博文
潜水动物直接感受低氧避免溺水  精选
精选
||
潜水动物能感受氧浓度
氧气对需氧生物极端重要,不可能缺少。几乎所有其他哺乳动物对血氧水平都不敏感,而是通过感知不断增加的二氧化碳来衡量血氧水平。我个人推测,可能是在进化过程中,由于低氧具有强致命性,往往无法给感受这种致命危害提供进化加强保留机会。导致大部分动物都无法从认知层面感受到不同水平的氧气不足。例如我们人类到高原上,只会感到心慌呼吸加快,但我们无法直接感受到低氧。这不同于低温和高温等伤害刺激,能让我们正常人都刻骨铭心地感受到。
编辑总结
海洋哺乳动物适应了水下环境,尽管需要呼吸空气,但它们仍会长时间在水下觅食。因此,它们特别容易受到低血氧水平的影响,这可能会在潜水时导致溺水。麦克奈特等人让潜水的灰海豹接触各种气体混合物,发现海豹对低血氧水平有反应,而对高二氧化碳水平无反应。几乎所有其他哺乳动物对血氧水平都不敏感,而是通过感知不断增加的二氧化碳来衡量血氧水平。直接感知氧气水平可能会提高潜水效率。——萨沙·维尼耶里

摘要
海洋哺乳动物在潜水时依赖维持足够的血氧水平来防止溺水。一般来说,哺乳动物无法认知感知氧气,而是将感知上升的二氧化碳作为低氧的替代指标。然而,对潜水哺乳动物而言,无法感知氧气存在风险。我们认为,任何基于直接感知氧气来改变潜水行为的能力都应受到强烈的选择。我们让潜水的海豹吸入经过实验调整的气体混合物,以影响其循环中的氧气和二氧化碳水平。潜水时长与循环氧气水平呈正相关,但不受二氧化碳水平和pH值的影响。这些结果表明,海豹确实能够认知感知循环中的氧气,并利用这一感知来改变潜水行为。
属于三个呼吸空气动物纲(爬行纲、鸟纲和哺乳纲)的众多物种会进行屏气潜水,且在潜水过程中不与周围环境气体进行额外的呼吸交换。在这些分类群中,存在着一系列不同的潜水方式,从短暂、偶尔的间歇性潜水到终生重复潜水。在非洲兽总目、偶蹄目和食肉目海洋哺乳动物中,进化出了一系列趋同的生理适应特征。这些适应特征包括诸如体温调节、水生推进和流体动力学、承受静水压力以及有效的远程感知等压力(1)。然而,对于潜水哺乳动物面临的最严峻选择压力——不溺水,目前尚不清楚存在哪些进化适应机制。在此,我们研究了海洋哺乳动物是如何避免溺水的。
即使是海洋哺乳动物,尽管其具有较大的内源性氧气储存能力(1)和对低脑氧的高耐受性(2, 3),但如果无法认知感知循环中的氧气何时耗尽并相应调整潜水行为,仍然面临着很高的溺水风险。在此,我们使用 “认知感知” 这一术语来描述通过化学感受器产生的传入信号经神经通路到达大脑中枢的过程,这些神经通路使化学感觉信息能够被整合到影响行为的决策中(4) [图1(5)]。我们将大脑决策过程与自主化学感觉髓质介导的反射(如呼吸和心脏反射)区分开来,因为后者在去大脑动物中依然存在(6)。哺乳动物通常无法认知感知环境和内源性氧气水平的变化(7 - 9),在某些情况下,只有在氧气水平极低、意识发生改变时或即将改变时才能感知到(5, 10)。相反,哺乳动物几乎普遍进化出了将认知感知二氧化碳作为低氧水平替代指标的能力(图1A)。哺乳动物将血液中二氧化碳水平升高(11, 12)感知为 “空气饥饿”、不适和呼吸困难等厌恶感(13, 14)。对升高的二氧化碳的认知感知使哺乳动物能够间接感知环境氧气水平中可能导致缺氧的潜在有害变化。

图1. 氧气和二氧化碳认知感知的神经通路
(A)人类中引起空气饥饿感认知表现的神经通路,箭头越粗,表示对高二氧化碳的相对感知度高于低氧气。该通路由班泽特等人报告(5)。(B)海豹中假设的对低氧气相对感知度高于高二氧化碳的通路。
哺乳动物对升高的二氧化碳的认知感知被强烈选择,这表明它是一种至关重要的生存特征。然而,在潜水的情境下,其有效性可能有限。人类屏气潜水时,只感知二氧化碳而不感知氧气的缺陷十分明显。人类自由潜水者在潜水前过度换气会降低循环中的二氧化碳水平,进而可能导致意识丧失和溺水(15)。这种微妙的生理变化会延迟由二氧化碳刺激的呼吸驱动点,延长潜水时间,从而使缺氧情况发展并导致溺水(15)。因此,仅靠对二氧化碳的认知感知不足以保护潜水哺乳动物免受低氧导致溺水的风险。
有间接证据表明,海洋哺乳动物在潜水时可能具有认知感知氧气的能力。对港海豹(斑海豹)(16)和亚马逊海牛(17)呼吸控制的研究报告称,与环境气体相比,暴露于低氧气体后,它们的潜水时间缩短。然而,这些研究并未探究认知和氧气感知的作用。此外,海豹表现出显著的解剖学适应性,这可能表明其存在更敏感的循环氧气感知装置。海豹的颈动脉体能检测循环中氧气的变化,与陆地哺乳动物相比,其拥有更多的自主神经节细胞(18),且1型细胞数量增加与低氧暴露有关(19)。海豹经常暴露于慢性间歇性低氧环境中,这也被证明能选择性地增强颈动脉体对低氧的感觉反应(20)。海豹还能在不同的觉醒状态(即清醒与睡眠)下保持化学感受器的敏感性(21)。
考虑到这些间接证据以及海洋哺乳动物在潜水时不会溺水的事实,我们对 “海豹等海洋哺乳动物对氧气认知不敏感” 这一零假设提出了质疑(图1A)。相反,我们提出,海洋哺乳动物的生态环境决定了二氧化碳作为潜水行为关键认知驱动因素的作用减弱,而对氧气认知感知的增强进化(图1B)定义了这一群体在生态上的成功以及终生潜水的转变。
实验方案:模拟觅食设置
为了验证上述假设,我们通过实验操纵吸入的氧气和二氧化碳水平,以影响肺泡气体浓度,从而改变海洋哺乳动物灰海豹(Halichoerus grypus)循环血液中的氧气和二氧化碳浓度(22 - 24)。我们研究了吸入氧气和二氧化碳浓度的变化对潜水行为、代谢率和静脉pH值的影响。六只野生捕获的幼年灰海豹在短期圈养条件下,在模拟觅食环境中进行重复潜水(图2A)。它们从呼吸室游到水下60米的水下喂食站,然后返回呼吸室。只有在呼吸室才能接触到空气(25)。觅食环境中的所有行为都是自发和自愿的,海豹可以自由选择潜水的频率和时长、水面呼吸间隔、游泳速度以及在喂食站停留的时间。一次潜水试验由海豹发起的一系列潜水组成,当海豹消耗完1.5千克鲱鱼(Sprattus sprattus)的固定定量后,试验结束。
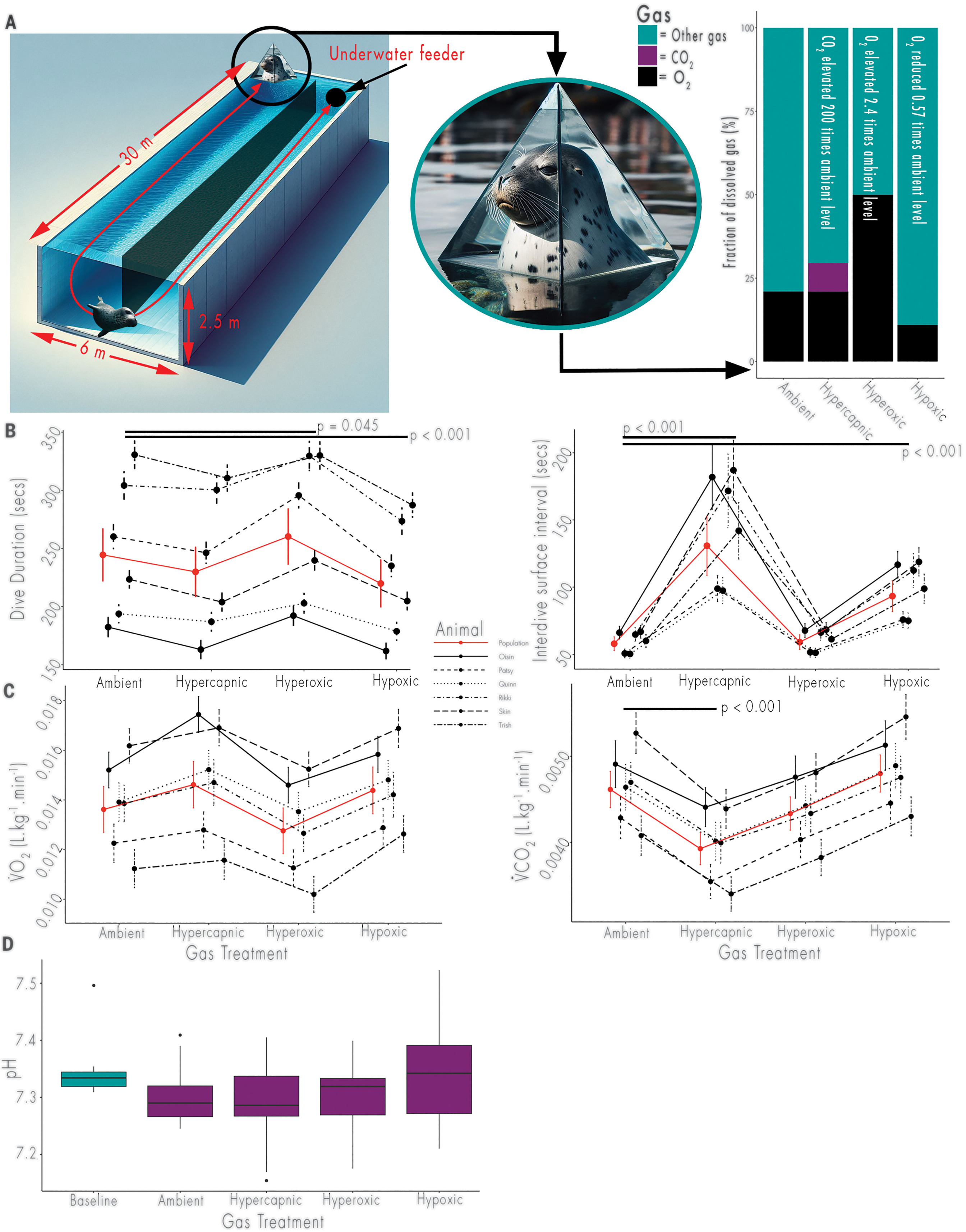
图2. 实验设置及对气体处理的行为和生理反应可视化
(A)模拟觅食范式(左)和呼吸室(中),海豹在呼吸室浮出水面并吸入四种处理气体之一(右)。(B)每种气体处理下潜水时长(左)和潜水间隔(右)的平均预测反应。符号表示个体平均反应,误差线表示标准误差。(C)每种气体处理下氧气消耗率(左)和二氧化碳产生率(右)的平均预测反应。(D)每种气体处理下基线(超过24小时未潜水时的水平)和潜水后不到2分钟的pH值。
试验期间,呼吸室中吸入气体的浓度(氧气百分比:二氧化碳百分比)设置为四种气体混合物处理之一:环境气体(21%氧气:0.04%二氧化碳)、高碳酸血症气体(21%氧气:8%二氧化碳)、高氧气体(50%氧气:0.04%二氧化碳)或低氧气体(11%氧气:0.04%二氧化碳)(图2A)。其余气体由氮气和其他自然大气气体的痕量元素组成。较低的氧气水平是根据对自愿潜水的幼年港海豹呼吸的研究选定的,研究表明11%的氧气(环境水平的0.52倍)不会影响呼吸频率,对潮气量也只有轻微影响(16)。较高的氧气水平(环境水平的2.38倍)和二氧化碳水平(环境水平的200倍)是气体控制系统所能达到的最高水平。在潜水开始前,海豹先接触每种气体混合物5分钟。总共进行了119次潜水试验,包括510次单独潜水(表S1)。通过每次试验的视频记录每次潜水的潜水时长和潜水间隔。潜水间隔仅包括潜水之间的水面间隔,因此试验中最后一次潜水后的恢复时间不计算在内(n = 405)。分别通过开放流呼吸测定法收集了107次和100次潜水试验的氧气消耗率(V̇O₂)和二氧化碳产生率(V̇CO₂)。通过远程动物携带的血液采样器采集静脉血样来测量pH值(图S1),采样时无需处理动物、使用镇静剂或麻醉剂。在之前一次潜水试验超过24小时后,使用该设备远程采集基线血样(n = 12)。在潜水试验完成后2分钟内采集潜水血样(n = 51)。每种动物和气体处理收集的数据总结在表S1中。使用广义相加混合效应模型(GAMM)框架分别对气体处理对潜水时长、潜水间隔、V̇O₂和V̇CO₂的影响进行建模(表S2)。使用Tukey检验比较气体处理对游泳时间、潜水后和基线状态下血液pH值的影响。
氧气和二氧化碳对潜水和浮出水面行为的影响
潜水时长
与环境条件下的潜水时长(平均246 ± 22秒)相比,增加氧气供应(高氧潜水)使潜水时长增加了6.4%(平均260 ± 24秒;P = 0.045)。相反,减少氧气供应(低氧潜水)使潜水时长缩短了10.0%(平均219 ± 20秒;P < 0.001)。升高的二氧化碳(高碳酸血症潜水)对潜水时长没有显著影响(229 ± 21秒;P = 0.06)。由于气体处理对往返喂食站的游泳时间没有影响(P分别为0.93和0.73),潜水时长的变化是由在喂食站静止停留的时间驱动的。
潜水间隔时长
潜水间隔受到较低氧气供应和较高二氧化碳的影响(图2B),这表明它受到氧气和二氧化碳暴露的共同影响。与暴露于环境空气下的潜水间隔(58 ± 5秒)相比,高碳酸血症处理时潜水间隔显著增加了125.8%(131 ± 22秒;P < 0.001),低氧处理时增加了60.3%(93 ± 12秒;P < 0.001)。高氧潜水(59 ± 5秒;P = 0.8)与环境潜水相比,对潜水间隔没有显著影响(表S3)。高氧对潜水间隔没有显著影响,这可能是由于二氧化碳排出受限(26),而非氧气加载动态变化所致。
氧气消耗和二氧化碳产生
气体处理对单位质量氧气消耗率(以每分钟每千克体重消耗的氧气升数衡量)没有显著影响(P > 0.05)(图2C)。它对单位质量二氧化碳产生率有显著影响(P = 0.001),但仅在高碳酸血症处理下(图2C)。与环境潜水相比,高碳酸血症使V̇CO₂(0.037 ± 0.0001升/千克·分钟)降低了17.8%(0.045 ± 0.0002升/千克·分钟)(表S3)。然而,高碳酸血症对V̇CO₂的影响可能是由于肺血和肺泡之间缺乏排出循环二氧化碳所需的适当扩散梯度而导致的二氧化碳潴留假象,而非代谢率的变化。
pH值
气体处理对静脉血pH值没有影响(P = 0.14)(图2D和表S4)。
潜水行为与氧气感知
海豹会根据吸入氧气的变化改变潜水行为,但对二氧化碳的变化无反应。与在环境空气中潜水相比,低氧潜水时潜水时长缩短,高氧潜水时潜水时长延长。这种对潜水时长的行为调节表明,海豹实际上是通过对循环氧气的认知感知来调节潜水,而非之前报道的哺乳动物通过认知感知二氧化碳的途径(图1A)。此外,在其他哺乳动物中,pH值产生的传入信号会被认知整合以引发行为反应(27),但在本研究中,pH值既不影响潜水时长(图2B),在不同气体处理组间也不存在统计学差异(图2D)。高碳酸血症条件下吸入的二氧化碳水平为60.8 mmHg,是威德尔海豹自愿潜水结束时测量的肺泡二氧化碳分压的1.5倍(28)。因此,尽管二氧化碳水平超过了海豹正常接触的水平,但影响潜水时长的是氧气。这使我们能够否定“海豹等海洋哺乳动物对氧气认知不敏感”这一零假设。相反,海豹将对氧气的感知作为调节潜水的主要认知输入,而非二氧化碳(图1B)。与小鼠、大鼠和貂等物种表现出的简单厌恶反应(29 - 31),或是仅在氧气分压极低、意识发生改变时才出现的反应不同,我们推测,对氧气的认知感知是海洋哺乳动物用于在潜水过程中及潜水后做出行为决策的信息反馈系统的一部分。具体而言,海豹能够通过行为调节潜水,避免动脉血氧水平降至会导致严重神经功能障碍(即缺氧性晕厥)和溺水的程度。
氧气感知的潜在机制
我们推断,海豹感知循环氧气的能力与已知的颈动脉体氧气化学感受器的自主功能有关。这些感受器在海豹的低氧性过度通气(22, 32, 33)(图1)、屏气潜水时维持心动过缓(32, 34)以及代谢率变化(35)中发挥着关键的氧气介导的自主调节作用。虽然海豹颈动脉体对循环氧气变化产生的传入信号会引发自主性反射传出反应这一点已得到充分证实,但在此我们提供证据表明,这些传入信号也会被认知整合从而影响行为。
我们对哺乳动物(包括人类)颈动脉二氧化碳化学感受器信息的认知整合已有深入理解。血液中二氧化碳水平升高在人类身上表现为空气饥饿感(7)和呼吸急促(36)等感觉。相反,在缺氧期间,通气反应增加会导致机械感受器放电增强,从而抑制空气饥饿感(5, 7)。这些现象背后的生理机制被认为与小脑(14)和杏仁核(13)有关(图1)。此外,杏仁核已被证明是小鼠对二氧化碳升高产生恐惧反应的主要部位,这种反应是通过基于二氧化碳的血液pH值酸化介导的(27)。同样,人类在二氧化碳升高时会出现类似恐慌的症状(13, 14),并且有人假设包括小胶质细胞在内的其他化学感觉系统参与了对高碳酸血症的检测,并将其转化为与恐惧相关的行为反应(37)。在本研究中,二氧化碳升高对潜水时长没有显著影响,海豹对8%的二氧化碳暴露也没有表现出任何厌恶迹象,这表明其对二氧化碳的认知敏感性有所降低。此外,我们发现海豹在水面间隔期间有明显的二氧化碳潴留现象(图2C),这可能意味着诸如恐惧和不适等体验背后的生理机制可能有所不同、有所减弱,甚至可能被pH缓冲作用抵消(图2D)。二氧化碳显然会影响海豹浮出水面的行为,因为高碳酸血症使潜水间隔显著延长(图2B),且影响程度比低氧更大,这可能与二氧化碳排出受限有关(26)。然而,海豹对二氧化碳的反应似乎与高度适应地下生活的哺乳动物(如裸鼹鼠[Fukomys damarensis(38),Spalax ehrenbergi(39)])类似。鉴于海洋哺乳动物由于二氧化碳交换受限(26),在连续潜水过程中二氧化碳负担会不断增加,且在本研究中海豹在高二氧化碳负担下仍继续潜水,因此依赖对二氧化碳的感知来替代对氧气的感知显然并不可靠。还需要进一步研究来确定潜水过程中对二氧化碳感知的减弱是在脑干区域(如后梯形核)的中枢层面产生,还是通过短暂抑制交感神经活动降低颈动脉体对二氧化碳的敏感性而在周边层面产生(41)。
基于氧气感知的决策
我们的研究结果表明,海豹似乎会基于对循环氧气的感知做出决策。这种决策可以在水面上做出,也可以在潜水过程中做出。然而,根据我们对通过颈动脉体感知循环氧气的理论,这些行为状态并非相互排斥。目前尚不清楚在潜水和/或浮出水面的过程中,这一信息何时会影响行为决策。由于较低的氧气可用性会缩短潜水时长并延长潜水间隔,我们认为认知整合在潜水和浮出水面阶段均发挥作用,具体讨论如下。
有观点认为海豹可能会在水面上“规划”潜水时长并相应调整代谢率(33, 42, 43),这支持了海豹在水面上具有认知感知氧气水平的能力。以心率作为氧气消耗率的近似指标,自由放养海豹在潜水时间较长时心动过缓加剧,这表明氧气消耗率与潜水时长呈负相关(44)。与这一观点相符的是,潜水时长与氧气可用性呈正相关,总体而言,在最长的潜水中氧气消耗率最低(图2B和图2C)。
在潜水过程中,溺水风险最大,这支持了海豹在潜水时能够感知氧气水平的观点。总体而言,尽管高碳酸血症潜水时氧气可用性相同,但氧气消耗率会增加,这会更快地降低循环氧气水平。因此,与低氧和高氧潜水的反应一致,氧气可用性也可能导致高碳酸血症潜水时潜水时长缩短,但在5%的显著性水平下,这一反应并不显著(P = 0.06)。
氧气感知的进化视角
在本研究中,我们证实了单一海洋哺乳动物物种具有认知感知氧气的能力。鉴于多种表型在重复潜水的共同选择压力下发生趋同进化(1),我们假设类似的通过感知氧气来调节潜水行为的现象可能在海洋哺乳动物中普遍存在。港海豹(16)和亚马逊海牛(17)的相关间接证据支持了这一假设。此外,一项关于高碳酸血症对斯特勒海狮(Eumetopias jubatus)呼吸控制和代谢影响的研究表明,二氧化碳升高不会影响其潜水时长(45),这支持了二氧化碳并非海洋哺乳动物潜水行为关键认知驱动因素的观点。为了确定在动物界中,认知感知氧气是否在更大范围内因潜水和间歇性缺氧而趋同进化,必须针对其他潜水动物类群(如鸟类和爬行类)开展有针对性的研究。关于氧气可用性对潜水物种代谢、呼吸和心血管功能影响的数据确实存在,但此前没有研究像我们对海豹所做的研究一样,实证探究认知感知氧气水平在潜水调节中的作用。对凤头潜鸭(Aythya fuligula)(46)、小潜鸭(Aythya affinis)(47)、滑龟(Pseudemys scripta)(48)、软壳龟(Trionyx spiniferus)(49)、鳄龟(Chelydra serpentina)(50)、枯叶龟(Chelus frimbriata)(51)和尼罗鳄(Crocodylus niloticus)(52)等物种的研究结果与海豹的行为反应极为相似。这可能表明认知感知氧气的能力在更广泛的范围内进化,是屏气潜水动物抵御缺氧溺水风险的关键适应性特征。
https://blog.sciencenet.cn/blog-41174-1478993.html
上一篇:饮用氢气纳米气泡水辐射防护作用研究【日本2025】
下一篇:富氢水对拥挤应激的保护作用
