博文
良性氧化应激和氧化还原稳态  精选
精选
|
原文作者:赫尔穆特·西斯
德国杜塞尔多夫海因里希-海涅大学医学院生物化学与分子生物学研究所

Sies H. Oxidative eustress: On constant alert for redox homeostasis. Redox Biol. 2021;41:101867.
在开放代谢系统中,氧化还原相关的信号传递需要对稳态氧化还原设定点进行持续监测和微调。正在进行的氧化代谢是一个持续的挑战,被称为氧化应激,它在生理范围内运行,被称为“内稳态空间”、“金发姑娘区”或“黄金均值”。通过隔离生成和去除氧化剂来实现氧化还原信号传递的空间和时间控制。过氧化氢是主要的氧化还原信号分子,其细胞景观的特点是器官之间的浓度差异达到数量级。这种浓度模式与氧化修饰蛋白质的模式相对应,例如S-谷胱甘肽化蛋白质。该综述介绍了短期(非转录)和长期(转录/翻译)内稳态机制以及应激和应激反应的概念背景。氧化还原设定点是一个可变的移动目标值,受昼夜节律和外部影响(统称为暴露组,包括营养和生活方式因素)的调节。简要介绍了生理环境中细胞特异性和组织特异性氧化还原调节的新领域,包括对胚胎发育和寿命、骨骼肌和运动、睡眠-觉醒节律以及神经系统功能(导致心理生物学方面)中氧化应激作用的新认识。
一、前言
维持氧化还原平衡是一个持续的挑战。恒定的监测是建立和维持氧化还原平衡的标志,由于其潜在的动态性质,更精确地称为“内稳态动力学”。最近在氧化还原生物学领域的研究进展揭示了一种生理功能的氧化还原结构,它按照一组被称为“氧化还原密码”的原则进行组织。其中一项原则是氧化还原代谢的激活/失活循环,特别是涉及过氧化氢的循环,与其他分子信号因子一起支持细胞和器官分化和生命周期的空间时间序列。氧化还原信号的更广泛领域已经得到了广泛的综述。
细胞氧化还原动力学与分子和细胞水平的复杂结构事件密切相关。后者通过超分辨率纳米显微镜揭示的线粒体内膜嵴的持续重塑来说明,这种重塑以秒为单位发生。这种重塑反过来又与线粒体超级复合物的形成变化有关。基本的分子监测事件包括氧化还原参数,这些事件的时间尺度比秒的范围要短得多,先于结构变化发生。过氧化氢对线粒体氧化还原网络的稳定性有贡献,并且在细胞及其器室水平存在一个集成的氧化还原网络来监测内稳态。通过在“氧化应激”中不断监测氧化还原活性来维持内稳态。
在这里,我们从氧化还原内稳态的概念背景开始,描述了代谢稳态的特征。接下来是关于过氧化氢的细胞氧化还原景观的一般快照,这是中心氧化还原信号代谢物。将概述短期(非转录)和长期(转录/翻译)机制。我们将介绍新兴的氧化还原研究领域的例子,重点关注昼夜节律、睡眠-觉醒周期、胚胎发育和寿命、骨骼肌和运动,以及涉及心理生物学中分子关系的神经系统的某些方面。
Oxidative eustress: On constant alert for redox homeostasis - ScienceDirect
二、开放系统和稳态维持
冯·贝塔兰菲(Von Bertalanffy)开创了流动平衡的生物物理学,也被称为“稳态”:“生命系统是开放的系统,通过与环境的物资交换来维持自身,并不断构建和分解其组成部分”。开放的代谢系统需要持续监测流入和流出,以最小化与稳态设定点的偏差。在氧化还原调节中,这指的是生理性氧化应激或氧化应激。从设定点出发的生理范围的偏移被称为“内稳态空间”、“金发姑娘区”和“黄金分割”。
此外,不断变化的代谢条件需要适当调整稳态设定点,即在时空背景下的还原-氧化目标值。这被塞尔耶(Selye)的适应性压力概念所体现。普里高津(Prigogine)分析了时间结构和波动,他将其称为非平衡热力学中的“耗散结构”。在生物学中,克劳德·伯纳德(Claude Bernard)的“内环境”概念在“韧性”和“异位调节”等术语中找到了吸引力。韧性表示恢复到原始状态的能力,即反弹能力,而异位调节则是指通过改变到新的设定点来实现稳定,这是一个矛盾修饰法。一个相关的术语是戴维斯(Davies)提出的“适应性内稳态”。
因此,维持内稳态的机制可以分为反应性(反馈、反向调节)和预测性(前馈、预期)模式。
至于后者,对内源性和外源性(“暴露组”)刺激的预适应能力,总结为“毒素激活效应”和更具体的“线粒体激活效应”,对于适应性压力反应非常重要。图1给出了压力和压力反应的概念的时间轴。
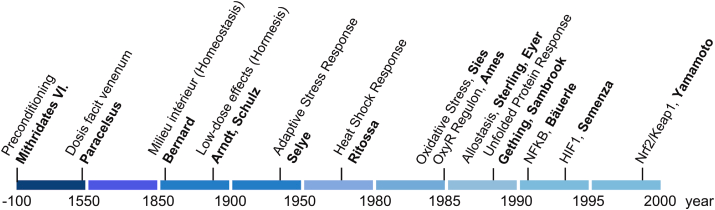
图1 压力和适应性压力反应的概念时间轴。
米特里达梯六世(Mithridates VI)和帕拉塞尔苏斯(Paracelsus)早期就认识到剂量在决定有益还是有害结果方面很重要。伯纳德的“内环境”概念被称为“内稳态”,而阿恩茨-舒尔茨(Arndt-Schulz)规则被称为“毒素兴奋效应”。20世纪带来了适应性压力综合症、热休克反应、氧化应激、OxyR、异位调节、未折叠蛋白反应以及主要的哺乳动物主调节因子NF-kB、HIF1和Nrf2/Keap1等。
塞尔耶于1975年引入了良性应激和应激的区别,近年来分子层面上的氧化应激引起了人们的兴趣,用来表示生理上的、无害的氧化应激。
三、时间空间控制
细胞组织的特点是亚细胞隔室化和梯度分布。有趣的是,快速作用的非转录信号因子以大约nM浓度存在;如表1所示,过氧化氢、Ca2+和pH值也在这个范围。
(可以指出,pH=7表示[H+]=100 nM H+。)其他重要的信号实体,如NO和H2S也以nM生理浓度存在,构成了一个“活性物种相互作用网络”。此外,通过活性亲电物进行氧化还原信号传导以维持亲核性。
表1:快速作用的信号因子过氧化氢和Ca2+以纳摩尔浓度存在。
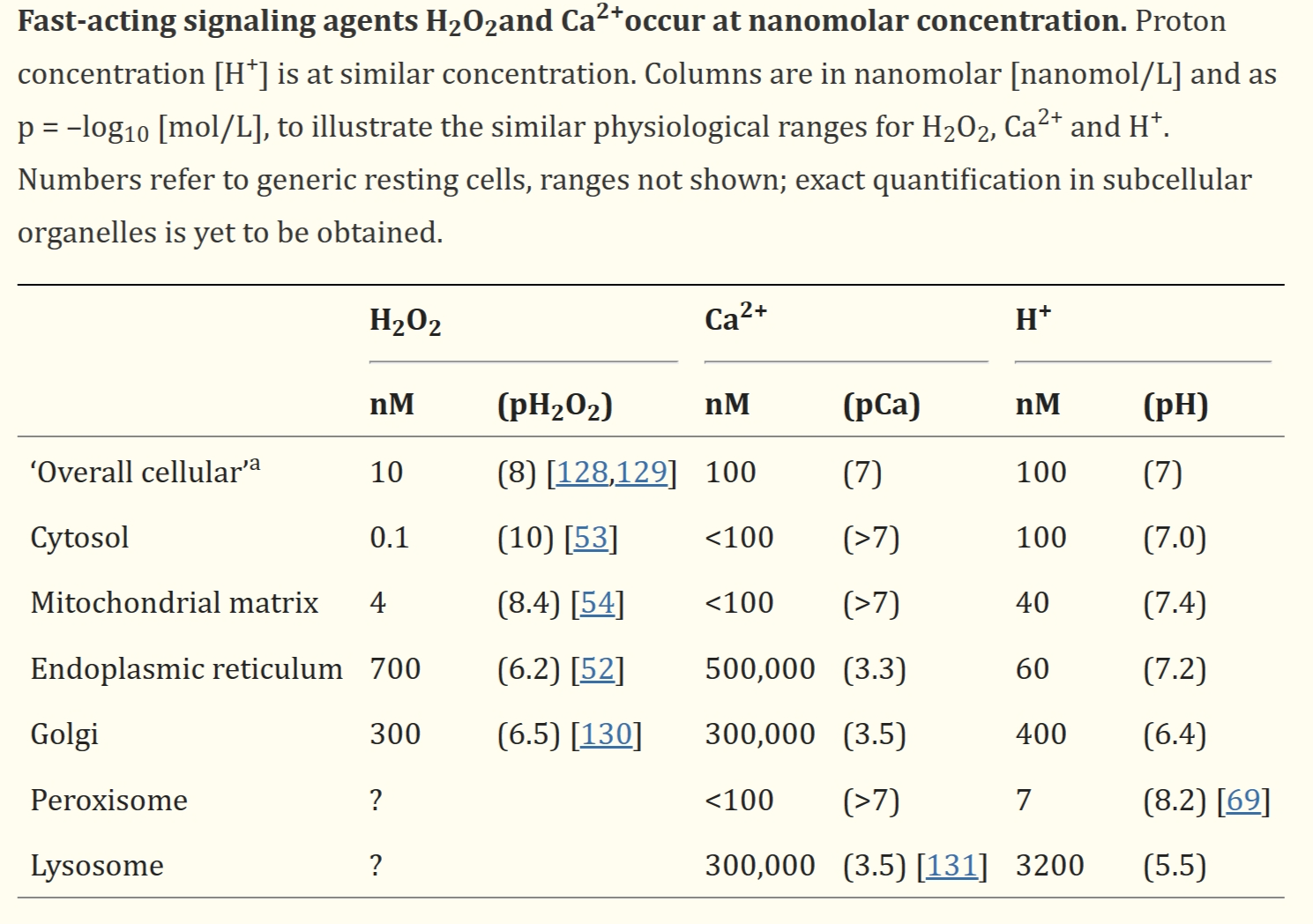
[Ca2+]的“缓冲”依赖于内质网和线粒体基质中的Ca2+储存,而[H+]的缓冲依赖于碳酸酐酶的作用和呼吸。相比之下,细胞内没有过氧化氢的“缓冲储存”。在这里,对[过氧化氢]的控制依赖于酶促合成和降解过氧化氢的快速微调以及梯度控制。已经提出了线粒体作为“活性氧稳定设备”的作用,这也扩展到由NADPH氧化酶等胞外来源的控制。
过氧化氢和Ca2+在许多信号过程中相互关联。如表2所示,一个典型的例子是瞬时受体电位(TRP)通道,它们作为氧化还原环境刺激的生物传感器,被过氧化氢、NO和亲电物激活。这些通道促进Ca2+内流,触发细胞反应。受体具有高度反应性的半胱氨酸(TRPA1中的C621),其修饰会导致门的打开。另一个例子是STIM2中半胱氨酸313的氧化抑制钙库操纵的钙通道。
表2:生命过程中生理性氧化应激(良性应激)的一些示例。

3.1. 空间:细胞氧化还原景观
根据目前可用的信息,过氧化氢在细胞中的浓度模式如图2所示,尽管成像的校准数字有限(见表1),但颜色尺度仍然很丰富。极端情况从内质网腔开始,接近μM 过氧化氢范围,向下到较低的一端,计算得出低至80 pM 过氧化氢。估计线粒体基质中含有约4 nM 过氧化氢。我们对于过氧化物酶体和溶酶体的相应数量仍然知之甚少。实验发现过氧化物酶体基质过氧化氢浓度远高于线粒体基质过氧化氢,但在roGFP2-Orp1实验中给过氧化物酶体数据赋值是困难的。图2给出了一个粗略的方向;显然,存在具有不同稳态分布的过氧化氢亚细胞μM或nM,表明该景观多样化并在接收到提示时微妙地重新适应。因此,由于已知的名为细胞器“接触学”的复杂的器官间关系,更精细的景观将更加复杂。
图2.过氧化氢的细胞“风景。
在亚细胞空间中估计过氧化氢浓度的一般概述。颜色从浅蓝色 (80 pM) 到蓝绿色 (4 nM)、绿色 (20 nM)、棕色 (300 nM) 和红色 (700 nM)。
值得注意的是,过氧化氢的“景观”类似于蛋白质S-谷胱甘肽化的模式:最近的研究显示,蛋白质被S-谷胱甘肽化的平均占据率在内质网中最高,而在线粒体基质中最低。因此,S-谷胱甘肽化蛋白质的亚细胞分布(图3)类似于图2中所示的过氧化氢浓度。将这种模式与其他类型的蛋白质氧化后转录修饰(oxPTM)进行比较将是有趣的,例如主要定位于线粒体的辅酶A,称为S-CoAlation,以及其他如氧化还原敏感蛋白质的S-过硫化、S-酰化、S-亚硝基化或S-棕榈酰化等。这些oxPTMs的功能前景正在阐明,例如对于S-谷胱甘肽化或S-CoAlation 。进一步的细化水平涉及蛋白质半胱氨酸巯基的反应性:谷胱甘肽过硫化物(GSSH)的pKa比谷胱甘肽(GSH)低约3.5个单位,使GSSH在生理pH下比GSH更具反应性。同样,在较碱性的8.2 pH值的微粒体基质中,硫醇化学的存在将比中性条件下更为普遍。因此,活性物质和位置的特殊性可以导致超过1000倍的反应性差异,说明了氧化还原化学与腔室pH之间的关系。
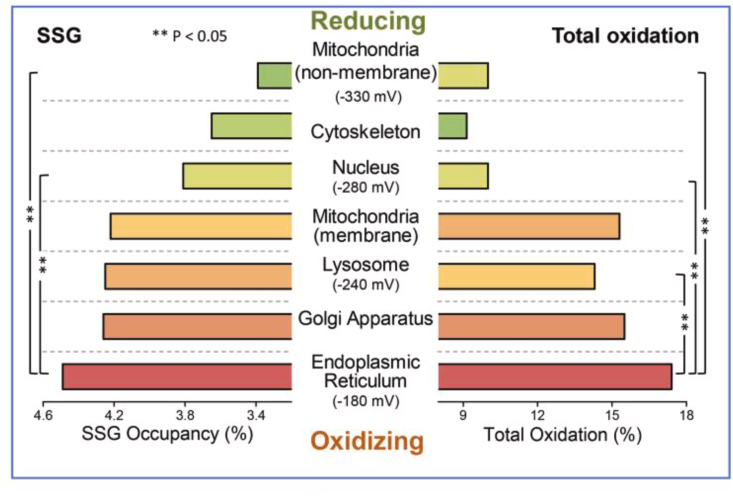
图3 蛋白s-谷胱甘肽化(SSG占用率)和总氧化的平均亚细胞分布。巨噬细胞的氧化还原蛋白质组分析。
3.2. 时间:氧化还原调定点是一个移动的目标
时间响应可以分为几秒钟范围内的即时氧化还原响应和更长的时间范围响应。在短时间范围内监测时,过氧化氢的来源,如NADPH氧化酶、线粒体呼吸链复合物和过氧化氢生成酶,会立即响应代谢和物理刺激而被激活。这不需要转录/翻译基因表达的激活,允许通过反馈环路实现对稳态设定点的紧密控制。一个典型的例子是伤口愈合的快速启动,它涉及过氧化氢、Ca2+和ATP等参与的无需转录的损伤信号,能根据生理需求立即执行。
较长时间范围的氧化还原响应在数小时范围内或更长,基于转录/翻译激活,允许前馈和反馈调节。其中一个主要特征是昼夜节律,即昼夜节律反应模式。转录/翻译反馈环路(TTFL)构成了哺乳动物生物钟的基础。过氧化物还原酶(peroxiredoxin)是昼夜节律的标志物,而过氧化物还原酶作为氧化还原中继枢纽的作用正在逐渐显现。过氧化物还原酶-6的周期性表达受到时钟蛋白Bmal1和Nrf2的共同调控。过氧化氢的昼夜振荡对CLOCK蛋白的氧化还原控制产生影响,这种影响是通过介导过氧化氢设定点在一天内的变化的适配器蛋白p66Shc实现的。因此,过氧化氢振荡与耦合的生物钟网络融合在一起,使过氧化氢调定点成为一个动态移动的目标。
昼夜节律广泛影响着生理功能。氧化还原对睡眠-觉醒模式的影响的分子基础开始被揭示。在果蝇中,发现Kv钾通道β亚基将线粒体氧化事件与睡眠联系起来。同样在果蝇中,观察到睡眠和氧化应激之间的双向关系;野生型果蝇睡眠增加能提高它们对氧化应激的抵抗力,而减少神经元中的氧化应激会缩短睡眠时间,这导致作者提出了这样的口号:“睡眠清除活性氧,活性氧促进睡眠”。睡眠不足会导致“活性氧”积累,而因睡眠不足导致的死亡已被归因于肠道中的氧化应激。这里对这个新兴研究领域的简要了解可能就足够了。
3.3. 梯度控制:过氧化物还原酶的作用
血浆中过氧化氢的浓度估计为1-5 μM。因此,从细胞外的血浆膜到细胞内的过氧化氢浓度之间存在一个陡峭的梯度,其在nM范围内(表1)。可以说,人们可以考虑将μM的细胞外过氧化氢浓度作为“过氧化氢储存库”,根据需求进行利用。同样,在亚细胞器和细胞质之间也存在大量的过氧化氢梯度。几种水通道(AQP3、AQP5、AQP8、AQP9、AQP11)促进了过氧化氢的跨膜扩散,因此水通道被更具体地称为“过氧蛋白”。关于AQP8的研究揭示了一种涉及半胱氨酸硫醇化的门控机制,表明过氧蛋白以氧化还原依赖的方式参与过氧化氢梯度控制。AQP11定位于内质网膜。研究发现,AQP11能有效地将过氧化氢转运到细胞质中,使其成为潜在的内质网基于氧化还原信号的调节因子。植物中的昼夜水孔蛋白表达很早就被发现了。虽然在哺乳动物细胞中,过氧蛋白是否经历昼夜节律尚未得到详细研究,但表皮中的AQP3已被发现经历这种节律。
(博主:细胞外过氧化氢浓度最高,延申的问题是,这些过氧化氢的来源是什么?细胞内过氧化氢浓度为什么低,气体通道不能建立细胞内外的平衡吗?外源性注射过氧化氢是否具有合理性,历史曾经使用过这种方法。至少在杀死厌氧菌方面有更大优势?)
四、功能:细胞和组织特异性氧化还原调节
细胞和组织通过氧化还原调节发挥其特殊的生理功能。特别是每个组织内的蛋白质-半胱氨酸氧化还原网络构成了组织特异性生物学的基础。图4回顾了氧化还原蛋白质组的一般概述,它通过动力学控制的巯基开关进行组织。蛋白质-半胱氨酸硫醇活性和静电门控赋予了特异性。使用一种名为“Oximouse”的方法在体内定量映射小鼠半胱氨酸蛋白质组已经开启了新的视野。对这种“景观”的测定揭示了特定蛋白质的氧化还原调节具有很强的组织特异性。Oximouse氧化还原网络有助于识别新的氧化还原调节途径。内质网感知和响应过氧化氢的途径已经被确定。
本节将介绍一些需要氧化应激的过程,参考了这些新兴领域的最新文献,但并未尝试全面覆盖这些快速发展的主题。在最近的一篇综述中,还讨论了其他重要主题,包括免疫系统、炎症和伤口修复、心血管系统、胰岛素敏感性和糖尿病的发病机制、衰老和癌症。
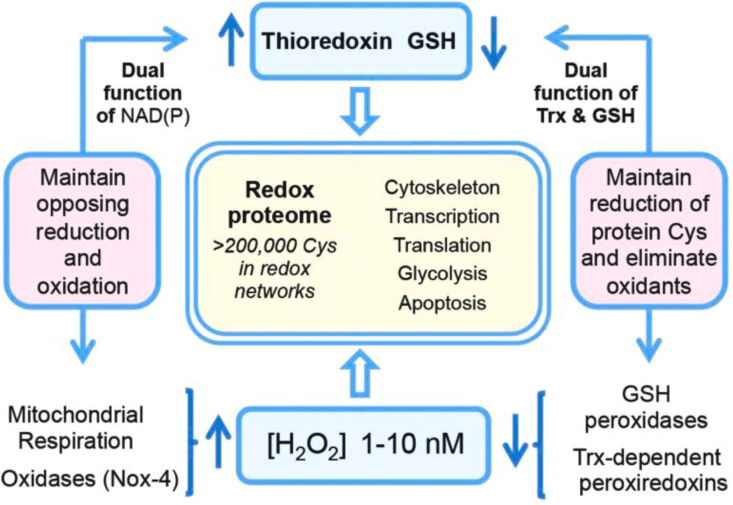
图4氧化还原蛋白质组通过动力学控制的巯基开关进行组织。
4.1. 发育和寿命
在胚胎发生和整个发育过程中,氧化还原信号对细胞命运决定和器官形成起着重要作用。胚胎的发育伴随着显著的氧化还原状态变化。过氧化氢参与形态发生和细胞分化,伴随着谷胱甘肽利用的变化。图5提供了斑马鱼胚胎在受精后48小时各个器官中过氧化氢浓度的模式快照,使用了遗传编码探针Hyper7。在这个阶段,过氧化氢浓度在发育中的神经系统(大脑、视网膜和脊髓)以及心脏中特别高。NADPH氧化酶在神经元发育中的作用已经得到证实。过氧化氢和同源蛋白Engrailed协同作用塑造斑马鱼的光窝,促进胚胎模式的形成,这扩展了早期关于过氧化氢在轴突生长锥路径寻找中的作用的发现。再生研究中也获得了关于活性氧在轴突生长中作用的信息。神经损伤后,NADPH氧化酶-2从巨噬细胞释放到外泌体中,然后通过胞吞作用被整合到受损的轴突中。作者提出了NOX2-PI3K-pAkt的信号传导途径用于轴突再生。
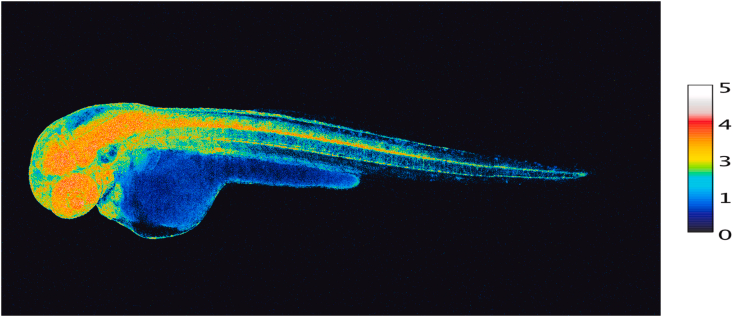
图5受精后48小时完整发育的斑马鱼胚胎各器官中过氧化氢浓度。
早期生活事件、活性氧敏感的表观遗传标记、抗压能力和寿命之间的分子联系已经得到确认。这些在秀丽隐杆线虫和HeLa细胞中观察到的效果,被归因于全身活性氧介导的特定发育组蛋白修饰的减少:组蛋白3(H3K4me3)的三甲基化降低,导致抗压能力增加。
干细胞的命运和功能与氧化还原稳态紧密相连,决定了静止、自我更新和分化之间的选择。此外,氧化剂在繁殖中的作用正在被认识。辅助繁殖中氧化剂的作用需要进一步阐明,以便在保持氧化还原信号的同时最大限度地减少氧化损伤。
4.2. 骨骼肌:运动和良性应激
过氧化氢 是骨骼肌生理学和适应运动的关键信号。无论是对体力活动的短期即时反应,还是长期的重塑和适应,都涉及由过氧化氢引起的氧化还原信号。研究发现,在肌肉收缩活动中,2-Cys过氧化物还原酶-2被迅速且可逆地氧化,因此确定这种蛋白质为肌肉氧化还原信号中的一种效应器。其他可能的效应器分子,如其他活性过氧化物还原酶,将在肌肉适应过程中发挥作用,而老年人锻炼反应减弱的原因可能是效应器如过氧化物还原酶-2的瞬时氧化减少。细胞外超氧化物歧化酶 (SOD3) 已被认为在运动期间减轻氧化挑战方面起作用。癌症中肌肉质量的损失(恶病质)与氧化还原稳态的改变有关。
4.3. 良性氧化应激、神经系统和心理生物学
大脑利用氧化还原信号执行许多功能,下丘脑是“主指挥家”。在神经元-星形胶质细胞界面存在氧化还原信号与代谢信号的交叉对话,星形胶质细胞和神经元在模式化硒蛋白方面存在紧密的关系,这些硒蛋白构成了氧化还原控制的一部分。突触可塑性、突触修剪和谷氨酸受体激活只是神经系统中氧化剂功能性使用的众多例子中的几个,确定了良性氧化应激作为关键。
心理生物学和氧化还原生物学之间在压力研究中的分子联系正在成为身心科学关注的焦点。一个关键的观察是在预期性皮质醇反应性中区分了好的应激和坏的应激,并且适度的压力增强了韧性。有趣的是,在关键事件压力训练中,即使在自我评估改善表现的时期,即心理上表示的“良性应激”期间,唾液皮质醇也观察到下降。 心理压力与线粒体功能之间的关系已经受到关注。已经建立了一种对情绪和照顾压力敏感的线粒体健康指数 (MHI)。MHI 整合了来自人类白细胞的信息,关于核和线粒体DNA编码参数,它反映了每单位线粒体内容量的呼吸链容量。它由琥珀酸脱氢酶/柠檬酸合成酶和细胞色素c氧化酶/线粒体DNA拷贝数组成。发现 MHI 与情绪参数相关,这些参数根据心理学压力和抑郁及焦虑症状的协议进行评估。线粒体功能障碍与抑郁症的表现密切相关,并且发现氧化应激参与了将社会孤立、孤独、努力-回报不平衡等心理社会压力与心血管疾病联系起来的过程。
五、结论
有氧生物系统中电子流动的主要方向是分解代谢、氧化的。然而,合成代谢也会发生,还原/合成途径是由从氧化还原反应获得的还原当量驱动的。对于当前上下文而言,电子流动的主要方向与持续的氧化挑战相关联,特征为氧化应激。相反,偏离氧化还原设定点的方向是向还原的方向。因此,这样的过程将被称之为‘还原性应激’,例如在缺氧或增加代谢还原条件的情况下会发生。
在这个意义上,生理学上的‘休息状态’将是一个误称,因为在那个状态下没有真的‘休息’;相反,它是连续测试和修正以朝着稳态平衡前进,即体内动态平衡。‘休息状态’可以比作汽车中的‘待机’或‘怠速’位置,准备加速而不是关闭引擎。现在三个经典的主要氧化还原响应分子开关体现了这一事实:NFkB、Nrf2/Keap1、HIF 系统通过将氧化剂和亲电体状态与它们的激活水平耦合来操作。
https://blog.sciencenet.cn/blog-41174-1417059.html
上一篇:内源性生物抗氧化系统
下一篇:水上漫步昆虫的“神奇”能力可以激发新的水生机器人
