²©ĪÄ
ĒāĘųŹĒÖ²ĪļÓ¦¼¤»ŗ½āŅņ×Ó
|
ĒāĘų»ŗ½āÖ²ĪļÓ¦¼¤µÄ×÷ÓĆ
ÕŖŅŖ ĘųĢåµŻÖŹŗĶĘųĢåŠÅŗÅ·Ö×Ó¶ŌÓŚæŲÖĘĻø°ū¹¦ÄÜ·Ē³£ÖŲŅŖ£¬ĢŲ±šŹĒŌŚÓ¦¶ŌÄę¾³Ó¦¼¤ÖŠ”£¹żČ„ŃŠ¾æÖ÷ŅŖ¼ÆÖŠŌŚŅ»Ńõ»ÆµŖ£ØŅ»Ńõ»ÆµŖ£©ŗĶĮņ»ÆĒāµČ·Ö×ÓÉĻ£¬¾”¹ÜĘäĖūµÄ·Ö×ÓČēŅŅĻ©ŗĶŅ»Ńõ»ÆĢ¼£ØŅ»Ńõ»ÆĢ¼£©ŌŚÖ²ĪļÉśĄķ¹¦ÄÜÖŠŅ²ŗÜÖŲŅŖ”£ŌŚÕāĄļ£¬Ēā·Ö×Ó±»¼ÓČėµ½ÕāøöĮģÓņÖŠ”£ĒāĘųŌų¾±»Ö¤Ć÷æÉøÄÉĘÖ²Īļ¶ŌŅ»ĻµĮŠÓ¦¼¤Ō“µÄ·“Ó¦£¬°üĄØ±©Ā¶ÓŚÖŲ½šŹō”¢ŃĪ¶Č”¢¼«¶ĖĪĀ¶ČŗĶ×ĻĶāĻß·ųÉ䔣ĻŌČ»£¬ĒāĘųŹĒŅ»ÖÖÖŲŅŖÖ²ĪļÉśĄķŠ§Ó¦ĘųĢ壬æÉÄÜÓŠÖśÓŚ“Ł½ųÖ²ĪļÉś³¤ŗĶĮøŹ³°²Č«”£ĒāĘų·½±ćŅŌø»ĒāĖ®£ØHRW£©µÄŠĪŹ½½ųŠŠĶāŌ“ŠŌøųÓč£¬µ«Ęä¹ć·ŗŹ¹ÓĆČŌ“ęŌŚŅ»Š©ĪŹĢā”£“ĖĶā£¬ĒāĘų¶ŌĻø°ū×÷ÓƵķÖ×Ó»ł“”ÉŠ²»Ź®·ÖĒ峞”£±¾ĪÄĢÖĀŪĮĖĒāĘųŌŚÖ²ĪļĮģÓņµÄÓ¦ÓƵÄø÷øö·½Ćę£¬ŅŌ¼°æÉÄÜ“ÓĘäĖūĪļÖÖÖŠĒāĘųµÄŠ§Ó¦Ń§µ½µÄ¶«Ī÷”£
Alleviation of Plant Stress by Molecular Hydrogen | SpringerLink
Hancock J T, LeBaron T W, May J, et al. Alleviation of Plant Stress by Molecular Hydrogen[M]//Gasotransmitters Signaling in Plants under Challenging Environment. Cham: Springer International Publishing, 2023: 101-120.

Ņ»”¢Ē°ŃŌ
1987Äź£¬¶ÆĪļŹµŃé±ķĆ÷ČʤŃÜÉśµÄŹęÕÅŅņ×Ó£ØEDRF£©Źµ¼ŹÉĻŹĒĘųĢåŅ»Ńõ»ÆµŖ£ØŅ»Ńõ»ÆµŖ£©”£ÕāĪŖŃŠ¾æÉśĪļĻµĶ³ÖŠµÄ·“Ó¦ŠŌµŖĪļÖÖ£ØRNS£©“ņæŖĮĖ“óĆÅ£¬Ķ¬Ź±Ņ²¹Ū²ģµ½ĮĖĘäĖūÉśĄķŠŌĘųĢåµŻÖŹ”£ÕāÖÖ·ÖĪöŅ²¼¤·¢ĮĖ¶ŌĘäĖūæÉÄܲĪÓėĻø°ūŠÅŗÅ“«µ¼µÄŠ”·Ö×Ó»īŠŌ»ÆŗĻĪļµÄŃŠ¾æ£¬°üĄØ·ĒĘųĢ¬»īŠŌŃõĪļÖŹ£ØROS£©£¬Čē¹żŃõ»ÆĒā£¬ŅŌ¼°»īŠŌĮņ»ÆŗĻĪļ£¬ČēĮņ»ÆĒā”£2019ÄźŹĒÖ²ĪļÖŠŅ»Ńõ»ÆµŖŃŠ¾æµÄ40ÖÜÄź£¬×ī½üŅ»øöŠĀµÄ²ĪÓėÕß¼ÓČėĮĖÕāøöĆūµ„£¬¼“ĒāĘų£¬ĖüæÉŅŌøıäÖ²ĪļĻø°ūµÄ»ī¶Æ£¬²¢æÉÄÜŌŚÓ¦¶ŌŃ¹Į¦·“Ó¦ÖŠ·¢»Ó×÷ÓĆ”£ÓÉÓŚÖ²ĪļŹĒ¹Ģ×ŵģ¬ĖüĆĒŠčŅŖøüø“ŌÓµÄÉś“ę²ßĀŌĄ“æĖ·žø÷ÖÖŃ¹Į¦ĢōÕ½£¬°üĄØ±©Ā¶ŌŚ×ĻĶāĻßĻĀ”¢ÖŲ½šŹō”¢¼«¶ĖĪĀ¶Č£ØøßĪĀŗĶµĶĪĀ£©”¢øÉŗµ”¢ŗéĖ®ŗĶŃĪ¶ČµČ”£
µ±Ö²Īļ“¦ÓŚ»·¾³Ń¹Į¦ĻĀŹ±£¬Ęä²ßĀŌŹĒÓÕµ¼ŠÅŗÅ×Ŗµ¼Ķ¾¾¶£¬ÕāĶس£»įµ¼ÖĀ»łŅņ±ķ“ļµÄøı䣬“Ó¶ųÓ°ĻģĻø°ūÄŚµ°°×ÖŹµÄ×é³É£¬Ź¹Ļø°ūÄܹ»ŌöĒæ»ņ²śÉśŠĀµÄ»ī¶Æ”£ÕāŠ©×÷ÓĆŹ¹Ļø°ūÄܹ»Ó¦¶Ōµ±Ē°Ń¹Į¦£¬ÉõÖĮĪ“Ą“ĢōÕ½”£Ö²ĪļÖŠÉę¼°µÄŠÅŗÅ“«µŻ°üĄØŅ»ĻµĮŠÖ²Īļ¼¤ĖŲ£¬µ«Ņ²Éę¼°Šķ¶ąĘųĢåµŻÖŹ£¬ÕāŠ©ĘųĢåµŻÖŹŌŚĘäĖūÉśĪļĢåÖŠŅ²ŗÜÖŲŅŖ”£ĘäÖŠ°üĄØŅ»Ńõ»ÆµŖ”¢Įņ»ÆĒā”¢Ņ»Ńõ»ÆĢ¼ŗĶŅŅĻ©”£Ö²ĪļµÄŃ¹Į¦¶ŌÖ²ĪļµÄÉś³¤ŗĶÉś²śĮ¦ÓŠÖŲ“óÓ°Ļģ£¬¶ųĘųĢåµŻÖŹŌŚÖ²ĪļÓ¦¶ŌŃ¹Į¦·½ĆęĘš×ÅÖŲŅŖ×÷ÓĆ”£Ķس££¬ÕāŠ©·Ö×ÓĖł½éµ¼µÄŠÅŗÅ“«µŻÖ®¼ä“ęŌŚĻą»„×÷ÓĆŗĶ/»ņŠµ÷”£ŌŚÕāĄļ£¬ĪŅĆĒĢÖĀŪĒāĘųÓėĘäĖūĘųĢåŠÅŗÅ·Ö×ÓÖ®¼äµÄĻą»„×÷ÓĆ£¬ÖŲµćĢÖĀŪĮĖĒāĘųČēŗĪ»ŗ½āÖ²ĪļµÄŃ¹Į¦”£
¶ž”¢ĒāĘųµÄÖ²Īļ×÷ÓĆ
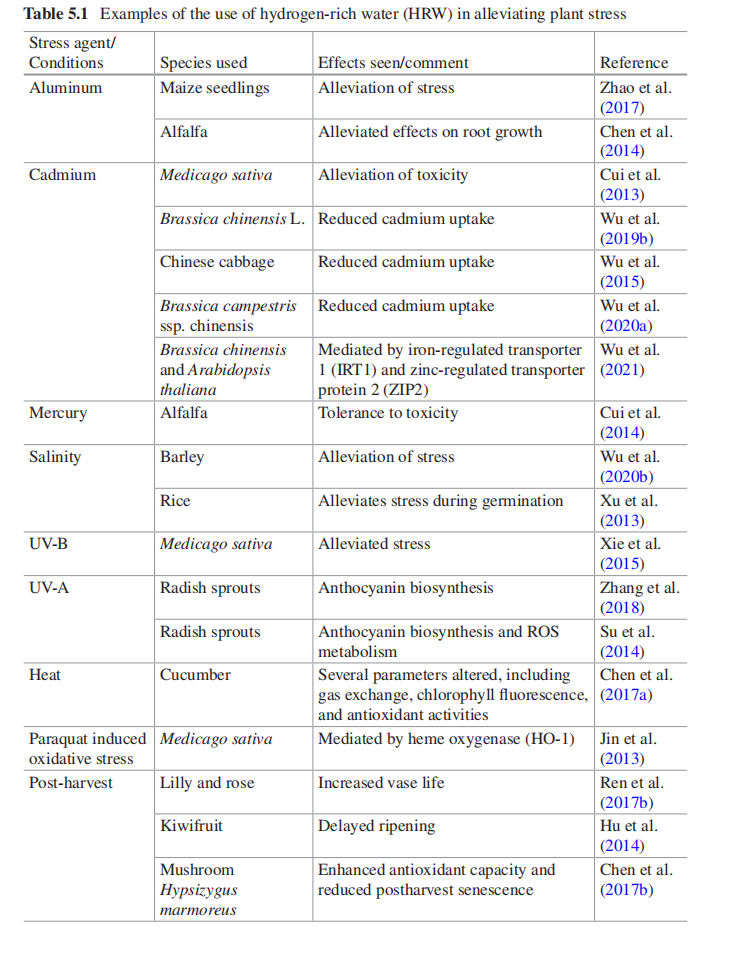
ĒāĘųŗÜÄѱ»Ö²ĪļĪüŹÕ”£²øČé¶ÆĪļæÉŅŌĪüČėĒāĘų»ģŗĻĘų»ńµĆĒāĘų£¬ĒŅÓŠŠķ¶ąŹ¹ÓĆĒāĘų³É¹¦µÄĄż×Ó£¬°üĄØÓĆÓŚÖĪĮĘCOVID-19”£Č»¶ų£¬ĒāĘų·Ē³£Ņ×Č¼£¬“ęŌŚ°²Č«ĪŹĢā£¬¶ųĒŅ±ČæÕĘųĒį£¬ĖłŅŌĒāĘų»įŃøĖŁ·ÖÉ¢µ½ÉĻ²ć“óĘųÖŠ£¬Ź¹µĆøųµŲĆęÖ²ĪļŹ¹ÓĆĘųĢå²»Ģ«ĻÖŹµ£ØŅ²²»ŹĒƻӊæÉÄÜ£¬ĪŅČĻĪŖĶعżŌŚĶĮČĄÄŚ¾¹ż¹ÜĀ·ŹäČėĒāĘų£¬ĄūÓĆĒāĘųŌŚĶĮČĄÄŚµÄĄ©É¢ŠŌÄÜ£¬ŹµĻÖøųĶĮČĄŹ©ĒāµÄ×÷ÓĆŅ²Ó¦øĆæÉŠŠ”£¹Ų¼üŹĒČēŗĪ¼õÉŁ¹¤³ĢŗĶ²ÄĮĻµÄ·ŃÓĆ”££©”£Ņņ“Ė£¬øųÖ²ĪļŹ¹ÓĆĒāĘųĶس£Éę¼°““½ØŗĶĻ”ŹĶ±„ŗĶµÄĒāĘųČÜŅŗ£¬¼“ø»ĒāĖ®£ØHRW£©”£Č»¶ų£¬ĒāĘų²»Ģ«ČÜÓŚĖ®£¬»įŃøĖŁ“ÓĖ®ÖŠŹĶ·Å³öĄ“²¢¶ŖŹ§”£ÕāæÉÄÜŠčŅŖ·“ø“ÖŲŠĀ½«HRWÓ¦ÓĆÓŚÖ²Īļ×éÖÆ£¬Ö±½ÓĶæÄØŌŚŅ¶Ćę»ņ×¢Čėøł²æ¹ąøČĖ®ÖŠ£¬ŅŌ²śÉśŠ§¹ū”£Č»¶ų£¬Čē±ķ5.1ĖłŹ¾£¬ŅŃ¾ÓŠŠķ¶ąŹ¹ÓĆHRWĄ“»ŗ½āÖ²ĪļŃ¹Į¦µÄĄż×Ó”£°üĄØ¶ŌÖŲ½šŹō”¢ĪĀ¶ČŃ¹Į¦ŗĶ¹āŃ¹Į¦µÄ·“Ó¦£¬ŗÜĆ÷ĻŌÖ²ĪļÄܹ»øŠÖŖŗĶÓ¦¶ŌĒāĘų»ņHRWµÄ“ęŌŚ”£ÓŠČ¤µÄŹĒ£¬ĒāĘųÓ¦ÓƵÄŅ»øöĒ±ŌŚÖŲŅŖÓĆĶ¾ŹĒŌŚŹÕ»ńŗóŃÓ³¤×÷ĪļµÄ“¢“ꏱ¼ä£¬ĢŲ±šŹĒĖ®¹ūŗĶ»Ø»Ü”£¶ŌHRWµÄŠĀÓ¦ÓĆŹĒŠĪ³ÉĒāÄÉĆ×ÅŻĖ®£ØHNW£©”£Õā±»ČĻĪŖæÉŅŌĢįøßĒāĘųµÄČܽā¶Č²¢ŃÓ³¤ĒāĘųµÄŹäĖĶ”£
±ķ5.1 ø»ĒāĖ®»ŗ½āÖ²ĪļÓ¦¼¤µÄŃŠ¾æ
ĪŖĮĖŹ¹ĒāĘųŌŚŹµ¼łÖŠÓŠÓĆ£¬æÉÄÜŠčŅŖæŖ·¢ŠĀµÄ”¢øüŅ×ÓŚŹ¹ÓƵÄĒāĘųŹäĖĶ·½·Ø”£ÕāŠ©·½·ØæÉÄÜĄ“×Ō²»Ķ¬µÄŠŠŅµ£¬ĄżČē£¬Ņ»ÖÖĒ±ŌŚµÄ¹©ĢåŹĒĒā»ÆĆ¾£ØMgH2£©£¬ÕāŹĒŅ»ÖÖ±»ĢįŅéÓĆÓŚĢ«ŃōÄÜĮģÓņµÄ»ÆŗĻĪļ”£ĒāĘųŹĶ·ÅµÄ¶ÆĮ¦Ń§±Č½öŹ¹ÓĆHRWøüĀżĒŅøü³Ö¾Ć£¬µ«·¢ĻÖŌŚÄūĆŹĖį»ŗ³åŅŗÖŠŹ¹ÓĆøüÓŠŠ§”£ĮķŅ»ÖÖ×ī½üÓĆÓŚÖ²ĪļÖŠŹĶ·ÅĒāĘųµÄ»ÆŗĻĪļŹĒAB@hMSN£¬Ņ»ÖÖøŗŌŲ°±ÅšĶéµÄæՊĽéæ׶žŃõ»Æ¹čÄÉĆ×æÅĮ£”£Č»¶ų£¬ÕāĄļÓŠŅ»øö¾Æøę”£Čē¹ūŹ¹ÓĆ¹©Ģå·Ö×Ó£¬ĖüĆĒæÉÄÜ»įĮōĻĀø±²śĘ·£¬ÕāæÉÄÜ»įŃĻÖŲĪ£¼°ĒāĘųµÄ°²Č«ÉśĪļŹ¹ÓĆ”£
Čż”¢ĒāĘųµÄŠ§Ó¦·Ö×Ó”£
ĒāĘų·Ö×Ó·Ē³£Š”£ØĻą¶ŌÓŚĘäĖūŠÅŗÅ·Ö×Ó£©£¬²¢ĒŅĻą¶Ō¶čŠŌ”£Ņņ“Ė£¬ŗÜÄŃĻėĻóĖüŹĒČēŗĪ±»Ļø°ūøŠÖŖŗĶ×÷ÓƵĔ£ĄżČē£¬Ź¹ÓĆ»ÆѧĒ÷»ÆŅņ×ӵľµä¼¤ĖŲŠĶŠÅŗÅ“«µ¼»įŹ¹ÓƵ°°×ÖŹŹÜĢ壬µ«Õā²»Ģ«æÉÄÜŹŹÓĆÓŚĒāĘųÕāŃłµÄ·Ö×Ó”£Ņ»Š©ŠÅŗÅ·Ö×Ó£¬ČēŅ»Ńõ»ÆµŖ£¬»įĶعż¼ŁĢå»łĶÅ»ņÓėŪĻ»ł·“Ó¦¶ųÓėµ°°×ÖŹ·“Ó¦”£Č»¶ų£¬Ķ¬ŃłµŲ£¬ŗÜÄŃ擳öÕāÖÖĄąŠĶµÄ·“Ó¦ČēŗĪŹŹÓĆÓŚĒāĘų£¬ŅņĪŖÓėŅ»Ńõ»ÆµŖ²»Ķ¬£¬ĒāĘųŹĒĪŽ¼«ŠŌµÄ£¬²¢ĒŅ²»¾ßÓŠ·“Ó¦ŠŌ”£Ņņ“Ė£¬±ŲŠė“ęŌŚĘäĖū»śÖĘĄ“½āŹĶĒāĘųøųŅ©ŗó¹Ū²ģµ½µÄÉśĪļŠ§Ó¦”£
¹ŲÓŚĒāĘų×÷ÓƵÄÖ÷ŅŖĀŪµćÖ®Ņ»ŹĒĖüÓ°ĻģĻø°ūÖŠµÄæ¹Ńõ»Æ¼ĮĖ®Ę½”£Šķ¶ąÕāŠ©Š§Ó¦ŹĒ¼ä½ÓµÄ£¬Éę¼°æ¹Ńõ»ÆÄÜĮ¦Ļø°ūÖŠĆøµÄ±ķ“ļ»ņ»żĄŪ·¢Éś±ä»Æ”£Č»¶ų£¬ÕāÖ»ÓŠŌŚĒāĘų·Ö×Ó±»Ö±½ÓøŠÖŖµÄĒéæöĻĀ²Å»į·¢Éś£¬¶ųÕāĶس£ŹĒĪÄĻ×ÖŠ»Ų±ÜµÄĪŹĢā”£
¾Ż±ØµĄ£¬ĒāĘųČ·ŹµæÉŅŌÖ±½Ó×÷ĪŖæ¹Ńõ»Æ¼ĮĘš×÷ÓĆ£¬ĶعżÓėōĒ×ŌÓÉ»ł£Ø⋅OH£©·“Ó¦£¬µ«²»ÓėĘäĖūROS£ØČē³¬ŃõŅõĄė×Ó£ØO2⋅−£©»ņ¹żŃõ»ÆĒā£©·“Ó¦”£ŅŃÖŖ⋅OH²ĪÓėÖ²ĪļÓ¦¼¤·“Ó¦£¬ČēÖŲ½šŹōŠ²ĘČ”¢°Ł²ŻæŻ“¦ĄķŅŌ¼°ŗ®ŗ¦ŗĶøÉŗµŠ²ĘȵȔ£Ņņ“Ė£¬Ķعż×ŌÓÉ»łĒå³ż¼ĮČ„³ż⋅OH£ØÕāĄļÖøĒāĘų£©æÉŅŌ½āŹĶŅ»Š©¹Ū²ģµ½µÄŠ§¹ū”£¾”¹ÜČē“Ė£¬ŗ󥓵ÄŅ»ĘŖĀŪĪÄĢį³ö¶ŌÕāÖÖ·“Ó¦¶ÆĮ¦Ń§µÄĆÜĒŠµ÷²é²¢²»Ö§³ÖÕāŅ»¹Ūµć£¬Źµ¼ŹÉĻ£¬ÓŠČĖĢį³ö⋅OH»įŌŚĒāĘųÖ®Ē°ÓėĘäĖūÉśĪļ·Ö×Ó·“Ó¦£¬Ņņ“Ė⋅OHµÄŠ§¹ū²»»įŅņĒāĘųµÄ¼ÓČė¶ų¼õČõ”£ĄąĖʵŲ£¬µŚ¶žøöÖ±½ÓÄæ±ź±»ČĻĪŖŹĒ¹żŃĒĻõĖįøłĄė×Ó£ØONOO−£©”£ÕāŹĒÓɳ¬ŃõŅõĄė×Ó£ØO2⋅−£©ÓėŅ»Ńõ»ÆµŖ·“Ó¦²śÉśµÄ£¬ÓÉÓŚĮ½ÕßŌŚÓ¦¼¤·“Ó¦ÖŠŹ±¼äŗĶæÕ¼äÉĻĶ¬Ź±²śÉś£¬Ņņ“ĖŗÜæÉÄÜ“ęŌŚONOO−”£Čē¹ūĒāĘųČ„³żONOO−£¬ÕāæÉŅŌ½āŹĶ¹Ū²ģµ½µÄŠ§¹ū”£Č»¶ų£¬½ųŅ»²½×ŠĻø¼ģ²é·Ö×Ó¶ÆĮ¦Ń§ĖĘŗõÅųżĮĖONOO−×÷ĪŖĒāĘųµÄÖ±½ÓÄæ±ź”£
ÓÉÓŚ⋅OHŗĶONOO−¶¼Äܱ»ÅųżŌŚĶā£¬ÓŠČĖ×ī½üĢį³öæÉÄܵÄÄæ±źŹĒČż¼ŪĢś£ØFe3+£©Ąė×Ó(Penders et al. 2014)”£ÕāÓėĘäĖūĘųĢåŠÅŗÅ“«µŻÕߵıØøę²¢ĪŽ²»Ņ»ÖĀÖ®“¦”£Ņ»Ńõ»ÆµŖµÄŅ»øöÖ÷ŅŖ×÷ÓĆŹĒĶعżŅ»Ńõ»ÆµŖÓėĆøµÄŃŖŗģĖŲøØ»łÖ±½ÓĻą»„×÷ÓĆ¼¤»īæÉČÜŠŌÄńÜÕĖį»·»ÆĆø£ØsGC£©”£ŌŚÕā·½Ćę£¬ŃŠ¾æĮĖĒāĘųŌŚ¼”ŗģµ°°×”¢Ļø°ūÉ«ĖŲP450ŗĶ¼Łµ„°ūŃõ»¹µ°°×ÖŠ¶ŌĢśµÄ»¹Ō×÷ÓĆ£¬µ«µĆ³ö½įĀŪČĻĪŖŌŚÕāŠ©µ°°×֏֊ƻӊŃŖŗģĖŲ»ņĢśĮņ£ØFe/S£©“ŲµÄ»¹Ō”£Č»¶ų£¬¾ßÓŠ-414 mVµÄŃõ»Æ»¹ŌµēĪ»ÖŠµć£¬ĒāĘųæÉŅŌŌŚø÷ÖÖĆøÖŠČČĮ¦Ń§ÉĻ»¹ŌŅ»ĻµĮŠŃŖŗģĖŲ»łĶÅ£¬²¢½ØŅ齫Ęä×÷ĪŖĪ“Ą“ŃŠ¾æµÄ½¹µć”£ÕżČēĖłĢÖĀŪµÄ£¬NAPDHŃõ»ÆĆøĶ¬ĻµĪļµČĆøÓČĘäæÉÄܲĪÓėø÷ÖÖÓ¦¼¤·“Ó¦”£²»ÄÜÅųżsGCŅ²æÉÄÜŹĒĒāĘųµÄÄæ±ź”£ĻŌČ»£¬ŌŚÕā·½ĆęŠčŅŖ×öøü¶ąµÄ¹¤×÷£¬Ź¹ÓĆŗ¬ÓŠŃŖŗģĖŲ»ņFe/SøØ»łµÄø÷ÖÖÖ²Īļµ°°×ÖŹ£¬Č»ŗóŌŁÅųżÕāÖÖ»śÖʵÄæÉÄÜŠŌ”£Č»¶ų£¬æÉÄÜŠčŅŖ¶Ō“Ė±£³ÖŅ»¶ØµÄ½÷É÷£¬ŅņĪŖĖü²»ÄÜ×ÜŹĒ¼ŁÉčŌŚ¶ÆĪļĻµĶ³ÖŠČ·¶ØµÄŠÅŗÅĶØĀ· ŌŚÖ²ĪļÖŠŅ²ŹĒĻąĶ¬µÄ”£ĄżČē£¬Ņ»Ńõ»ÆµŖ¶ŌÖ²ĪļsCGµÄ×÷ÓĆŅŃ¾ŹÜµ½ÖŹŅÉ”£Ņņ“Ė£¬ĒāĘųµÄ×÷ÓĆŅ²æÉÄܲ»Ķ¬£¬¾”¹ÜŹ¹ÓĆĘäĖūÉśĪļĻµĶ³Ą“ĶĘ½ųÖ²ĪļæĘѧŹĒŅ»ÖÖĒæ“óµÄ¹¤¾ß£ØĻĀŹö£©”£
¾Ż±ØµĄ£¬ĒāĘųµÄŅ»Š©Š§Ó¦ŹĒÓÉŃŖŗģĖŲ¼ÓŃõĆø£ØHO-1£©½éµ¼µÄ”£øĆĆø“ß»ÆŃŖŗģĖŲ·Ö½āµÄ·“Ó¦£Ø1£©Éę¼°ŃõĘų£»£Ø2£©Ź¹ÓĆNADPH×÷ĪŖøØŅņ×Ó£»£Ø3£©²śÉśµØĀĢĖŲ”¢Ņ»Ńõ»ÆĢ¼ŗĶĢśĄė×Ó”£Č»¶ų£¬ÓėĒāĘųµÄ¾ßĢå·“Ó¦Ļø½ŚÉŠĪ“±ØµĄ£¬Ņņ“ĖŅ²æÉÄÜÖ»ŹĒĒāĘųŠ§Ó¦ĻĀÓĪŠÅŗÅ“«µ¼µÄ½į¹ū£¬ÕāÉŠ“żČ·¶Ø”£ĮķŅ»ÖÖ¾ŻŠÅ½éµ¼ĒāĘųŠ§Ó¦µÄĆøŹĒ¹Čė×øŹėĹżŃõ»ÆĪļĆø£¬ÕāŹĒŅ»ÖÖĪ¬³ÖĻø°ūÄŚŃõ»Æ»¹ŌĘ½ŗāµÄÖŲŅŖĆø”£ĶعżŹ¹ÓĆ»łŅņȱĻŻ¾śÖźŗĶŅÖÖĘ¼ĮµÄŃŠ¾æĻŌŹ¾£¬ŌŚĮéÖ„Õę¾śÖŠŠčŅŖ¹Čė×øŹėĹżŃõ»ÆĪļĆøĄ“½éµ¼ĒāĘųµÄ×÷ÓĆ”£ÕāÖÖ¹Čė×øŹėĹżŃõ»ÆĪļĆøŹĒŅ»ÖÖŗ¬Īųµ°°×£¬Ź¹Ęä³ÉĪŖŅ»ÖÖӊȤµÄĒ±ŌŚĒāĘų°Š±ź£¬µ«Ņ²²»ÄÜÅųżĒāĘųµÄÖ±½Ó×÷ÓĆĪ»ÓŚĆøµÄÉĻÓĪŠÅŗÅ”£
×īŗó£¬ÓŠČĖĢį³ö£¬ÓÉÓŚĒāĘųÓŠĮ½ÖÖ×ŌŠż×“Ģ¬£¬ÕāæÉÄÜŹĒĒāĘųÓ°ĻģĘäĖūÉśĪļ·Ö×ÓµÄŅ»ÖÖ·½Ź½”£Č»¶ų£¬µ½ÄæĒ°ĪŖÖ¹£¬»¹Ć»ÓŠÕā·½ĆęµÄŹµŃéÖ¤¾Ż”£
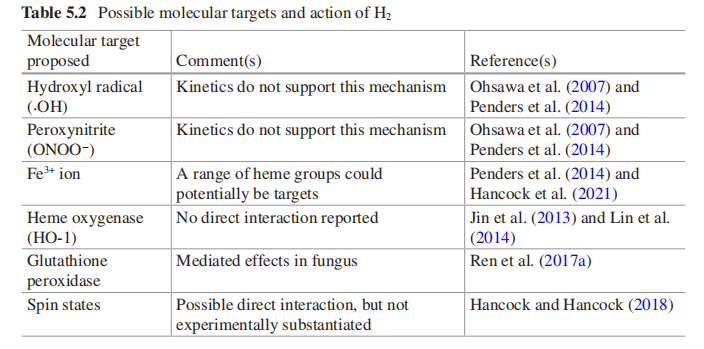
ĖäČ»ŅŃ¾Ģį³öĮĖ¶ąÖÖ»śÖĘ£¬µ«ÄæĒ°»¹Ć»ÓŠČ·¶ØĒāĘų¾ßĢåČēŗĪÖ±½ÓÓėÉśĪļĻµĶ³Ļą»„×÷ÓƵÄĆ÷Č·»śÖĘ£Ø±ķ5.2£©”£Ņņ“Ė£¬ŌŚĒāĘųŠ§Ó¦»śÖĘ·½ĆęČŌČ»ŠčŅŖ×öøü¶ąµÄ¹¤×÷”£¾”¹ÜČē“Ė£¬ÓÉÓŚĒāĘų¶ŌÖ²ĪļµÄ×÷ÓĆ·Ē³£Ć÷Č·£Ø±ķ5.1£©£¬ŌŚ²»ĮĖ½ā·Ö×Ó»śÖʵÄĒéæöĻĀČŌČ»æÉŅŌĄūÓĆÕāŅ»ĻÖĻó£¬ĢŲ±šŹĒĖĘŗõƻӊ±Øøę±ķĆ÷ĒāĘųÓ¦ÓƶŌÖ²ĪļŗĶ¶ÆĪļÓŠĆ÷ĻŌŗ¦“¦”£ĒāĘų»ÆѧĢŲÕ÷¾ö¶ØĮĖĖĘŗõ²»»įĮōĻĀø±²śĘ·£¬Ņņ“Ė¶ŌŹ³Ę·°²Č«²»»į²śÉśÓ°Ļģ”£
±ķ5.2 ĒāĘųµÄæÉÄÜ·Ö×Ó°ŠµćŗĶ×÷ÓĆ
ĖÄ”¢ĒāĘųµÄĻø°ūŠÅŗÅ×÷ÓĆ”£
ÓėĖłÓŠĪļÖÖŅ»Ńł£¬Ö²ĪļÖŠĻø°ūŠÅŗÅ“«µ¼¶ŌÓŚÉśĪļĢå×Ā׳³É³¤ŗĶÓ¦¶ŌŃ¹Į¦ĢōÕ½ÖĮ¹ŲÖŲŅŖ”£øŠÖŖĶā²æŠÅŗÅ£ØÉśĪļ»ņ·ĒÉśĪļ£©ŅŌ¼°ŠÅŗÅ“«µ¼Ķ¾¾¶£¬µ¼ÖĀ·“Ó¦µÄ¹ż³ĢÉę¼°Ņ»ĻµĮŠµ°°×ÖŹŗĶŠ”·Ö×Ó”£ŌŚÕāŠ©Ķ¾¾¶ÖŠĘš¹Ų¼ü×÷ÓƵďĒĻą¶Ō»īŌ¾µÄŠ”ĘųĢåŠÅŗÅ·Ö×Ó£¬ČēŅ»Ńõ»ÆµŖŗĶĮņ»ÆĒā”£¾”¹ÜČēĒ°ĖłŹö£¬ŗÜÄŃĻėĻóĒāĘųŅ²ÄÜÖ±½Ó²ĪÓėÉś»Æµ÷½Ś×÷ÓĆ²¢¶Ō¶ąėIJśÉśÓ°Ļģ£¬µ«ŹĒÓŠ“óĮæÖ¤¾Ż±ķĆ÷ĒāĘųÄÜÓėĘäĖūĘųĢåŠÅŗÅ·Ö×ÓŗĶŠ”Ńõ»Æ»¹Ō»ÆŗĻĪļµÄĻą»„×÷ÓĆ»ņÓ°Ļģ”£ŅŌĻĀĢÖĀŪŅ»Š©Õā·½ĆęµÄÖ¤¾Ż”£
4.1 ĒāĘų¶ŌŅ»Ńõ»ÆµŖµÄÓ°Ļģ
¼øŹ®ÄźĄ“ŅŃÖŖ£¬Ö²Īļ²śÉśŅ»Ńõ»ÆµŖ²¢¶ŌĘ书ÄܲśÉśÉīŌ¶Ó°Ļģ”£ŗĮĪŽŅÉĪŹ£¬Ņ»Ńõ»ÆµŖŌŚæŲÖĘĻø°ū¹¦ÄÜ·½ĆęĘš×ÅŗĖŠÄ×÷ÓĆ£¬¶ų×ī½ü·¢ĻÖĒāĘųŌŚŅ»Ńõ»ÆµŖĶ¾¾¶ÖŠĻą»„×÷ÓĆ”£
µ±HRWÓĆÓŚ»ŗ½āÜŁŽ£µÄĀĮŠ²ĘČŹ±£¬¾Ż±ØµĄŅ»Ńõ»ÆµŖÉś³É¼õÉŁ”£50%±„ŗĶHRW½µµĶĮĖŅ»Ńõ»ÆµŖ¹©ĢåµÄŠ§Ó¦£¬±ķĆ÷Ņ»Ńõ»ÆµŖæÉÄܽ鵼ĮĖĒāĘųŠ§Ó¦”£Ļą·“£¬µ±ŃŠ¾æøłÉś³¤Ź±£¬ĒāĘųŌö¼ÓĮĖ·¬ĒŃÓ×ĆēÖŠµÄŅ»Ńõ»ÆµŖ²śÉś”£Õā±»Ņ»Ńõ»ÆµŖĒå³ż¼Į2-4-ōČ»ł±½»ł-4,4,5,5-Ėļ׻łßäßņßų-1-Ńõ»ł-3-Ńõ»ÆĪļ£ØcPTIO£©Ėł½µµĶ£¬Õā±ķĆ÷ĒāĘų²¢Ī“Ö±½ÓĒå³żŅ»Ńõ»ÆµŖ”£ŃŠ¾æ½įĀŪŹĒ£¬Éś³¤ĖŲÓÕµ¼µÄĒāĘųÉś³ÉĖęŗóÓÉĻõĖį»¹ŌĆø²śÉśµÄŅ»Ńõ»ÆµŖ½éµ¼”£ŌŚ»Ę¹ĻÖŠŅ²±ØµĄĮĖĄąĖĘ½į¹ū£¬ĘäÖŠHRWŌö¼ÓĮĖøłÉś³¤ŗĶŅ»Ńõ»ÆµŖ»żĄŪ”£HRWŗĶŅ»Ńõ»ÆµŖ¶¼Ōö¼ÓĮĖĻø°ūÖÜĘŚ»łŅņµÄ±ķ“ļ£¬°üĄØCycA£ØAŠĶÖÜĘŚµ°°×£©”¢CycB£ØBŠĶÖÜĘŚµ°°×£©”¢CDKA£ØÖÜĘŚµ°°×ŅĄĄµŠŌ¼¤ĆøA£©ŗĶCDKB£ØÖÜĘŚµ°°×ŅĄĄµŠŌ¼¤ĆøB£©”£ÕāŠ©Š§Ó¦±»ĄąĖĘĻõĖį»¹ŌĆøŗĶŅ»Ńõ»ÆµŖŗĻ³ÉĆøŅÖÖĘ¼ĮŅŌ¼°Ņ»Ńõ»ÆµŖĒå³ż¼ĮĖł¼õČõ”£Ņ»Ńõ»ÆµŖ»¹½éµ¼ĮĖ»Ę¹ĻÖŠÓÉĒāĘųÓÕµ¼µÄøłÉś³¤£¬ĘäÖŠĻĀÓĪµ°°×ÖŹ±»¼ų¶ØĪŖ֏ĤH+-ATPaseŗĶ14-3-3µ°°×”£ŗóÕߏĒÖīČēĖæĮŃŌ»ī»Æµ°°×¼¤Ćø£ØMAPK£©ŗĶp53µČ°ūÄŚŠÅŗÅ“«µ¼¼¶ĮŖµÄ¹Ų¼üµ÷½Śµ°°×”£µ±øłŠĪ³ÉÓÉĒāĘųŹĶ·Å¹©ĢåAB@hMSNÓÕµ¼Ź±£¬»¹·¢ĻÖĆøĻõĖį»¹ŌĆø²ĪÓėĮĖŅ»Ńõ»ÆµŖµÄ²śÉś”£
ĒāĘų¾ßÓŠŌŚÖ²Īļ²ÄĮĻ²Éŗó“¢“ę·½ĆęµÄĒ±Į¦”£1% HRW [2.2 ¦ĢM H2]£Øøł¾Ż×÷ÕߵĊÅĻ¢¼ĘĖć£©ŗĶĻõĘÕÄĘ£ØSNP£ŗ150 ¦ĢM£©øÄÉĘĮĖĒŠ»Ø°ŁŗĻµÄĘæ²åŹŁĆü£¬µ±Č„³żŅ»Ńõ»ÆµŖŹ±ÕāŠ©Š§Ó¦¼õČõ”£ŌŚ¶Ō±ķ“ļµÄ»łŅņ½ųŠŠŃŠ¾æŹ±»¹·¢ĻÖŅ¶ĀĢĢåATPŗĻĆøCF1¦ĮŃĒ»ł£ØAtpA£©æÉÄÜŌŚ½éµ¼ÕāŠ©Š§Ó¦·½ĆęŗÜÖŲŅŖ”£“ĖĶā£¬Ī÷ŗģŹĮÖŠµÄĻõĖįŃĪ»żĄŪĶعżH2¼õÉŁ£¬ÕāæÉÄܶŌĖ®¹ūµÄ“¢“ę·½Ź½²śÉśÓ°Ļģ”£
Ņņ“Ė£¬ĻŌČ»ĒāĘų¶ŌŅ»Ńõ»ÆµŖ“śŠ»ÓŠÓ°Ļģ£¬²¢ÓÉĘä½éµ¼£¬ÕāĖĘŗõ²»ŹĒÓÉÓŚĒāĘųÖ±½ÓĒå³żŅ»Ńõ»ÆµŖĖłÖĀ£¬ÕāÓėÖ®Ē°µÄ±ØµĄŅ»ÖĀ”£Č»¶ų£¬ĒāĘųŅ²æÉÄÜĶعżROS²śÉśÓ°Ļģ£¬ÕāŅ²æÉÄÜ»įÓ°ĻģŅ»Ńõ»ÆµŖ“śŠ»”£ĄżČē£¬ĶŃĀäĖį£ØABA£©ÓÕµ¼ÄāÄĻ½ęÖŠĒāĘųµÄ»żĄŪ£¬“Ó¶ųŌöĒæĮĖøÉŗµÄĶŹÜŠŌ”£Č»¶ų£¬ÕāŠ©Š§Ó¦Ņ²Éę¼°ROSŗĶŅ»Ńõ»ÆµŖ»żĄŪ£¬Ź¹ÓĆĮĖĆøĻõĖį»¹ŌĆøŗĶNADPHŃõ»ÆĆø”£ŹĀŹµÉĻ£¬·¢ĻÖĒāĘų“Ł½ųŅ»Ńõ»ÆµŖ»żĄŪŅĄĄµÓŚROSµÄ²śÉś£¬±ķĆ÷ĒāĘų²ĪÓėĮĖŅ»øöø“ŌÓĒŅĻą»„ŅĄ“ęµÄĻµĶ³”£
4.2 ĒāĘų¶Ō»īŠŌŃõŗĶæ¹Ńõ»ÆĆøµÄÓ°Ļģ
ŗÜĆ÷ĻŌ£¬ŌŚæ¼ĀĒĒāĘųŠ§Ó¦Ź±ŠčŅŖæ¼ĀĒROS“śŠ»£¬ĢŲ±šŹĒŅņĪŖĒāĘųæÉÄܾßÓŠæ¹Ńõ»Æ¼ĮŗĶ“ŁŃõ»Æ¼ĮµÄ×÷ÓĆ”£
¾”¹ÜÖ±½ÓĒå³ż³¬ŃõŅõĄė×ÓO2⋅−ŗĶ¹żŃõ»ÆĒā±»ÅųżŌŚĶā£¬¹żŃõ»ÆĒāµÄĒå³żŅ²ŹÜµ½ÖŹŅÉ£¬µ«Šķ¶ą±Øøę±ķĆ÷ĒāĘųĶعżµ÷½ŚĻø°ūµÄæ¹Ńõ»ÆÄÜĮ¦¶ŌÖ²Īļ²śÉśÓ°Ļģ”£ÓĆĒāĘų“¦Ąķ¾Ā²Ėŗó¼õÉŁĮĖŃõ»ÆĖšÉĖ²¢Ōö¼ÓĮĖ¼øÖÖæ¹Ńõ»ÆĆøµÄ»īŠŌ£¬°üĄØ³¬Ńõ»ÆĪļĘē»ÆĆø£ØSOD£©”¢¹żŃõ»ÆĒāĆø£ØCAT£©”¢Óś““ľ·Ó¹żŃõ»ÆĪļĆø£ØPOD£©ŗĶæ¹»µŃŖĖį¹żŃõ»ÆĪļĆø£ØAPX£©£¬“Ó¶ųŃÓ³¤ĮĖ¾Ā²ĖµÄ±£ÖŹĘŚ”£UV-BÕÕÉäŗó“óĀóÖŠµÄŃõ»ÆÓ¦¼¤Ņ²Ķعżøıäæ¹Ńõ»Æ¼ĮµĆµ½»ŗ½ā£¬ÓČĘäŹĒĄą»ĘĶŖ”£HRW»¹Ź¹ÓńĆ×øüÄÜÄĶŹÜ¹āŠ²ĘČ£¬Ķ¬ŃłŅ²ŹĒĶعżæ¹Ńõ»ÆĆøŹµĻֵģØÕŵČČĖ£¬2015Äź£©”£ĘäÖŠ°üĄØSOD”¢CATŗĶAPX£¬ĖüĆĒ¼õÉŁĮĖO2⋅−ŗĶ¹żŃõ»ÆĒāµÄ»żĄŪ”£
ĶعżĻņÖ²ĪļŹ©ÓĆĒāĘųĄ“Ó¦¶ŌĘäĖūŃ¹Į¦ĢōÕ½Ėł“ųĄ“µÄÄĶŹÜŠŌŅ²ŹĒÓÉæ¹Ńõ»Æ¼Į½éµ¼µÄ”£Õā°üĄØÓńĆ׶ŌĀĮµÄÄĶŹÜŠŌ£¬ĘäÖŠHRWøıäĮĖCAT”¢APX”¢SODŗĶPODµÄĻø°ūĖ®Ę½”£ŌŚ±©Ā¶ÓŚŗ®ĄäÓ¦¼¤µÄĖ®µ¾Ó×ĆēÖŠ£¬SODĖ®Ę½·¢Éś±ä»Æ£¬ÕāĖĘŗõŹĒÓÉmiRNAĖ®Ę½µÄ±ä»Æ½éµ¼µÄ£¬ĢŲ±šŹĒmiR-398×ŖĀ¼±¾”£×÷ÕßČĻĪŖÕā¶ŌÓŚĪ¬³ÖĻø°ūÄŚµÄŃõ»Æ»¹ŌĘ½ŗāÖĮ¹ŲÖŲŅŖ”£
Ļø°ūÄŚæ¹Ńõ»Æ»īŠŌµÄ±ä»Æ²»½öæÉŅŌ¼õĒįŃõ»ÆÓ¦¼¤µÄÓŠŠĪ·½Ćę£¬Čē¼õÉŁÖ¬ÖŹ¹żŃõ»ÆŗĶµ°°×ÖŹŃõ»Æ£¬¶ųĒŅ»¹½«³ÉĪŖĪ¬³ÖĻø°ūŃõ»Æ»¹ŌĘ½ŗāĻµĶ³µÄŅ»²æ·Ö£¬Õā½«ŹĒŠÅŗÅ“«µ¼ÖŠø“ŌÓĻą»„×÷ÓƵÄŅ»²æ·Ö”£½µµĶROSĖ®Ę½½«ŅāĪ¶×ÅÓėŅ»Ńõ»ÆµŖµÄ·“Ó¦æÉÄÜ»į¼õÉŁ£¬“Ó¶ų¼õÉŁONOO−µÄ²śÉś£¬ŗóÕߏĒŅ»Ńõ»ÆµŖµÄĻĀÓĪŠÅŗÅ·Ö×Ó”£ÖŚĖłÖÜÖŖ£¬ROS»¹»į×÷ÓĆÓŚ¹Čė×øŹėÄ£¬ÕāŹĒŅ»ÖÖÓŠÖśÓŚĪ¬³ÖĻø°ūŃõ»Æ»¹ŌĘ½ŗāµÄ·Ö×Ó”£ÓŠČĖČĻĪŖĻø°ūµÄŃõ»Æ»¹ŌדĢ¬±»¾«ŠÄ±£³ÖŌŚŅ»øö”°½š·¢¹ĆÄļĒų”±£¬Ņņ“ĖČĪŗĪĻø°ūÄŚŃõ»Æ»¹Ō·Ö×ӵıä»Æ¶¼»įÓ°ĻģÕāøöĒųÓņ”£Ņ»øöŗÜŗƵĥż×ÓŹĒ£¬ÕāÖÖŃõ»Æ»¹Ō»īŠŌ·Ö×ÓČēŗĪĻą»„×÷ÓĆŅŌŌŚÖ²ĪļÖŠ²śÉśŠ§¹ū£¬ÕāæÉŅŌ“Ó¶¹æĘÖ²ĪļÓėøłĮö¾ś¹²ÉśĻµĶ³ÖŠæ“µ½£¬ĘäÖŠŃŠ¾æĮĖGSH”¢Ņ»Ńõ»ÆµŖŗĶROS”£
4.3 ĒāĘųÓėŅŅĻ©ŠÅŗÅ“«µ¼
ŅŅĻ©£ØC2H4£»H2C=CH2£©ŹĒ×īÖŖĆūµÄĘųĢåŠÅŗÅ·Ö×ÓÖ®Ņ»”£Ėü²ĪÓėÖ²ĪļµÄø÷ÖÖÉśĄķĻµĶ³£¬ČēÖ²ĪļÉś³¤£¬µ«æÉÄÜ×īÖŖĆūµÄŹĒĖüŌŚ¹ūŹµ³ÉŹģÖŠµÄ×÷ÓĆ”£
ŅŅĻ©ÓėĘäĖūĘųĢåŠÅŗÅ·Ö×ÓµÄĻą»„×÷ÓĆ²¢²»ŠĀĻŹ”£ĄżČē£¬ŅŃ¾±ØµĄŅ»Ńõ»ÆµŖŗĶŅŅĻ©ŌŚ»Ę¹ĻµÄøłĻµ·¢ÓżÖŠ¹²Ķ¬×÷ÓĆ”£Ņņ“Ė£¬ĒāĘųÓėŅŅĻ©µÄĻą»„×÷ÓĆ²¢²»ĮīČĖ¾ŖŃČ”£ĶعżĒāĘų“¦ĄķæÉŅŌ½µµĶƵ¹å»Ø¶äµÄ²ÉŗóĖ„ĄĻ£¬ÕāŹĒĶعżøıäŅŅĻ©ŠÅŗÅ“«µ¼Ą“ŹµĻֵĔ£µ×ĪļŗĶÉśĪļŗĻ³ÉĆø¼õÉŁĮĖ£ŗ1-°±»ł»·±ūĶé-1-ōČĖįõ„£ØACC£©£»ACCŗĻĆø£ØACS£©£»ŗĶACCŃõ»ÆĆø£ØAŅ»Ńõ»ÆĢ¼£©”£±ąĀėÉśĪļŗĻ³ÉĆøµÄRh-ACS3ŗĶRh-AŅ»Ńõ»ÆĢ¼1×ŖĀ¼±¾µÄ»łŅņ±ķ“ļŅ²¼õÉŁĮĖ”£ÓŠČ¤µÄŹĒ£¬ŅŅĻ©ŹÜĢåRh-ETR1µÄ±ķ“ļŌö¼ÓĮĖ”£ÕāŠ©Źż¾ŻĒ峞µŲ±ķĆ÷ĒāĘų¶ŌŅŅĻ©“śŠ»ŗĶ°éĖęµÄŠÅŗÅ“«µ¼ÓŠÓ°Ļģ”£
Ņ»Ļīµ°°×ÖŹ×éŃŠ¾æ»¹±ķĆ÷£¬ĒāĘųŗĶŅŅĻ©ŌŚŠÅŗÅ“«µ¼ÖŠŗĻ×÷”£Ź¹ÓĆ»Ę¹Ļøł×÷ĪŖÄ£ŠĶĻµĶ³£¬ŃŠ¾æ±ķĆ÷ŅŅĻ©ŠÅŗÅ“«µ¼µÄŅÖÖĘ¼ĮAgNO3ŗĶaminoethoxyvinylglycine£ØAVG£©½µµĶĮĖĒāĘų“¦ĄķÓÕµ¼µÄ²»¶Øøł·¢Óż”£µ°°×ÖŹ×é·ÖĪöŹ¹ÓƶžĪ¬Äż½ŗµēÓ¾½įŗĻÖŹĘ×·ÖĪöĻŌŹ¾£¬HRWÓÕµ¼ĮĖ¾ÅÖÖµ°°×ÖŹµÄÉĻµ÷ŗĶŹ®ĪåÖÖµ°°×ÖŹµÄĻĀµ÷”£×÷ÕßµĆ³ö½įĀŪ£¬ŅŅĻ©ŹĒĒāĘųµÄĻĀÓĪ£¬²¢ĒŅÓŠĮłÖÖµ°°×ÖŹÖµµĆ¹Ų×¢£¬ĖüĆĒ½éµ¼ĮĖĒāĘųŠ§Ó¦”£ÕāŠ©µ°°×ÖŹŹĒRuBisco”¢ŃõŗĻŌöĒæµ°°×£ØOEE1£©”¢ĘßĮ×ĖįĮņŠĮĖįĶŃĒāĆø£ØSBPase£©”¢ĖÕ°±ĖįĶŃĒāĆø£ØTDH£©”¢°ūÖŹæ¹»µŃŖĖį¹żŃõ»ÆĪļĆø£ØcAPX£©ŗĶµ°°×ÖŹ¶žĮņ¼üŅģ¹¹Ćø£ØPDI£©”£
4.4 ĒāĘųÓėĮņ»ÆĒāŠÅŗÅ“«µ¼
Įņ»ÆĒā±»ČĻĪŖŹĒÓŠ¶¾µÄ£¬µ«ĻÖŌŚŅ²±»½ÓŹÜĪŖæŲÖĘÉśĄķѧŗĶĻø°ū¹¦ÄܹŲ¼üŹĀ¼žµÄÖĪĮĘŠŌĘųĢåŠÅŗÅ·Ö×Ó”£Č»¶ų£¬ÓėĘäĖūŠ”·“Ó¦ŠŌ»ÆŗĻĪļŅ»Ńł£¬Įņ»ÆĒā²¢²»ŹĒµ„¶Ą×÷ÓĆ£¬¶ųŹĒÕāŠ©·Ö×Ó²ĪÓėµÄø“ŌÓĻą»„×÷ÓƵÄŅ»²æ·Ö”£ÓŠČĖĢį³öĮņ»ÆĒāæÉÄܶŌĘäĖūŅ»Š©ŠÅŗÅĶØĀ·Ęšµ½ŅÖÖĘ×÷ÓĆ”£“ĖĶā£¬»¹ÓŠ±ØµĄ³ĘĮņ»ÆĒāŅ²ŹĒĻø°ūÄŚĒāĘųŠÅŗÅ“«µ¼µÄŅ»²æ·Ö”£
ĶعżŌŚÄāÄĻ½ęÖŠ±ķ“ļĄ“×ŌĄ³ŅšŅĀŌå£ØCrHYD1£©µÄĒāĘųĆø»łŅņ£¬µ¼ÖĀĒāĘųÉśĪļŗĻ³É£¬ŃŠ¾æ±ķĆ÷Ö²ĪļŠčŅŖÄŚŌ“ŠŌĒāĘųĄ“µÖæ¹ÉųĶøŠ²ĘČ”£±©Ā¶ÓŚĒāĘų“Ģ¼¤ĮĖĮņ»ÆĒāµÄ²śÉś£¬²¢ČĻĪŖĪŖĮĖµ÷½ŚĘųæ×æŖ·Å¶Č“Ó¶ųµ¼ÖĀ¹Ū²ģµ½µÄÄĶŹÜŠŌ£¬Įņ»ÆĒāŹĒĒāĘųµÄĻĀÓĪ”£ŌŚĒŠ»ØŃŠ¾æÖŠŅ²·¢ĻÖĮĖĄąĖĘ½į¹ū”£ŌŚ¶ŌæµÄĖÜ°µÄŃŠ¾æÖŠ·¢ĻÖ£¬Ēā»ÆĆ¾ŗĶÄūĆŹĖįČÜŅŗŌö¼ÓĮĖĮņ»ÆĒāµÄ²śÉś”£ŗģŃõ»Æ»¹ŌĘ½ŗāµĆŅŌĪ¬³Ö£¬¶ųĖ„ĄĻ»łŅņµÄ±ķ“ļ±»ŅÖÖĘ”£Å£»ĒĖįŹĒŅ»ÖÖĮņ»ÆĒāĒå³ż¼Į£¬Äę×ŖĮĖÕāŠ©Š§Ó¦£¬²¢ČĻĪŖĒāĘųµÄĻĀÓĪŠ§Ó¦ŹĒÓÉĮņ»ÆĒā½éµ¼µÄ£¬ÕāÓėĘųæ×ŃŠ¾æŅ»ÖĀ”£
Ņņ“ĖæÉŅŌ擳ö£¬ĒāĘų²ĪÓėĮĖ°üĄØŅ»Ńõ»ÆµŖ”¢ŅŅĻ©ŗĶĮņ»ÆĒāµČŌŚÄŚµÄŅ»ĻµĮŠĘųĢåŠÅŗÅ·Ö×ӵĊÅŗÅ“«µ¼Ķ¾¾¶”£
Ī唢Ą“×Ō·ĒÖ²ĪļĄąĒāĘųŠ§Ó¦µÄĘō·¢
ŌŚÉśĪļ½ēÖŠ½ųŠŠæēĪļÖÖŃŠ¾ææÉÄÜ»į“ųĄ“»Ų±Ø£¬µ«Č·ŹµŠčŅŖ×¢ŅāŅ»Š©¾Æøę”£ĄżČē£¬¶ŌČĖĄąNAPDHŃõ»ÆĆøµÄ±ķÕ÷¼«“óµŲ“Ł½ųĮĖ¶ŌÖ²ĪļÖŠÕāŠ©ĆøĶ¬Ō“ĪļµÄŃŠ¾æµÄ½ųÕ¹”£ŹĀŹµÉĻ£¬Ą“×ŌÖ²ĪļŗĶ¶ÆĪļµÄŃõ»ÆĆøµ°°×æÉŅŌ×éŗĻŌŚŅ»Ęš£¬ŅŌĢåĶāÖŲ½Ø»īŠŌ”£ĮķŅ»·½Ćę£¬¶ÆĪļŅ»Ńõ»ÆµŖSµÄ·¢ĻÖÖ»ŌŚÖ²ĪļæĘѧ֊ŅżĘšĮĖÕłŅ锣“ĖĶā£¬Ö²Īļ֊ȱ·¦ŌŚ¶ÆĪļÖŠ±»³ä·ÖĆčŹöµÄsGCŠÅŗÅĶØĀ·½ųŅ»²½Ēæµ÷ĮĖæÉÄÜŠčŅŖ½÷É÷Ź¹ÓĆ”£»°ĖäČē“Ė£¬ĻĀĆęĢÖĀŪĮĖĶعż²ÉČ”¹ć·ŗµÄ·½·ØæÉŅŌѧµ½¶ąÉŁ¹ŲÓŚĒāĘųŌŚÉśĪļĻµĶ³ÖŠµÄ×÷ÓƵÄÖŖŹ¶”£
Čē¹ūĒāĘųÄܹ»ŌöĒæÖ²ĪļµÄÓ¦¼¤·“Ó¦£¬ÄĒĆ“Ļą¹ŲĻø°ūÖŠµÄĒāĘųÅØ¶ČŠčŅŖŌö¼Ó”£ÕāæÉŅŌĶعżĮ½ÖÖ»śÖĘŹµĻÖ£ŗŅŖĆ“ÄŚŌ“ŠŌĒāĘų²śÉśŌö¼Ó£¬ŅŖĆ“ĶāŌ“ŠŌ¹©Ó¦ĒāĘų”£
ŅĀŌåæÉÄÜŹĒŌåĄą×īÖŖĆūµÄÄŚŌ“ŠŌÉśĪļĻµĶ³Ö®Ņ»£¬ÓĆÓŚÉś²śĒāĘų”£ÕāÖÖÉśĪļ·Ē³£ÉĆ³¤²śÉśĒāĘų£¬ŅŌÖĮÓŚÓŠČĖ½ØŅ齫ĘäÓĆ×÷ÉśĪļČ¼ĮĻ”£ŅĀŌåĒāĘųµÄ²śÉśŹĒĶعżĒā»ÆĆøŹµĻֵģ¬ÕāÖÖ»śÖĘ×ī½üŅŃ¾µĆµ½ĮĖ×ŪŹö”£Čē¹ūæÉŅŌĶعż²Ł×ŻÕāÖÖĆøµÄæŲÖĘ»ņŌö¼Ó±ķ“ļŗĶĻą¹ŲµÄ¶ąėÄ»żĄŪĄ“Ōö¼ÓÖ²ĪļÖŠ»łÓŚĆøµÄĒāĘų²śÉś£¬ÄĒĆ“¾ĶæÉŅŌÓŠÕė¶ŌŠŌµŲĄūÓĆĒāĘųŠÅŗÅĄ“ŌöĒæÖ²ĪļµÄÉś³¤ŗĶÉś“ęÄÜĮ¦”£ÓŠŃ§Õߏ¹ÓĆÄ£ŠĶÉśĪļČēŅĀŌ壬ŅŌ¼°øüøß¼¶µÄÖ²ĪļÄ£ŠĶČēÄāÄĻ½ę½«ŌŚÕāĄąŃŠ¾æÖŠ·¢»ÓÖŲŅŖ×÷ÓĆ”£
ĮķŅ»ÖÖ·½·ØŹĒĶāŌ“ŠŌ¹©Ó¦ĒāĘų”£ČēÉĻĖłŹö£¬ÕāæÉÄÜĄ“×ŌÓŚČĖĪŖ»ī¶Æ£¬ĄżČēÓ¦ÓĆHRW”£Č»¶ų£¬Ö²ĪļÓėŠķ¶ąÉśĪļŅ»Ńł£¬æÉÄÜÓėŌŗĖÉśĪļŗĶÕę¾ś“¦ÓŚŠĶ¬»ņ¹²Éś×“Ģ¬£¬ĖüĆĒ±¾ÉķæÉŅŌ²śÉśĒāĘų”£ŌŚČĖĄąÖŠ£¬ÓŠŃŠ¾æ±ķĆ÷³¦µĄĪ¢ÉśĪļČŗ²śÉśµÄĒāĘųŌö¼ÓæÉÄÜÓŠŅęÓŚ½”æµ”£Ņņ“Ė£¬Ö²ĪļøłĻµÖÜĪ§ŌŗĖÉśĪļĒāĘų²śÉśµÄŌö¼ÓæÉÄܾßÓŠÓŠŅęµÄŠ§¹ū”£ĮķŅ»·½Ćę£¬ÓŠŃŠ¾æ±ķĆ÷ĒāĘųŃõ»ÆĶĮČĄĻø¾śŅ²ÓŠŅꓦ”£Ņņ“Ė£¬²Ł×ŻĶĮČĄĻø¾ś¾śČŗæÉÄÜŗÜø“ŌÓ£¬µ«øıäøłĻµø½½üµÄĒāĘų“śŠ»æÉÄÜ»įÓŠĪ“Ą“µÄŗĆ“¦”£
¶ŌĻø¾śµÄŃŠ¾æŅ²ÓŠÖśÓŚ½ŅŹ¾ĒāĘųŹĒČēŗĪĘš×÷ÓƵĔ£ÓÉÓŚĒāĘųż¾ßÓŠ·Ē³£µĶµÄ»¹ŌÖŠµćµēĪ»£Ø-414 mV£©£¬Ņņ“ĖŌŚÉśĪļĻµĶ³ÖŠ»¹ŌŠķ¶ąµ°°×ÖŹøØ»łæÉÄÜŹĒČČĮ¦Ń§ÉĻæÉŠŠµÄ”£ĶŃĮņ»”¾śÖŠĻø°ūÉ«ĖŲc3µÄ»¹Ō±Øøę¾ĶĖµĆ÷ĮĖÕāŅ»µć”£ÓŠČ¤µÄŹĒ£¬ŌŚĖęŗó½ųŠŠµÄŃõ»Æ»¹Ō·“Ó¦ÖŠ£¬ÓŠæÉÄÜ·¢ÉśĮņ»ÆĒāµÄ²śÉś£¬ŅŃÖŖĮņ»ÆĒāŹĒÖ²ĪļÖŠµÄÖŲŅŖĘųĢåŠÅŗÅ·Ö×Ó£¬°üĄØŌŚÓ¦¼¤Ģõ¼žĻĀ£¬ÕāÓė±¾ĪĵÄĢÖĀŪÓŠ¹Ų”£PeckŌŚ1959ÄźµÄŃŠ¾æ±ķĆ÷ĮĖĮ½µćÖŲŅŖÄŚČŻ”£Ź×ĻČ£¬ŌŚÉśĪļĻµĶ³ÖŠĒāĘų»¹ŌŃŖŗģĖŲĶÅ“ŲŹĒæÉÄܵĔ£Ęä“Ī£¬Ņ»µ©ŃŖŗģĖŲ±»»¹Ō£¬¾ĶÓŠæÉÄÜ·¢ÉśĻĀÓĪ·“Ó¦£¬“Ó¶ųæÉÄܲśÉśŠÅŗÅ·Ö×Ó”£ÕżČēŅŃ¾Ģį³öµÄ£¬ÕāŠčŅŖŌŚÖ²ĪļŗĶ¶ÆĪļÖŠ½ųŅ»²½Ģ½Ė÷£¬¶ų²»½ö½öŹĒŌŚŌŗĖÉśĪļÖŠ”£
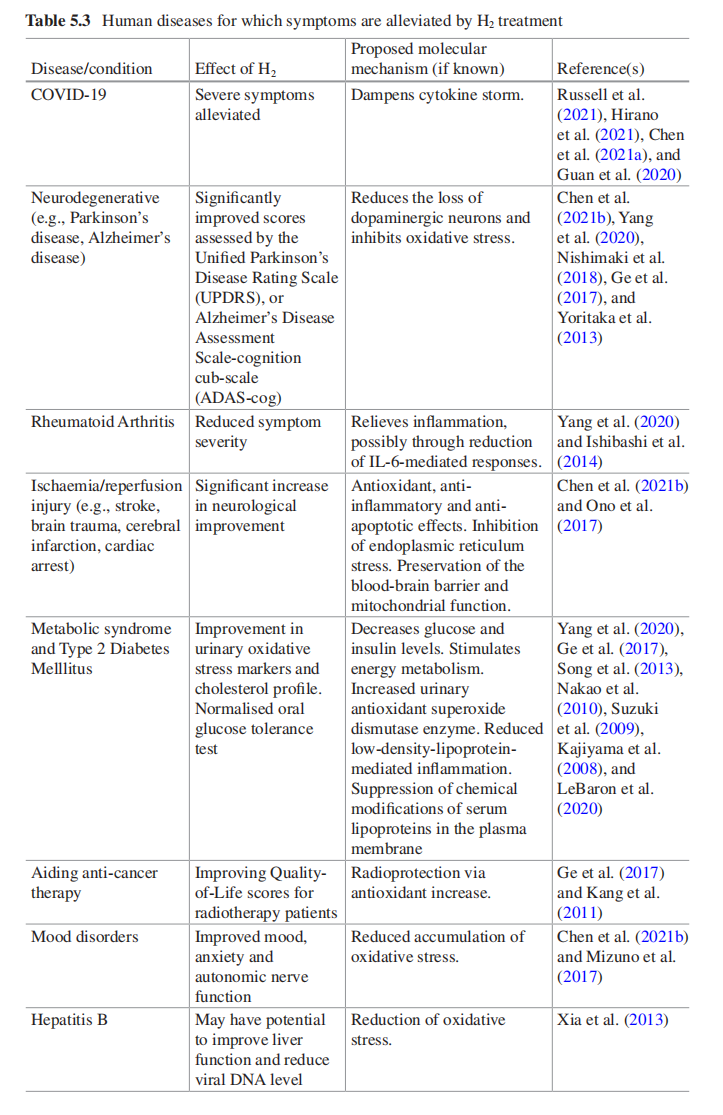
ĘäĖūĪļÖÖæÉŅŌŌŚÉśĪļŅ½Ń§ĮģÓņ·¢»Ó×÷ÓƵÄ×ī“óĮģÓņÖ®Ņ»¾ĶŹĒŌŚÕāĄļ”£ŌŚÕāĄļ£¬ŅŃ¾ÓŠŃŠ¾æ±ķĆ÷ĒāĘų¶Ōø÷ÖÖ¼²²”ÓŠŅę£¬°üĄØ±ķ5.3ÖŠĖłĮŠµÄ¼²²””£ŃŠ¾æ·¢ĻÖĒāĘųæÉŅŌ»ŗ½āCOVID-19µÄ֢ד²¢ŅŃÓĆÓŚĮŁ“²ŹŌŃ锣Ėü»¹±»·¢ĻÖ¶ŌÉń¾ĶĖŠŠŠŌ¼²²””¢Ąą·ēŹŖŠŌ¹Ų½ŚŃ×ŗĶĢĒÄņ²”ÓŠŅę”£Ņņ“Ė£¬ŗÜĆ÷ĻŌĒāĘų¶ŌČĖĢå½”æµŗĶ»ŗ½ā¼²²”֢ד¾ßÓŠŅ»ĻµĮŠŅꓦ”£“ĖĶā£¬Čē¹ūŅŃÖŖĒāĘųŌŚÉśĪļŅ½Ń§ĮģÓņµÄ»śÖĘ£¬ŹĒ·ńæÉŅŌ½«Ęä×Ŗ»Æ²¢ŌŚÖ²ĪļæĘѧ֊Ź¹ÓĆ£æ
±ķ5.3 ĒāĘųÖĪĮĘ»ŗ½āµÄČĖĄą¼²²”֢ד
²»½öŹĒ±ķ5.3ÖŠµÄŹż¾ŻÖ§³ÖĮĖĒāĘųŌŚÉśĪļĻµĶ³ÖŠ²śÉśÉīŌ¶Ó°ĻģµÄ¹Ūµć£¬¶ųĒŅĒāĘųĘš×÷ÓƵķ½Ź½Ņ²ŹĒĻą¹ŲµÄ”£ĻŌČ»£¬¶ÆĪļÖŠµÄŅ»Š©×÷ÓĆŗĶ»śÖĘ²¢²»Ö±½ÓŹŹÓĆÓŚÖ²Īļ”£ĄżČē£¬½µµĶIL-6Ė®Ę½»ņŅÖÖĘĻø°ūŅņ×Ó·ē±©µÄæ¹Ń×Ö¢×÷ÓĆ²»»įŌŚÖ²ĪļÖŠ³öĻÖ”£Č»¶ų£¬ĒāĘųæÉÄÜ»įÓ°ĻģÖ²ĪļÖŠµÄĄąĖĘ°ū¼äŠÅŗÅ·Ö×Ó£¬ČēŅŅĻ©”£ĘäĖū×÷ÓĆæÉÄÜøüĪŖĻą¹Ų”£ČēĒ°ĖłŹö£¬ĒāĘųæÉÄÜĶعżĘä¶Ōæ¹Ńõ»Æ¼ĮµÄ×÷ÓĆĘš×÷ÓĆ£¬ÕāŅ»Š§Ó¦ŅŃ¾ŌŚÖ²ĪļÖŠ¹Ū²ģµ½¹ż”£Ņņ“Ė£¬æ¹Ńõ»Æ¼ĮµÄ±ä»ÆŗĶ¶ŌŃõ»ÆÓ¦¼¤µÄŅÖÖĘŹĒÉń¾ĶĖŠŠŠŌ¼²²”»ŗ½ā”¢ĢĒÄņ²”ÖĪĮĘ”¢ĒéŠ÷øıäŗĶŅŅŠĶøĪŃ׵Ĺ²Ķ¬ĢŲÕ÷£¬ÕāŅ²ŹĒĒāĘų¼õĒįÖ²ĪļÓ¦¼¤µÄ¹²Ķ¬ĢŲÕ÷”£¶ÆĪļŗĶÖ²ĪļĻø°ūµÄÉś»ÆĻø½ŚÓŠĖł²»Ķ¬£¬µ«»ł±¾ŌĄķ“ęŌŚĄąĖĘŠŌ”£Ņņ“Ė£¬ĆÜĒŠŃŠ¾æ¶ÆĪļѧŗĶŅ½Ń§ĮģÓņÖŠµÄĒāĘųŃŠ¾æ¶ŌĪ“Ą“Ö²ĪļæĘѧæÉÄÜ»į·Ē³£ÓŠŅę£¬·“Ö®ŅąČ»”£¼ųÓŚĒāĘųæÉÄÜÓŠŅęÓŚČĖĄą½”浵ļ²²”ĮŠ±ķ£¬ÄŃ¹ÖČĖĆĒŅŃ¾æŖŹ¼æ¼ĀĒ½«ĒāĘų×÷ĪŖĪ“Ą“µÄČĖĄąĮĘ·Ø”£æÉÄÜŹĒÓÉÓŚCOVID-19“óĮ÷ŠŠ²”µÄŌŅņ£¬ÄæĒ°ŃŠ¾æŗĶÓ¦ÓĆÖŲµćŌŚÓŚŗōĪüĻµĶ³¼²²”£¬ĪŽĀŪČēŗĪĒāĘųµÄŃŠ¾æŗĶÓ¦ÓĆŌŚÉśĪļŅ½Ń§ĮģÓņæÉÄÜ»į³ÖŠųŹÜµ½¹Ų×¢”£ĮķŅ»·½Ćę£¬ŅŃ¾ÓŠŃŠ¾æ±ķĆ÷ĒāĘųµÄÓ¦ÓƶŌÅ©ŅµæÉÄÜ·Ē³£ÓŠŅę”£Ö²ĪļŗĶ¶ÆĪļÖ®¼äæÉÄÜ“ęŌŚ¹²Ķ¬µÄ·Ö×Ó»śÖĘĄ“ĻģÓ¦ĒāĘųµÄ×÷ÓĆ£¬ĪŽĀŪŹĒĶعżæ¹Ńõ»Æ¼Į»¹ŹĒFe3+»¹Ō»ņĘäĖū·½Ź½”£Ö²ĪļæĘѧæÉÄÜÓŠŗܶąæÉŅŌ“ÓŌŗĖÉśĪļŗĶøüøß¼¶µÄ¶ÆÖ²ĪļŃŠ¾æ֊ѧµ½µÄ¶«Ī÷£¬·“Ö®ŅąČ»”£
Įł”¢½įĀŪŗĶĒ°¾°
ĻÖŌŚæ“Ą“£¬ĒāĘų¶ŌÖ²ĪļŹĒŅ»ÖÖÓŠÓĆµÄ¹¦Äܵ÷½ŚŗĶ¼²²”·ĄÖĪ·½·Ø£¬æÉŅŌ»ŗ½āŅ»ĻµĮŠŃ¹Į¦ĢōÕ½£Ø±ķ5.1£©£¬Ņ²ŹĒĖ®¹ūŗĶ»Ø»ÜŹÕ»ńŗ󓢓ęµÄĒ±ŌŚ·½°ø£¬ŅņĪŖĖüæÉŅŌĆ÷ĻŌŃÓ»ŗĖ„ĄĻ”£Č»¶ų£¬ĒāĘųµÄÉśĪļ»ÆѧӊŠķ¶ą·½ĆęÉŠ²»Ē峞”£Ź×ĻČ£¬ĖäČ»ŅŃ¾Ģį³öĮĖ¼øÖÖ»śÖĘ£Ø±ķ5.2£©£¬°üĄØĒå³ż×ŌÓÉ»łŗĶĘäĖū·“Ó¦ŠÅŗÅ£¬»ņĶعżHO-1Ęš×÷ÓĆ£¬µ«ĒāĘųŌŚĻø°ūÖŠµÄÖ±½Ó°Š±źÉŠ²»Ē峞”£Ęä“Ī£¬ĖäȻӊŠķ¶ą¹ŲÓŚĒāĘųÓ¦ÓƵıØøęŹĒÓŠŅęµÄ£Ø±ķ5.1£©£¬µ«ĒāĘųµÄČ«²æ×÷ÓĆÉŠ²»Ē峞”£
ÓėĘäĖūÉśĪļ·Ö×ÓĻą±Č£¬ĒāĘųµÄŃõ»Æ»¹ŌµēĪ»Ļą¶Ō½ĻµĶ”£“ÓČČĮ¦Ń§½Ē¶ČĄ“æ“£¬ĒāĘųæÉŅŌ½«Fe3+»¹ŌĪŖFe2+£¬Õā¶ŌŠķ¶ąĆø¶¼ÓŠÓ°Ļģ£¬Õā±ķĆ÷øØ»łµÄ»¹Ō£¬ĢŲ±šŹĒŠķ¶ąŃŖŗģĖŲ»łĶŵĻ¹Ō£¬ŌŚĄķĀŪÉĻŹĒÓŠæÉÄܵģ¬¾”¹Üƻӊ¹ć·ŗ±ØµĄ”£“ĖĶā£¬°“ÕÕĄąĖʵÄĀß¼£¬ŗ¬ĪųĆøŅ²æÉÄÜŹĒĒāĘųµ÷æŲµÄÄæ±ź”£Č»¶ų£¬Čē¹ūƻӊĢŲ¶ØµÄ»·¾³Ą“½µµĶÕāÖÖ»¹Ō·¢ÉśµÄ»ī»ÆÄÜ£¬ÕāÖÖ·“Ó¦ŌŚ¶ÆĮ¦Ń§ÉĻæÉÄÜŹĒ²»æÉŠŠµÄ”£Ņņ“Ė£¬»¹ŠčŅŖ¶ŌĒāĘųæŲÖʵĵ°°×ÖŹ½ųŠŠČ«ĆęŃŠ¾æ£¬¼“Ź¹Ö»ŹĒ½«ĘäÅųżŌŚĶā”£ÓėŅ»Ńõ»ÆµŖŗĶĮņ»ÆĒāµÄŃŠ¾æ²»Ķ¬£¬µ°°×ÖŹ×éѧ·½·ØĖĘŗõ²»ŹŹÓĆÓŚĒāĘų”£ŅņĪŖÄæĒ°»¹Ć»ÓŠ¹ŲÓŚøĆ·Ö×ÓÖ±½Ó¹²¼Ū·ŅėŗóŠŽŹĪµÄ±ØµĄ”£ĮķŅ»·½Ćę£¬ĻĀÓĪ·Ņėŗóµ°°×ÖŹŠŽŹĪ½«»į·¢Éś£¬ĮĖ½āÕāŠ©Š§Ó¦µÄČ«Ģ×øŁŅŖ½«ŹĒÓŠÓƵĔ£
ĖäČ»Ņ»Š©Ö²ĪļÄŚŌ“ŠŌĒāĘųµÄ²śÉśŹĒæÉÄܵģ¬²¢ĒŅæÉŅŌ¼ÓŅŌæŲÖĘ£¬µ«ĶāŌ“ŠŌĒāĘųµÄæŲÖĘÓ¦øĆŹĒøüŗƵďµÓĆ·½·Ø£¬ŅņĪŖĶāŌ“ŠŌĒāĘųøüČŻŅ×Ģį¹©ŗĶµ÷½Ś”£ĒāĘųæÉŅŌ×÷ĪŖÖĪĮĘĘųĢåÓ¦ÓĆÓŚÖ²Īļ£¬Ņ²æÉÓÉÖ²ĪļÖÜĪ§µÄĪ¢ÉśĪļČŗµ÷æŲ¹©Ó¦”£Ź¹ÓĆĘųĢå·½Ź½²»Ģ«æÉÄÜÓŠÓĆ£¬µ«ĶعżHRW»ņHNWæÉÄÜŌŹŠķÓ¦ÓĆÓŚŅ¶×Ó»ņøł²æ£¬»ņĮ½Õß¼ę¶ųÓŠÖ®”£ĻŌČ»£¬“ÓĪļĄķ½Ē¶ČĄ“æ“£¬“ęŌŚ°²Č«·½ĆęµÄĪŹĢā£¬ŅņĪŖĒāĘų¼«Ņ×Č¼£¬µ«“ÓÉśĪļ½Ē¶ČĄ“æ“£¬ĒāĘųĖĘŗõ¶ŌÖ²ĪļŗĶ¶ÆĪļ¶¼ŹĒ°²Č«µÄ”£ÓėĘäĖūĄąĖĘ·Ö×Ó£ØĄżČēĮņ»ÆĒā£©Ņ»Ńł£¬¹©Ģå·Ö×ÓæÉŅŌøüŗƵŲĄūÓĆĒāĘų”£ŌŚĪ“Ą“£¬Ņ»Š©ÓĆĶ¾ŅŃ¾±»±ØµĄ£¬ČēĆ¾Ēā»ÆĪļŗĶAB@hMSN”£
ĒāĘųŌŚÅ©ŅµŗĶŌ°ŅÕÖŠµÄÓ¦ÓĆÉŠĪ“µĆµ½¹ć·ŗ²ÉÓĆ£¬µ«ČĖĆĒ¶ŌÕāÖÖÉśĪļ°²Č«“¦Ąķ·½·ØµÄŠĖȤČÕŅęÅØŗń”£Ėę×ÅČĖĆĒ¶ŌĒāĘųČēŗĪĘš×÷ÓĆŅŌ¼°ĘäÓ°ĻģµÄÖŲŅŖŠŌµÄĮĖ½āŌ½Ą“Ō½¶ą£¬ĒāĘųæÉÄÜ»į³ÉĪŖĪ“Ą““Ł½ųÖ²ĪļÉś³¤ŗĶ×÷Īļ“¢“ęµÄŅ»ÖÖæÉ½ÓŹÜµÄ·½·Ø”£
https://blog.sciencenet.cn/blog-41174-1412385.html
ÉĻŅ»ĘŖ£ŗ³ĒŹŠ»ÆČĖČŗøōĄėć£ĀŪ
ĻĀŅ»ĘŖ£ŗĶęĖ£ŹĒŗ¢×ÓĆĒµÄøÕŠč
