ВЉЮФ
ЩњЮяФмРэТлЕФгІгУЧАОАЃЈ4ЃЉ
||
ЦпЁЂЫЅРЯЕФЯпСЃЬхРэТлЃКФмСПВњЩњгыбѕЛЏЫ№ЩЫЕФРлЛ§
ЫЅРЯЕФЯпСЃЬхРэТлЃЌЪЧЯжДњЫЅРЯЩњЮябЇжазюжїСїЧвБЛЙуЗКНгЪмЕФРэТлПђМмжЎвЛЁЃЫќКЫаФжїеХЃКЫЅРЯВЂЗЧвЛИіБЛвХДЋГЬађдЄЯШЩшЖЈЕФЙ§ГЬЃЌЖјЪЧЯпСЃЬхетвЛЁАФмСПКЭздгЩЛљЙЄГЇЁБдкТўГЄвЛЩњжаЃЌЦфЙІФмж№НЅЫЅЭЫКЭгаКІИБВњЦЗГжајЛ§РлЫљЕМжТЕФБиШЛНсЙћЁЃ
ЦфБОжЪЪЧвЛИіЁАздЮвЗХДѓЕФЖёадбЛЗЁБЃЌОпЬхЙ§ГЬПЩвдЭЈЙ§ЭМ2РДРэНтЃК
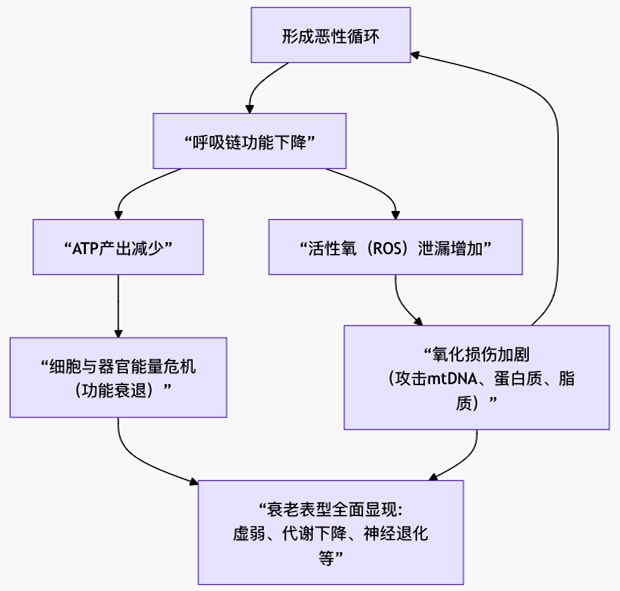
ЭМ2 ЫЅРЯЕФЯпСЃЬхРэТлЃКФмСПВњЩњгыбѕЛЏЫ№ЩЫЕФРлЛ§
ЭМ2ЧхЮњЕиеЙЪОСЫРэТлЕФКЫаФТпМСДЬѕЃЌЯТУцЮвУЧРДж№вЛВ№НтЦфжаЕФЙиМќЛЗНкЁЃ
1. РэТлЕФКЫаФжЇжљ
ЃЈ1ЃЉЯпСЃЬхDNAЕФДрШѕадгыЭЛБфРлЛ§
ЮЛжУСгЪЦЃКЯпСЃЬхDNAНєСкВњЩњДѓСПROSЕФКєЮќСДЃЌЧвШБЗІзщЕААзБЃЛЄКЭИпаЇЕФаоИДЯЕЭГЃЌвђДЫЫ№ЩЫТЪЪЧКЫDNAЕФ10-100БЖЁЃ
ИДжЦДэЮѓЃКЫцзХЯИАћЗжСбКЭЯпСЃЬхздЩэИќаТЃЌИДжЦДэЮѓВЛЖЯРлЛ§ЃЌЕМжТБрТыКєЮќСДЕААзЕФЛљвђГіЯжЕуЭЛБфКЭШБЪЇЁЃетаЉЭЛБфЛсЯёЁАгаШБЯнЕФСуМўЁБвЛбљБЛзщзАНјКєЮќСДЁЃ
ЃЈ2ЃЉКєЮќСДЙІФмеЯАгыROSЕФЖёадбЛЗ
КЌгаШБЯнЕААзЕФКєЮќСДИДКЯЮяЃЈгШЦфЪЧИДКЯЮяIКЭIIIЃЉаЇТЪЕЭЯТЃЌЕчзгИќШнвзЁАТЉГіЁБЃЌгыбѕЗжзгНсКЯЩњГЩГЌбѕвѕРызгЕШROSЁЃ
етаЉдіМгЕФROSЛсНјвЛВНЙЅЛїИННќЕФmtDNAКЭКєЮќСДЕААзБОЩэЃЌдьГЩИќЖрЫ№ЩЫКЭИќЕЭаЇТЪЃЌаЮГЩЩЯЮФЭМжаФЧИіВЛЖЯЖёЛЏЕФе§ЗДРЁбЛЗЁЃ
ЃЈ3ЃЉФмСПЪфГіЯТНЕгыЯИАћЫЅЭі
КєЮќСДећЬхЙІФмЫЅЭЫЃЌЕМжТжЪзгЬнЖШНЈСЂаЇТЪКЭATPКЯГЩЫйТЪЯТНЕЁЃ
ИпКФФмзщжЏЃЈШчДѓФдЁЂМЁШтЁЂаФдрЃЉЕФЯИАћзюЯШИаЪмЕНЁАФмдДЮЃЛњЁБЃЌЙІФмж№НЅМѕЭЫЁЃЕБЯИАћФмСПЕЭгкЮЌГжЛљБОЮШЬЌЕФуажЕЪБЃЌЛсДЅЗЂЕђЭіЛђЫЅРЯЁЃ
ЃЈ4ЃЉжЪСППижЦЯЕЭГЕФЫЅЭЫ
ФъЧсЯИАћЭЈЙ§ЯпСЃЬхздЪЩетЬзЁАжЪМьЛиЪеЯЕЭГЁБМАЪБЧхГ§ЪмЫ№ЯпСЃЬхЁЃЕЋЫЅРЯЙ§ГЬжаЃЌздЪЩФмСІЦеБщЯТНЕЃЌЕМжТЙІФмЪЇЕїЕФЯпСЃЬхдкЯИАћФкЖбЛ§ЃЌгЬШчЁАРЌЛјЗЂЕчГЇЁБГжајжЦдьЮлШОКЭЕЭаЇФмдДЁЃ
2. РэТлЕФЬєеНгыЯжДњаое§
ИУРэТлЫфШЛЧПДѓЃЌЕЋВЂЗЧЭъУРЃЌе§ОРњзХПЦбЇжЄОнЕФМьбщгыаое§ЃК
ЃЈ1ЃЉвђЙћадељвщЃКвЛаЉвХДЋбЇЪЕбщЗЂЯжЃЌШЫЮЊдіМгmtDNAЭЛБфШЗЪЕФмМгЫйЫЅРЯЃЌЕЋдіМгбѕЛЏЫ№ЩЫШДЮДБиЁЃетБэУїmtDNAЭЛБфБОЩэПЩФмЪЧБШЙувхбѕЛЏЫ№ЩЫИќЙиМќЕФЧ§ЖЏвђЫиЁЃ
ЃЈ2ЃЉаХКХЕФИДдгадЃКбаОПЗЂЯжЃЌЕЭЫЎЦНROSВЛНіЪЧЫ№ЩЫвђзгЃЌвВЪЧживЊЕФДйЪЪгІадаХКХЃЈШчМЄЛюПЙбѕЛЏЗРгљЃЉЁЃЫЅРЯПЩФмЪЧЁАаХКХЪЇЕїЁБЖјЗЧЕЅДПЕФЁАЫ№ЩЫРлЛ§ЁБЁЃ
ЃЈ3ЃЉЯЕЭГадЕФЪгНЧЃКЯпСЃЬхВЂЗЧЙТСЂЫЅРЯЁЃЫќгыКЫЛљвђзщЮШЖЈадЁЂЕААзжЪЮШЬЌЯЕЭГЁЂгЊбјИажЊЭЈТЗЃЈШчmTORЃЌ AMPKЃЉЕФЯрЛЅзїгУЙВЭЌЙЙГЩСЫЫЅРЯЕФЭјТчЁЃЯпСЃЬхЫЅЭЫМШЪЧвђЃЌвВЪЧЙћЁЃ
3. ЧАбиИЩдЄЃКЛљгкРэТлЕФПЙЫЅРЯВпТд
ОЁЙмДцдкЬєеНЃЌИУРэТлШдЮЊИЩдЄЫЅРЯЬсЙЉСЫзюИЛГЩЙћЕФТЗОЖЁЃВпТдКЫаФдкгкЁАДђЦЦЖёадбЛЗЃЌдіЧПЯЕЭГЙІФмЁБЃЈБэ10ЃЉЁЃ
Бэ10 ЛљгкЯпСЃЬхРэТлЕФПЙЫЅРЯВпТд
ИЩдЄВпТд | ДњБэЗНЗЈ/ЮяжЪ | зїгУЛњжЦгыФПБъ |
МѕЧсбѕЛЏЫ№ЩЫ | ЯпСЃЬхАаЯђПЙбѕЛЏМСЃЈШчMitoQЃЌ SS-31ЃЉ | НЋПЙбѕЛЏМСжБНгЕнЫЭжСЯпСЃЬхЛљжЪЃЌжаКЭROSЃЌБЃЛЄmtDNAКЭЕААзжЪЁЃ |
ЬсЩ§NAD+ЫЎЦН | ВЙГфNAD+ЧАЬхЃЈШчбЬѕЃАЗКЫЬЧЃЌNRЃЉ | NAD+ЪЧSIRTЕШГЄЪйЕААзЕФЕзЮяЃЌЬсЩ§ЦфЫЎЦНПЩдіЧПЯпСЃЬхздЪЩЁЂЩњЮяКЯГЩКЭаоИДЁЃ |
діЧПЯпСЃЬхздЪЩ | Urolithin AЃЈUrolithin AЃЉЁЂРзХСУЙЫи | МЄЛюЯИАћЧхГ§ЪмЫ№ЯпСЃЬхЕФФмСІЃЌИќаТЯпСЃЬхШКЬхЁЃ |
ДйНјЯпСЃЬхЩњЮяКЯГЩ | дЫЖЏЖЭСЖЁЂAMPKМЄЖЏМСЃЈШчЖўМзЫЋывЃЉ | ЭЈЙ§МЄЛюPGC-1ІСЕШжїПивђзгЃЌгеЕМВњЩњаТЕФЁЂНЁПЕЕФЯпСЃЬхЁЃ |
МѕЩйmtDNAЭЛБф | ЛљвђжЮСЦЃЈШчАаЯђБрМЃЉ | ЩаДІдчЦкЃЌФПБъЪЧжБНгОРе§ЛђЯћГ§РлЛ§ЕФжТВЁадЭЛБфЁЃ |
4. змНс
ЫЅРЯЕФЯпСЃЬхРэТлЬсЙЉСЫвЛИіЧПДѓЁЂздЧЂЧвПЩМьбщЕФПђМмЃЌЫќНЋФмСПДњаЛЁЂздгЩЛљЩњЮябЇКЭвХДЋЫ№ЩЫСЊЯЕЦ№РДЃЌНтЪЭСЫЮЊКЮИпКФФмЦїЙйзюЯШЫЅЭЫЃЌвдМАЮЊКЮЫЅРЯгыЖржжТ§адВЁЙВЯэВЁРэЛљДЁЁЃ
ЫфШЛЦфЯИНкЃЈШчROSЕФКЫаФНЧЩЋЃЉе§БЛИќОЋЯИЕФбаОПаое§ЃЌЕЋЁАЯпСЃЬхЙІФмЫцСфЫЅЭЫЪЧЫЅРЯЕФКЫаФЧ§ЖЏСІЁБетвЛКЫаФЙлЕувбБЛЙуЗКНгЪмЁЃРэНтетвЛРэТлЃЌВЛНіШУЮвУЧПДЧхЫЅРЯЕФЁАв§ЧцЁБШчКЮТ§Т§ЯЈЛ№ЃЌИќжИУїСЫЭЈЙ§ЮЌЛЄЯИАћФмСПДњаЛНЁПЕРДбгЛКЫЅРЯЁЂДйНјНЁПЕРЯСфЛЏЕФПЦбЇЕРТЗЁЃетЛђаэЪЧФњДгЁАЛюСІТлЁБЕНЁАЩњЮяФмбЇЁБЫМЯыжЎТУжаЃЌзюОпЯжЪЕвтвхЕФвЛИіжеЕуЁЃ
АЫЁЂГЌдНИіЬхЃКЩњЬЌЯЕЭГжаЕФФмСПСї
дкЩњЬЌЯЕЭГжаЃЌФмСПСїЖЏзёбгыЯИАћФкЯрЭЌЕФШШСІбЇЬњТЩЃЌЕЋЦфЙцФЃКъДѓЁЂТЗОЖИДдгЃЌЙЙГЩСЫЧ§ЖЏећИіЩњЮяШІдЫзЊЕФЁАЩњУќжЎСїЁБЁЃетЪЧЩњЮяФмбЇдРэДгЗжзгГпЖШЕНШЋЧђГпЖШЕФжеМЋбгЩьЁЃ
ЩњЬЌЯЕЭГжаЕФФмСПСїЃЌЦфКЫаФЬиеїЪЧЁАЕЅЯђСїЖЏЁЂж№МЖЕнМѕЁБЃЌВЂбЯИёзёбШШСІбЇЖЈТЩЁЃЮЊСЫжБЙлЕиАбЮеетвЛКъЙлЙ§ГЬЃЌЭМ3еЙЪОСЫФмСПдкЩњЬЌЯЕЭГжаСїЖЏгызЊЛЛЕФШЋОАТЗОЖЃК
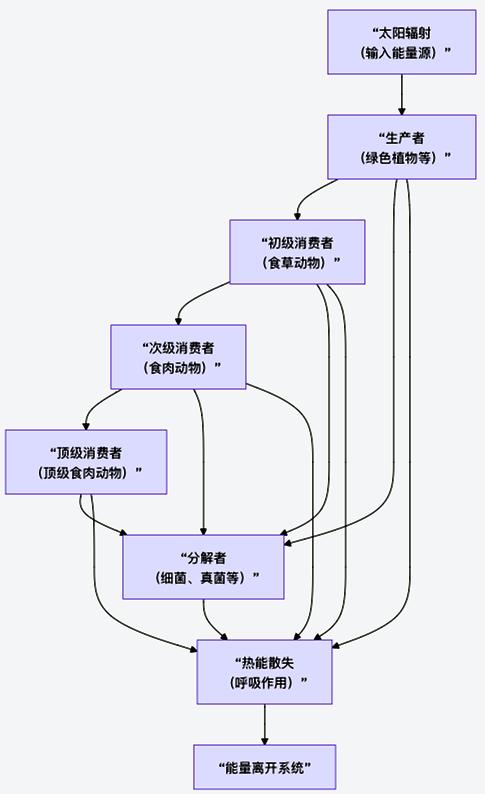
ЭМ3 ФмСПСїЖЏЕФЕЅЯђадгыКФЩЂад
ЭМ3НвЪОСЫФмСПСїЖЏЕФЕЅЯђадгыКФЩЂадЁЃЯТУцЃЌЮвУЧРДНтЮіетвЛЙ§ГЬЕФКЫаФЛЗНкгыЖЈСПЙцТЩЁЃ
1. ФмСПСїЕФЦ№ЕуЃКГѕМЖЩњВњ
ФмСПЪфШыЩњЬЌЯЕЭГЕФЮЈвЛЭООЖЪЧЙтКЯзїгУЃЈМЋаЁВПЗжЮЊЛЏФмКЯГЩЃЉЁЃТЬЩЋжВЮяЕШЩњВњепНЋЬЋбєФмзЊЛЏЮЊЛЏбЇФмЃЌДЂДцдкгаЛњЮяжаЃЌЦфЫйТЪГЦЮЊГѕМЖЩњВњСІЁЃетЪЧећИіЩњЬЌЯЕЭГФмСПЕФзмРДдДЁЃ
2. ФмСПСїЕФаЇТЪгыЕнМѕЃКСжЕТТќаЇТЪ
ФмСПдкДгвЛИігЊбјМЖДЋЕнЕНЯТвЛИігЊбјМЖЃЈШчДгВнЕНбђЃЌДгбђЕНРЧЃЉЪБЃЌОјДѓВПЗжЛсвдШШЕФаЮЪНЩЂЪЇЃЌетЪЧгЩКєЮќзїгУЁЂЮДРћгУВПЗжКЭХХаЙЮядьГЩЕФЁЃЦНОљДЋЕнаЇТЪжЛгадМ10%ЃЌМДжјУћЕФЁАЪЎЗжжЎвЛЖЈТЩЁБЃЈБэ11ЃЉЁЃ
Бэ11 СжЕТТќаЇТЪЕФФмСПСїДЋЕнЙцТЩ
гЊбјМЖ | ФмСПРДдД | ФмСПШЅЯђЃЈвдЩуШы100%ФмСПМЦЃЉ | ОЛЩњВњСПЃЈдМжЕЃЉ |
ЩњВњеп | ЬЋбєФм | КєЮќКФЩЂ (50-60%) + ЮДРћгУ/ЗжНтепРћгУ + ОЛГѕМЖЩњВњСП | 40-50% |
ГѕМЖЯћЗбеп | жВЮя | КєЮќКФЩЂЁЂХХаЙЁЂЮДЪГгУ + ЭЌЛЏСП | дМ10% |
ДЮМЖЯћЗбеп | ЪГВнЖЏЮя | ЭЌЩЯЃЌаЇТЪИќЕЭ | дМ1% |
ЖЅМЖЯћЗбеп | ЯТМЖЯћЗбеп | ФмСПвбМЋЖШЯЁБЁЃЌФбвджЇГжИќИпгЊбјМЖ | МЋЩй |
ЩњЬЌН№зжЫўЃЈЪ§СПЁЂЩњЮяСПЁЂФмСПН№зжЫўЃЉе§ЪЧетвЛЕнМѕЙцТЩЕФжБЙлЬхЯжЃЌЦфжаФмСПН№зжЫўгРдЖБЃГже§Н№зжЫўаЮЁЃ
3. ФмСПСїЕФжеЕуЃКЗжНтгыКФЩЂ
ЫљгагЊбјМЖВњЩњЕФПнжІТфвЖЁЂЪЌЬхЁЂХХаЙЮязюжеБЛЗжНтепЃЈЯИОњЁЂецОњЃЉРћгУЁЃЗжНтепЭЈЙ§КєЮќзїгУНЋЦфЛЏбЇФмГЙЕзЪЭЗХЮЊШШФмЃЌЭъГЩФмСПСїЕФзюжеЛЗНкЃЌЭЌЪБНЋЮоЛњЮяЙщЛЙЛЗОГЃЌЭъГЩЮяжЪбЛЗЁЃ
4. ЫМЯыЪЗЕФжеМЋЭГвЛЃКДгжЪзгЬнЖШЕНЩњЮяШІ
дкФњЫљзЗЫнЕФДгЁАЛюСІТлЁБЕНЁАЩњЮяФмбЇЁБЕФЫМЯыЪЗжаЃЌЩњЬЌЯЕЭГФмСПСїЬсЙЉСЫзюжеГпЖШЕФЭГвЛаджЄУїЃК
ШШСІбЇЦеЪЪадЕФЭъећЬхЯжЃКЫќжЄУїЃЌЮоТлЪЧATPКЯГЩЁЂЯИАћКєЮќЃЌЛЙЪЧЪЈШКВЖСдЁЂЩСжЩњГЄЃЌЖМбЯИёзёЪиФмСПЪиКугыьидіЖЈТЩЁЃЩњЬЌЯЕЭГЪЧвЛИіОоДѓЕФПЊЗХКФЩЂНсЙЙЁЃ
ФмСПЛѕБвЕФЭГвЛЃКЧ§ЖЏЩњЬЌЯЕЭГдЫзЊЕФЁАЭЈЛѕЁБЃЌЙщИљНсЕзЪЧжВЮяЭЈЙ§ЙтКЯзїгУЗтзАдкЦЯЬбЬЧЃЈМАЦфбмЩњгаЛњЮяЃЉжаЕФЛЏбЇФмЃЌетгыЯпСЃЬхжабѕЛЏСзЫсЛЏЕФЁАШМСЯЁББОжЪЯрЭЌЁЃ
ЖдЛюСІТлЕФГЙЕзГЌдНЃКЩњЬЌЯЕЭГЕФКъЮАЁЂИДдггыПДЫЦОЋУюЕФЦНКтЃЌЭъШЋВЛашвЊЁАИЧбЧМйЫЕЁБЪНЕФећЬхЩњУќСІЁЃЫќдДгкЮоЪ§ИіЬхзёбЮяРэЛЏбЇЖЈТЩНјааФмСПзЊЛЛгыОКељКЯзїКѓЃЌздЯТЖјЩЯгПЯжГіЕФКъЙлФЃЪНЁЃ
СЌНгЮЂЙлгыКъЙлЕФЧХСКЃКФњжЎЧАЬНЬжЕФЁАЮжВЎИёаЇгІЁБЁЂЁАЯпСЃЬхЙІФмеЯАЁБЕШЃЌдкЩњЬЌГпЖШЩЯЖдгІзХФмСПзЊЛЏаЇТЪЕФИФБфЃЌжБНггАЯьИіЬхЪЪКЯЖШЃЌНјЖјЭЈЙ§здШЛбЁдёЫмдьжжШККЭШКТфНсЙЙЁЃ
НсТлЃКЩњЬЌЯЕЭГФмСПСїЃЌЪЧНЋЩњЮяФмбЇдРэЪщаДдкЕиЧђГпЖШЩЯЕФзГРіЦЊеТЁЃЫќНвЪОЃЌЩњУќЪРНчЕФЗБЛЊЁЂжШађгыбнЛЏЃЌЦфзюЕзВуЕФЧ§ЖЏСІЃЌЮоЗЧЪЧЬЋбєФмзЊЛЏЮЊЛЏбЇФмЃЌВЂбиЪГЮяСДдкКФЩЂжаж№МЖДЋЕнЕФЮяРэЙ§ГЬЁЃетЮЊЮвУЧТўГЄЕФЫМЯыЬНЫїЛЩЯСЫвЛИіКъДѓЁЂЭГвЛЧвМсЪЕЕФОфКХЃКЩњУќМДЪЧФмСПСїОЮяжЪЪБЫљГЪЯжЕФЃЌЖЬднЖјгаађЕФаЮЬЌЁЃ
ОХЁЂДгЙтКЯзїгУЕНЪГЮяСДЃКФмСПдкЩњЮяШІжаЕФДЋЕнгыКФЩЂ
ЃЈЮДЭъД§ајЃЉ
https://blog.sciencenet.cn/blog-279293-1515534.html
ЩЯвЛЦЊЃКЩњЮяФмРэТлЕФгІгУЧАОАЃЈ3ЃЉ
ЯТвЛЦЊЃКЩњЮяФмРэТлЕФгІгУЧАОАЃЈ5ЃЉ
