博文
肥胖诱导高血压因血管内皮细胞上的蛋白Panx3被抑制  精选
精选
|
肥胖诱导高血压因血管内皮细胞上的蛋白Panx3被抑制
肥胖会诱发内皮功能障碍,从而导致高血压。Wolpe 等人发现了 Panx3 作为限制内皮和高血压氧化应激的支架蛋白的作用。Panx3 与转录抑制因子 Bcl6 结合并稳定,Bcl6 抑制了 Nox4 的表达,Nox4 是编码过氧化氢产生酶的基因。内皮细胞中缺乏Panx3或用破坏Panx3-Bcl6相互作用的肽处理的小鼠在内皮细胞中表现出更大的氧化应激并发展为高血压。此外,饮食诱导肥胖小鼠的Panx3 mRNA表达和Bcl6蛋白丰度降低,Nox4 mRNA表达增加,但药理诱导高血压小鼠没有增加。内皮 Panx3 和 Bcl6 丰度在高血压、肥胖个体中降低,表明内皮 Panx3 的减少可能导致肥胖相关高血压。
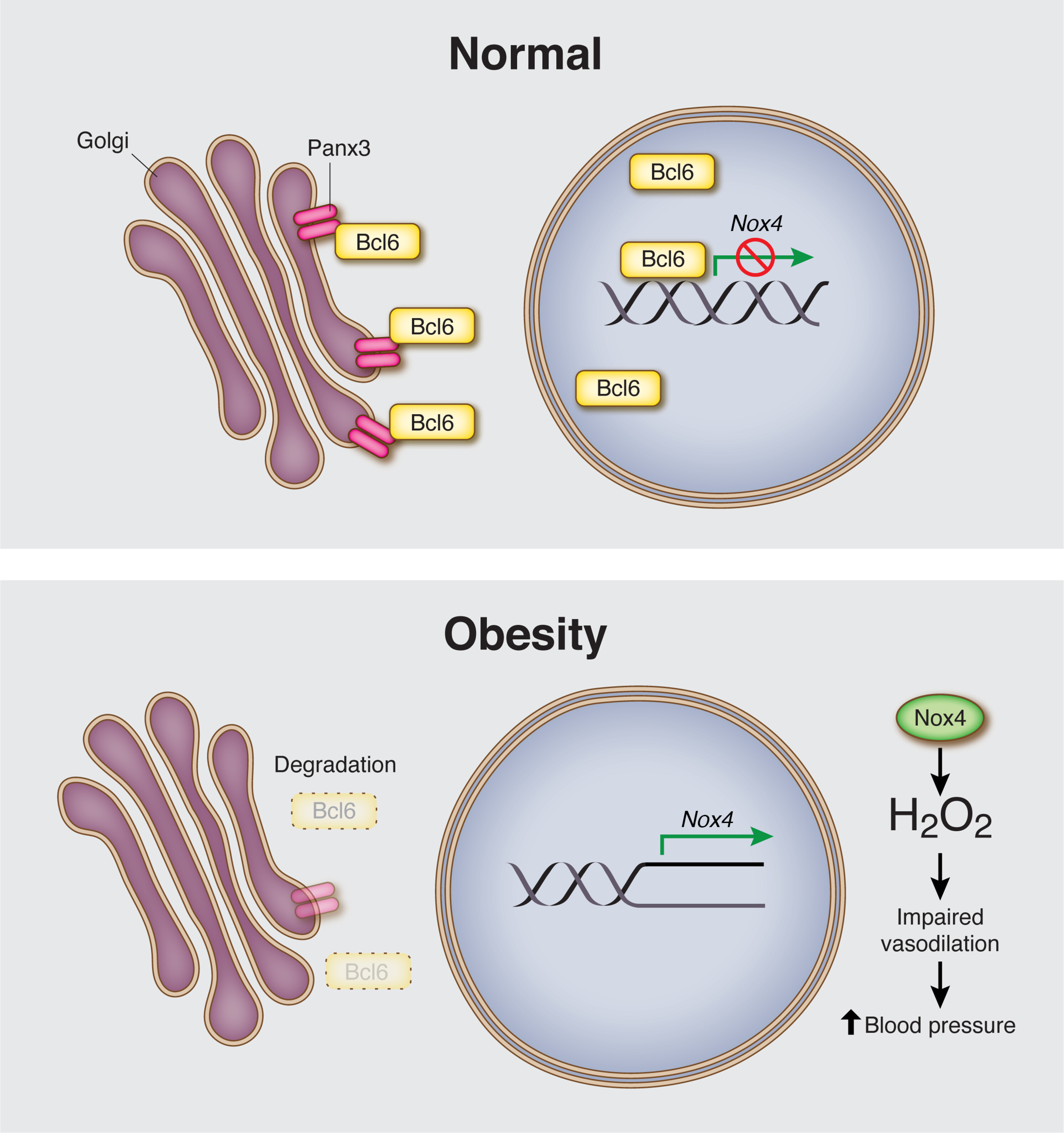
图Panx3/Bcl6 相互作用在内皮细胞中的作用示意图。
在正常生理条件下,Panx3存在于高尔基体内皮膜上,在那里它可以与Bcl6的细胞质池相互作用,通过抑制Bcl6的靶向降解有效地增加Bcl6的丰度。Nox4 表达受核定位 Bcl6 的转录抑制限制。在肥胖中,内皮 Panx3 的丰度降低,这与 Bcl6 丰度和活性的降低有关。在没有 Bcl6 抑制的情况下,Nox4 表达增加,导致 H 产生过剩2O2,损害内皮介导的扩张,并最终导致全身血压升高。
靶向降解调节转录阻遏蛋白 Bcl6 的活性及其抑制氧化应激和炎症的能力。在这里,我们报告了内皮 Bcl6 的丰度取决于其与高尔基体定位的膜联蛋白 3 (Panx3) 的相互作用,并且 Bcl6 转录活性可防止血管氧化应激。与肥胖、高血压人类的数据一致,Panx3 中具有内皮细胞特异性缺陷的小鼠患有自发性全身性高血压,但通道功能、Ca 2+处理、ATP 量或高尔基体管腔 pH 值没有明显变化。Panx3 与 Bcl6 结合,其缺失降低了 Bcl6 蛋白丰度,表明与 Panx3 的相互作用通过阻止 Bcl6 降解来稳定 Bcl6。Panx3 缺乏与编码H2O2产生酶 Nox4的基因表达增加有关,通常被Bcl6 抑制,导致 H2O2-诱导脉管系统的氧化损伤。过氧化氢酶挽救了缺乏内皮Panx3的小鼠的血管舒张受损。施用新开发的肽以抑制 Panx3-Bcl6 相互作用,概括了内皮性 Panx3 缺乏症小鼠中 Nox4 表达和血压的增加。Panx3-Bcl6-Nox4失调发生在肥胖相关的高血压中,但在没有肥胖的情况下诱发高血压时则不然。我们的研究结果提供了对 Panx3 的通道非依赖性作用的见解,其中它与 Bcl6 的相互作用决定了血管氧化状态,特别是在肥胖的不利条件下。
活性氧化物质 (ROS) 是氧代谢的结果,包括高活性自由基,例如超氧化物 (O2−),以及更稳定的非自由基过氧化物物质,如过氧化氢(H2O2).ROS是分子双刃剑;尽管它们是各种生理信号传导过程不可或缺的一部分,但ROS的过量产生可以超过抗氧化防御,对心血管系统和其他组织造成损害。例如,线粒体 ROS 有助于人动脉中血流介导的扩张 (1),但不能充分清除线粒体 O2−损害一氧化氮 (NO) 生物利用度,从而诱发高血压和动脉粥样硬化 (2, 3)。ROS 失衡的影响在肥胖症中尤为严重,肥胖症会加剧氧化应激并持续血管功能障碍,以及与肥胖相关的过氧亚硝酸盐失衡(ONOO−) 在内皮细胞中损害内皮细胞功能并驱动高血压 (4)。因此,ROS生成和抗氧化机制之间错综复杂的相互作用需要严格的监管。参与氧化应激的基因的转录调控是维持稳态氧化还原平衡的核心,这在肥胖等疾病状态下变得更加重要,这些疾病状态本质上会使天平向氧化应激倾斜。
B 细胞淋巴瘤 6 (Bcl6) 是一种转录因子,可差异调节氧化应激源和氧化还原蛋白的表达。作为其在体液反应中充分证明的作用的一部分,Bcl6 转录活性可在生发中心形成的遗传毒性和氧化攻击期间促进 B 细胞存活 (5–8)。在非霍奇金淋巴瘤中,淋巴瘤中 Bcl6 的过表达通过阻止诱导细胞凋亡的 ROS 的产生来促进对化疗药物的耐药性 (9)。同样,弥漫性大 B 细胞淋巴瘤细胞的一个亚群会经历向氧化磷酸化的代谢转变,这需要硫氧还蛋白的 Bcl6 依赖性表达来耐受氧化激发反应 (10)。Bcl6 的氧化保护不仅限于其直接改变氧化剂和还原剂表达的能力。Bcl6 与促氧化、促炎转录因子 NF-κB 表现出相互拮抗作用,可抑制 NF-κB 激活的许多转录靶标 (11, 12)。Bcl6 参与多种蛋白质-蛋白质相互作用以履行这些作用。在胞质溶胶中,丝裂原活化蛋白激酶 (MAPK) 可以磷酸化 Bcl6 中的脯氨酸、谷氨酸、丝氨酸和苏氨酸 (PEST) 基序,导致转录因子在通过与 FBXO11 相互作用募集到 SCF(Skp1、Cullin 和 F-box 蛋白)泛素连接酶后蛋白酶体降解 (13, 14).同样,Bcl6 的转录抑制可以通过过氧化物酶体增殖物激活受体 (PPAR) 的隔离来调节,PPAR 是调节脂质和葡萄糖代谢的核激素受体 (15–17),突出了转录因子对生理系统的多种影响。然而,尚未确定所有 Bcl6 相互作用的功能角色。例如,对 Bcl6 结合伴侣的无偏筛选发现了与 pannexin 3 (Panx3) 的相互作用 (18),尽管这种相互作用的影响尚不清楚。
pannexin 通道家族由三种亚型组成:Panx1、Panx2 和 Panx3。Panx1 在质膜上具有充分证明的作用,有助于嘌呤能、肾上腺素能和 Ca2+脉管系统中的信号转导 (19–27)。相比之下,Panx3的作用仍然相对难以捉摸。尽管根据 RNA 测序 (RNA-Seq) 数据集,已在整个全身微血管系统中检测到其表达水平较低 (28),但其功能属性尚未完全阐明。已发表的关于Panx3的定位和功能意义的报告表明,Panx3在细胞类型和表达系统中具有高度的可塑性。当定位于内质网时,Panx3 被认为有助于 Ca2+商店发布 (29)。然而,据报道,Panx3 也定位于质膜,与染料摄取 (30–32) 和腺苷 5′-三磷酸 (ATP) 释放有关 (29, 33)。在这里,我们证明了 Panx3 的通道非依赖性功能,因此它与高尔基体定位的 Bcl6 池的相互作用可以决定氧化应激相关基因的转录抑制。我们的研究结果为肥胖相关高血压中内皮功能障碍的发展提供了见解。
https://blog.sciencenet.cn/blog-41174-1419996.html
上一篇:阿尔茨海默病可传染性的意外证据
下一篇:癌症劫持神经系统生长和扩散
