噬细胞集落刺激因子 | 证据不足 |
激酶抑制剂 | 巴里西替尼 | 抑制JAK1和JAK2 | 作为第二种免疫调节药物,用于需氧迅速增加和全身炎症的使用地塞米松的住院患者 |
激酶抑制剂 | 托法替尼 | 抑制JAK1、JAK2和JAK3 | 在无法获得或不可行的情况下,作为baricitinib的替代药物 |
激酶抑制剂 | 鲁索利替尼 | 抑制JAK1和JAK2 | 除了巴里西替尼或托法替尼之外,不推荐使用JAK抑制剂,除非是在临床试验中 |
免疫球蛋白 | 非新型冠状病毒特异性静脉免疫球蛋白 | 炎症反应的一般抑制和/或改变 | 不推荐,除非在临床试验中 |
00001. JAK,杰纳斯激酶。a国立卫生研究院新冠肺炎治疗建议
皮质甾类
多项随机对照临床试验(RCT)证明了系统性皮质类固醇治疗对一部分患有严重新冠肺炎的住院患者的有效性,这可能是由于其能够总体缓解新型冠状病毒诱导的炎症。在大(n= 6,425)、开放性RCT(恢复)、低剂量地塞米松降低了接受有创机械通气或单独吸氧患者的死亡率,但未降低未接受呼吸支持的患者的死亡率。一项包括1703名新冠肺炎重症患者的七项随机对照试验的世卫组织荟萃分析也报告了使用地塞米松后死亡率的降低。由于这些发现,目前的NIH指南推荐在需要机械通气或氧气支持的新冠肺炎住院患者中标准使用地塞米松或其他全身性皮质类固醇。
白介素-1和白介素-6抑制剂
表征宿主对新型冠状病毒反应的早期研究强调了威胁生命的过度炎症反应的发展,其特征在于IL-1和IL-6的强烈诱导。这一发现与细胞因子释放综合征相似,细胞因子释放综合征是一种异常的危及生命的免疫反应,通常与各种癌症治疗有关。因此,对IL-1抑制剂(阿那金拉和卡纳金单抗)或IL-6抑制剂(妥珠单抗和沙立单抗)进行了单独测试,以评估其对新冠肺炎的治疗价值。尽管一些临床试验报告了炎症生物标记物的改善和肺部病理学的改善,但其他临床试验并不确定或仅证明了在减少住院后更严重疾病进展方面的适度价值。尽管没有足够的证据支持使用IL-6抑制剂进行单一治疗,但在需要通气的住院患者中使用皮质类固醇治疗在临床上是有益的。正在努力评估同时阻断IL-1和IL-6途径的价值(Clinicaltrials.gov;NCT05279391)189。
JAK抑制剂
作为对新型冠状病毒感染的反应,通过细胞因子介导的Janus激酶(JAK)诱导STAT依赖性信号事件,触发了多种免疫相关活动。这一途径的治疗性抑制剂的子集已显示出对新冠肺炎住院患者的治疗有益。COV屏障RCT(n= 1,525),该研究观察了患有新冠肺炎肺炎和至少一种炎症标志物升高的患者,证明在标准治疗中与皮质类固醇一起使用时,baricitinib (JAK1和JAK2抑制剂)的生存率增加。在ACTT-2 RCT(n= 1,033),尽管该药物未与皮质类固醇联合使用。此外,对289名使用低流量氧气的住院患者进行的双盲、安慰剂对照STOP-COVID RCT研究显示,托法替尼(JAK1、JAK2和JAK3抑制剂)与地塞米松联合使用可降低呼吸衰竭和死亡的风险。
目前,针对主要病毒蛋白酶的直接抗病毒药物(Paxlovid,由辉瑞公司生产)是高危、有症状的新型冠状病毒患者的首选治疗方法。如果疾病继续发展,需要住院治疗,最初的努力集中在预防凝血,同时用廉价的类固醇减少患者的全身炎症。上述免疫调节药物,以及其他仍在评估中的药物,可以作为标准治疗无效时可能使用的实验方法。
结论
新冠肺炎疫情对全球健康造成了重大负担,其整体程度仍不清楚。尽管主要局限于气道,新型冠状病毒感染诱导全身性先天免疫激活,这在临床上可表现为多种肺外表现以及更常见的呼吸系统并发症。除了全世界无数的感染和数百万人死亡之外,新型冠状病毒还具有独特的能力,在感染后数周至数月内产生持续症状——这是一种俗称为长期COVID的临床现象。基于作者目前对新型冠状病毒生物学的理解,许多急性疾病结果的潜在驱动因素已经变得更加清楚,而其他的,尤其是那些与长新冠相关的,仍然是谜。
呼吸道中的初始感染导致大量病毒复制,将受感染的细胞转化为病毒工厂,并阻碍减缓这一过程所需的一线IFN介导的防御。让事情变得更复杂的是,新型冠状病毒还让细胞参与促进亲病毒环境,这顺便激活了后期先天免疫防御的一些方面。这种不平衡的反应导致促炎细胞进入呼吸道,同时病毒复制不受抑制,导致广泛的炎症和呼吸系统损伤。随着受感染细胞的死亡,与病毒相关的炎症物质开始出现在细胞外环境中,使得能够通过局部旁观者细胞诱导IFN应答。随着病毒群体的扩大,炎症碎片的积累和检测加速,导致IFN信号可以在身体的每个器官中被记录。这种反应可以在感染后持续7-10天,对远端感染提供一些保护,但如果存在任何潜在的情况,也可能导致器官功能障碍。在健康个体中,感染的急性期可以实现而几乎没有后果,并且导致成功的病毒中和和感染的消除。
尽管最初的病毒-宿主动力学现在已被普遍理解,但新型冠状病毒感染的长期后果仍不清楚。长新冠包含无数没有明确根本原因的临床表现。为了解长新冠所做的初步努力已经发现了嗅觉系统中局部和持续炎症、大脑区域变化和血液异常的证据。当在小动物中建模时,可比较的观察进一步表明,这些事件可以导致感官知觉、神经生物学、肾功能和心血管性能的变化。尽管这些疾病的许多表现可能是随机的或由共同机制驱动的,但目前作者仍不了解持续炎症的基础。在动物模型和尸体组织中观察到的转录变化表明,能够诱导炎症的物质在病毒清除后仍然存在。目前关于这种物质来源的主要理论包括低水平病毒复制、持续缺陷的病毒基因组,甚至是由最初的新型冠状病毒病毒感染造成的二次感染。可能是所有这些可能性,或者没有,驱动长新冠。尽管如此,解决这一科学问题应该是社区向前发展的中心焦点,因为这对于为继续受新型冠状病毒影响的许多生命开发有效的治疗方法至关重要。
Minkoff JM, tenOever B. Innate immune evasion strategies of SARS-CoV-2. Nat Rev Microbiol. 2023 Mar;21(3):178-194. doi: 10.1038/s41579-022-00839-1. Epub 2023 Jan 11. PMID: 36631691; PMCID: PMC9838430.
https://blog.sciencenet.cn/blog-55647-1377958.html
上一篇:354.新型冠状病毒病毒载量和脱落动力学
下一篇:356.尼日利亚阿布贾自报被犬咬伤的受害者中与人类狂犬病死亡概率相关的可变因素建模