博文
42亿年前以氢气为食的生命始祖
||
以氢气为食的地球生命始祖
关于最后通用共同祖先(LUCA)的性质、年龄及其对地球系统的影响,一直是跨学科领域激烈辩论的主题,这些讨论往往基于不同的数据和方法。LUCA的年龄估计通常基于化石记录,随着每一次重新解释而变化。LUCA的新陈代谢性质也同样备受争议,一些人将所有核心代谢归因于LUCA,而另一些人则重建了一个依赖地质化学的更简单生命形式。在这里,我们通过前LUCA基因复制品的分歧时间分析推断出LUCA生活在大约42亿年前(40.9-43.3亿年前),这一分析利用了微生物化石和同位素记录在一种新的交叉支撑实现下进行了校准。系统发育调和表明,LUCA的基因组至少有2.5 Mb,编码约2600种蛋白质,与现代原核生物相当。我们的结果暗示LUCA是一个具有早期免疫系统的原核级厌氧产乙酸菌。尽管有时认为LUCA是孤立生活的,但我们推断LUCA是一个已建立的生态系统的一部分。LUCA的新陈代谢为其他微生物群落成员提供了生态位,大气光化学中的氢循环可能支撑了一个适度生产力的早期生态系统。
Moody, E.R.R., Álvarez-Carretero, S., Mahendrarajah, T.A. et al. The nature of the last universal common ancestor and its impact on the early Earth system. Nat Ecol Evol (2024).
所有现存细胞生命的共同祖先由通用遗传密码、蛋白质合成机制、几乎普遍存在的20种氨基酸的共同手性以及ATP作为通用能量货币的使用所证明。最后通用共同祖先(LUCA)是生命树上的一个节点,从该节点分出了基本的原核域(古细菌和细菌)。因此,我们对LUCA的理解影响了我们对地球生命早期进化的理解。LUCA是一个简单的还是有复杂结构的生物?它生活在什么样的环境中,又是何时?之前对LUCA的估计存在冲突,要么是因为对LUCA是什么的概念性分歧,要么是由于不同的方法学方法和数据导致的。已发布的分析在推断LUCA的基因组时有所不同,从保守估计的80个直系同源蛋白到多达1529个不同潜在基因家族不等。解释范围从小至信息处理和代谢核心,到拥有现代古细菌和细菌大部分基因库的原核级生物不等,最近在参考文献7中进行了回顾。在这里,我们使用分子钟方法、意识到水平基因转移的系统发育调和以及现有的生物地球化学模型来解答有关LUCA的年龄、基因内容、新陈代谢及其对早期地球系统影响的问题。
一、估计LUCA的年龄
生命的进化时间尺度通常以最古老的化石出现为标准。然而,对早期太古宙时期化石发现的可信度一直存在争议。宽松的贝叶斯节点校准分子钟方法提供了一种手段,将早期生命的稀疏化石和地球化学记录与分子数据提供的信息相结合;然而,由于原核生物化石校准有限以及它们在系统发生树上位置的不确定性,确定LUCA的年龄具有挑战性。分子钟对LUCA的估计依赖于系统发生学中保守的通用单拷贝标记基因,其中LUCA代表根。确定一棵树的根部是困难的,因为错误从树的末端传播到被测系统发生学的根部,并且没有可用的信息来估计与根节点相连的分支的进化速率。因此,我们分析了在LUCA之前复制的基因,这些基因在LUCA的基因组中有两份(或更多)副本。在这些基因树中,根代表了这次复制发生在LUCA之前,而LUCA由两个后代节点表示。使用这些通用旁系同源基因还有一个优势,即相同的校准至少可以应用两次。复制后,相同的物种分化在基因树的两侧都有所体现,因此可以假设具有相同的年龄。这极大地减少了当遗传距离(分支长度)被解析为绝对时间和速率时的不确定性。当一个共享节点被分配了一个化石校准时,这种交叉支撑也有助于将校准数量加倍,从而提高分歧时间估计的准确性。我们使用13个校准对我们的分子钟分析进行了校准(见补充信息中的“化石校准”)。生命之树根部的校准尤为重要。一些先前的研究基于生命无法在晚期重轰炸期(LHB)(约37-39亿年前)存活下来的假设,对LUCA的年龄设定了一个较年轻的最大约束。然而,LHB假说是从月球撞击记录推断和扩展的,其强度、持续时间甚至LHB事件的真实性的解释已受到质疑。因此,LHB假说不应被视为对LUCA年龄的一个可信的最大约束。我们使用了软均匀边界,最大年龄边界基于形成月球的撞击时间(45.10亿年±10百万年),这将有效地使地球的前身Tellus和Theia灭绝。我们对LUCA年龄的最小边界基于低δ98Mo同位素值,指示与氧化锰相容的光合作用,因此,南非Pongola超群Mozaan组中的全群Oxyphotobacteria最早可追溯至29.54亿年。
我们对LUCA年龄的估计是通过串联和分区数据集推断的,两者都包括五个前LUCA旁系同源基因:ATP合成酶的催化和非催化亚基、延伸因子Tu和G、信号识别蛋白和信号识别颗粒受体、酪氨酰-tRNA和色氨酰-tRNA合成酶,以及亮氨酰和缬氨酰-tRNA合成酶。边际密度(通常称为有效先验)在拓扑相邻的校准不重叠时落在校准密度内(即用户指定的先验),但当它们重叠时可能会有所不同,以确保祖先-后代节点之间的相对年龄关系。我们认为边际密度是对给定系统发生学的校准证据的合理解释;我们并不是试图测试化石记录是进化历史的准确时间档案这一假设,因为它不是。我们在自相关速率(几何布朗运动(GBM))和独立速率对数正态(ILN)放松时钟模型下,使用我们的分区数据集获得的复制LUCA节点年龄估计(GBM, 41.8-43.3亿年; ILN, 40.9-43.2亿年;图1)落在我们对LUCA的综合年龄估计范围内,从39.4亿年到45.2亿年,与先前的研究相当。基于单个基因或依次排除每个基因的串联进行的时间分析返回了兼容的时间尺度(扩展数据图1和2及方法中的“附加方法”)。
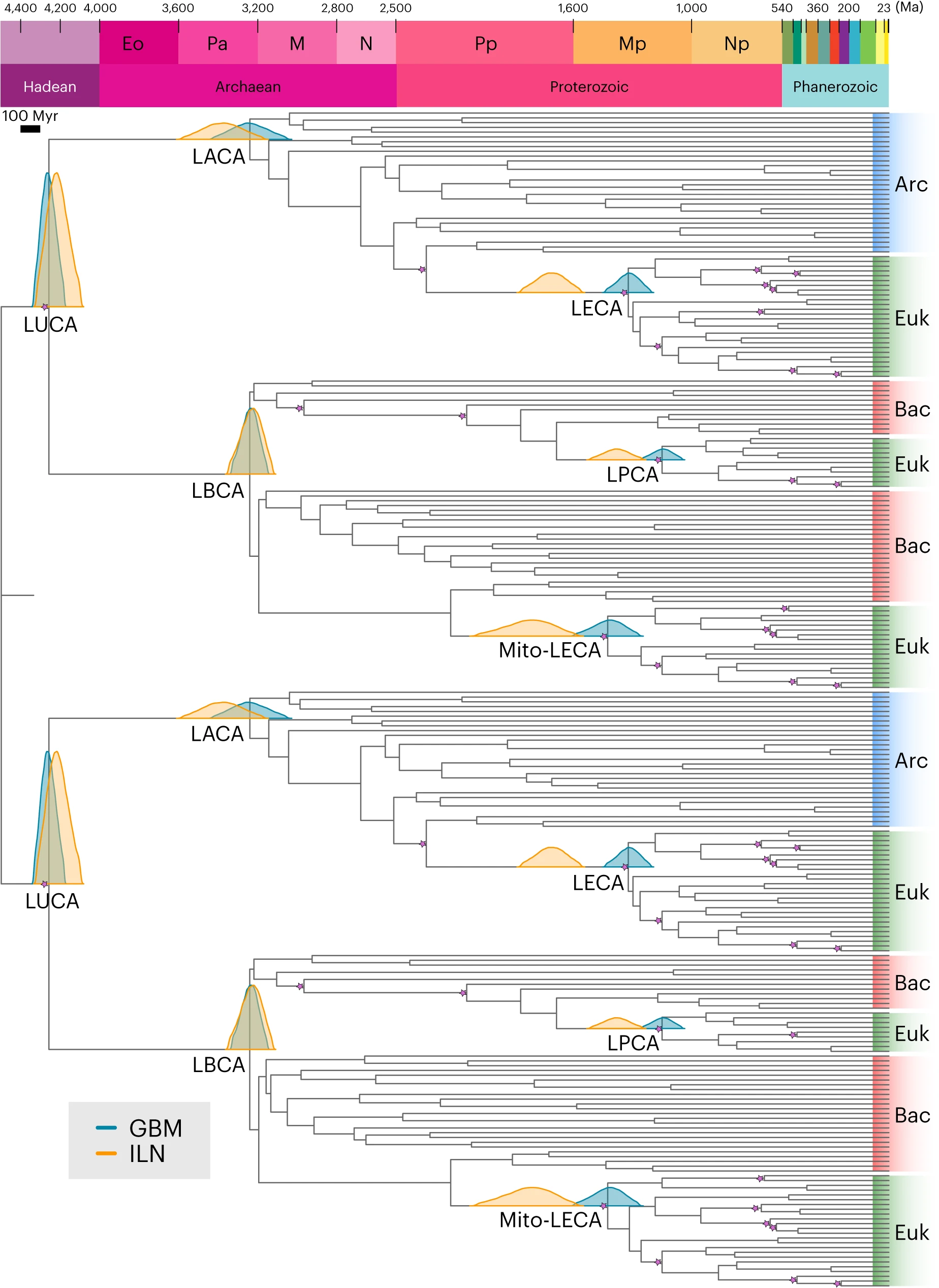
图1:使用贝叶斯节点定年法和交叉支撑方法,通过五个前LUCA旁系同源基因的分区数据集推断出的时间树。
我们的结果暗示LUCA生活在约42亿年前,95%置信区间在ILN放松时钟模型下为4.09-4.33Ga(橙色),在GBM放松时钟模型下为4.18-4.33Ga(蓝绿色)。在交叉支撑方法下,对应于同一物种分化的节点(即镜像节点)具有相同的后验时间密度。该图显示了最后通用祖先、古细菌、细菌和真核生物共同祖先(分别为LUCA、LACA、LBCA和LECA)、线粒体谱系的最后一位共同祖先(Mito-LECA)以及最后一位带有质体的共祖(LPCA)的镜像节点对应的后验时间密度;紫色星星表示用化石校准的节点。Arc, 古菌;Bac, 细菌;Euk, 真核生物。
二、LUCA的生理学
为了估计LUCA的生理学,我们首先从57个系统发育标记基因(参见方法中的“通用标记基因”)对700个基因组进行了更新的微生物系统发育推断,包括350个古细菌和350个细菌。这棵树与最近关于古菌和细菌生命域的系统发育非常一致。例如,TACK和阿斯加德群的古菌以及细菌中的纤细杆菌都被恢复为单系群。然而,对于Patescibacteria(CPR)和DPANN这两个难以在树中定位的小基因组谱系,分析结果并不明确。大约无偏44测试无法区分这些类群的位置,既不能确定它们各自域的根部位置,也不能确定衍生位置,CPR作为绿弯菌门的姐妹群和DPANN作为广古菌门的姐妹群。为了考虑这种系统发育不确定性,我们在两棵树上进行了LUCA重建:我们的最大似然(ML)树(拓扑结构1;扩展数据图3)和一棵树,其中CPR被放置在绿弯菌门的姐妹群位置,DPANN被放置在所有其他古菌的姐妹群位置(拓扑结构2;扩展数据图4)。在这两种情况下,映射到LUCA的基因家族非常相似(LUCA存在概率(PP)的相关性,r=0.6720275, P<2.2×10−16)。我们在拓扑结构2的树上讨论结果,并在补充信息中的“拓扑结构1”中讨论剩余的差异。
我们使用概率性基因和物种树调和算法ALE46来推断KEGG直系同源数据库(KO)47中每个样本条目的基因家族树的演化过程,基于我们的物种树。ALE根据一系列自展基因树与参考物种树之间的比较来推断基因复制、转移和丢失的历史,使我们能够估计基因家族在树的一个节点处存在的概率。这种调和方法对于推断LUCA有若干优势。大多数基因家族自LUCA时代以来经历了基因转移,因此显式建模转移使我们能够在分析中包含比之前方法可能包含的更多的基因家族。由于分析是概率性的,我们还可以通过平均不同的场景使用调和模型来考虑基因家族起源和演化历史的不确定性。使用这种方法,我们估计了每个KEGG基因家族(KO)存在于LUCA中的概率,然后使用得到的概率构建了LUCA基因内容、代谢潜力(图2)和环境背景(图3)的假设模型。使用KEGG注释是有益的,因为它允许我们将推断与经过策划的功能注释联系起来;然而,它的一个缺点是一些可能在LUCA中普遍存在的广泛基因家族被分成多个KO家族,这些家族个别看起来仅限于特定分类群并被推断为后来出现。为了解决这一限制,我们还对COG(直系同源基因簇)基因家族进行了分析,这些基因家族对应于更粗略的功能注释(补充数据2)。

图2: 现代生活中存在的LUCA代谢网络的概率估计。
黑色:推断存在于LUCA中的酶和代谢途径,至少PP(后验概率)=0.75,在两个原核生物域中都有抽样。灰色:在我们最不严格的阈值PP=0.50下推断出的。分析支持存在一个完整的WLP(伍德-隆达尔途径)和一个几乎完整的TCA(三羧酸循环)跨越多个置信阈值。代谢图谱来源于KEGG数据库通过iPath工具。GPI,糖基磷脂酰肌醇;DDT,1,1,1-三氯-2,2-双(对氯苯基)乙烷。
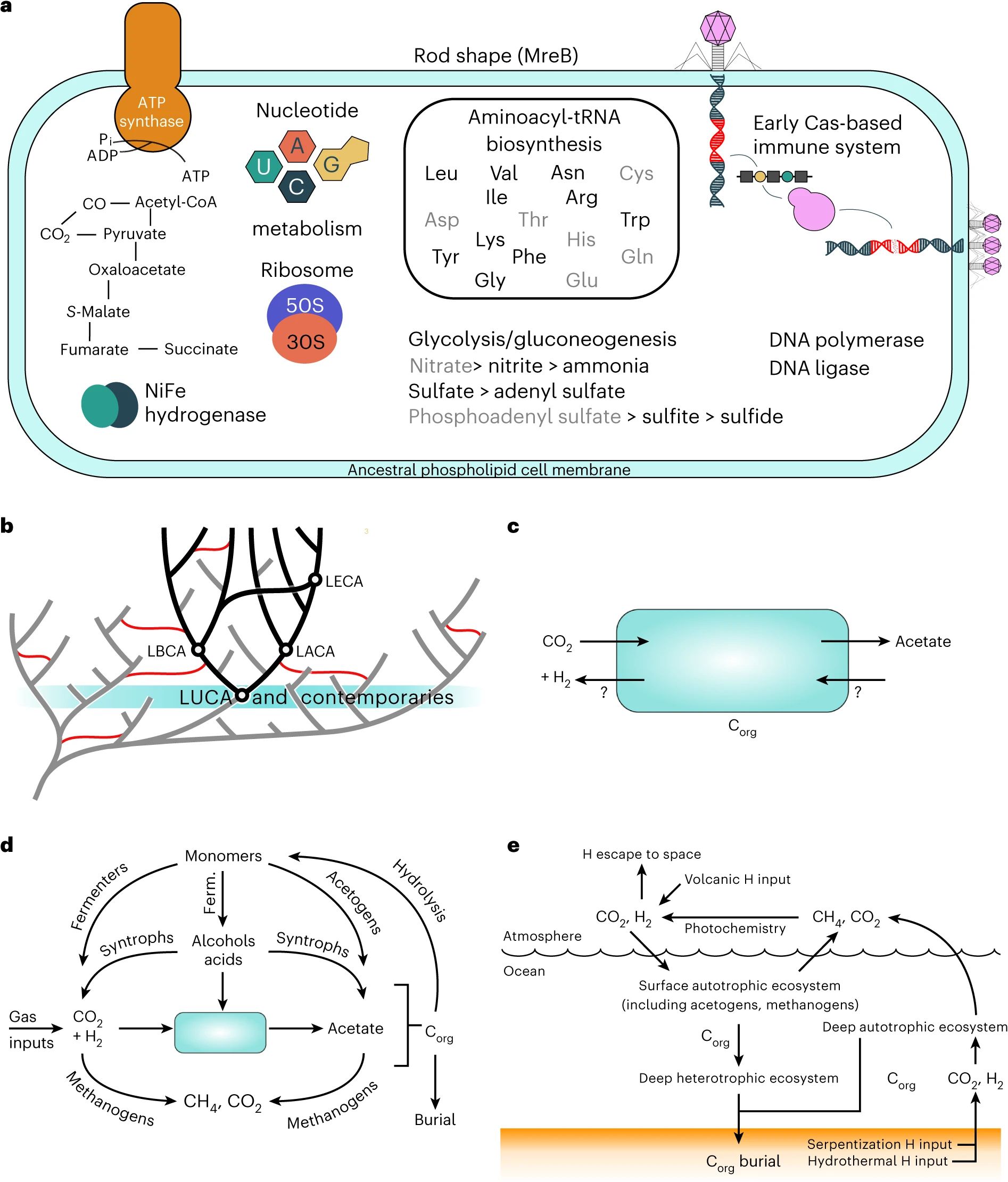
图3:在其进化和生态背景下的LUCA重建。
a,基于我们祖先基因内容重建的LUCA表示。黑色基因名称是在最严格阈值(PP=0.75,在两个域中都有抽样)下推断存在于LUCA中的;灰色的是在最不严格阈值(PP=0.50,不需要在两个域中都存在)下存在的。b,生命之树背景下的LUCA。生命之树上留下了今天被抽样后代的分支被涂成黑色,那些没有留下被抽样后代的分支是灰色的。作为现存细胞生命的共同祖先,LUCA是可以使用系统发生学方法重建的最古老节点。它将与早期地球上的其他谱系(以蓝绿色突出显示)共享地球,这些谱系在今天的被抽样细胞生命中没有留下后代。然而,这些谱系可能在灭绝前通过将基因转移到被抽样的生命之树(红线)中而在现代生物中留下了痕迹。c,LUCA的化学自养代谢很可能依赖于与直接环境的气体交换来实现有机碳(Corg)的固定,通过产乙酸作用,并且可能还以相反的方式运行代谢。d,早期生态系统背景下的LUCA。推动LUCA可能是产乙酸代谢的CO2和H2可能来自地质化学和生物输入。LUCA产生的有机物和乙酸可能为其他代谢方式创造了生态位,包括那些回收CO2和H2的代谢方式(如现代沉积物中)。e,地球系统背景下的LUCA。产乙酸的LUCA可能是表面和深层(化学)自养生态系统的关键部分,由H2提供动力。如果甲烷菌也存在,氢气将以CH4的形式释放到大气中,通过光化学反应转化为H2,从而循环回到表面生态系统,提高其生产力。发酵。
三、基因组大小和细胞特征
通过使用现代原核生物基因组作为训练数据,我们使用预测模型来估计LUCA的基因组大小和编码的蛋白质家族数量,基于现代原核生物基因组编码的总蛋白质数量与KEGG基因家族数量之间的关系。基于KEGG KO基因家族的PPs,我们确定了399个KOs的一个保守子集,这些KOs很可能存在于LUCA中,PPs≥0.75,并且在古细菌和细菌中都有发现;这些家族构成了我们代谢重建的基础。然而,通过整合所有KO基因家族推断的PPs,包括那些概率低的,我们也估计了LUCA的基因组大小。我们的预测模型估计基因组大小为2.75 Mb(2.49-2.99 Mb),编码2657(2451-2855)个蛋白质(方法)。虽然我们可以估计LUCA基因组中的基因数量,但根据现代古细菌和细菌的基因组来确定哪些特定基因家族可能已经存在于LUCA中则更加困难。基于LUCA的基因内容,现代版本的通路可能会因随后的进化变化而被认为是不完整的。因此,我们应该预期由于这种系统发生噪音和分析管道的其他限制,代谢途径的重建是不完整的。例如,当查看可以毫无争议地映射到LUCA的基因和途径时,如实施遗传密码的核糖体和氨酰-tRNA合成酶,我们发现我们将许多(但不是全部)关键组分映射到LUCA。我们将其解释为意味着我们的重建可能是不完整的,但我们对LUCA代谢的解释依赖于我们对途径的推断,而不是单个基因。
LUCA的推断基因内容表明它是一种厌氧生物,因为我们没有找到支持存在末端氧化酶的证据。相反,我们几乎识别出了编码古细菌版本(以及大多数细菌版本的)伍德-隆达尔途径(WLP)的所有基因(PP>0.7),这表明LUCA具有乙酸生成生长和/或碳固定53,54,55的潜力。LUCA编码了一些NiFe氢化酶亚基(K06281,PP=0.90;K14126,PP=0.92),这可能使其能够在氢气上生长。我们推断,参与产甲烷作用的复合物,如甲基辅酶M还原酶和四氢甲烷吡咯S-甲基转移酶缺失,这表明LUCA不太可能像现代的甲烷菌那样起作用。我们发现对TCA循环的一些组分(包括酮戊二酸/2-氧代酸铁氧还蛋白氧化还原酶(K00175和K00176)、琥珀酸脱氢酶(K00239)和高柠檬酸合成酶(K02594)的亚基)的支持很强,尽管有些步骤缺失。考虑到WLP的存在,这可能表明LUCA能够进行糖异生/糖解作用,因为我们发现支持这些途径中涉及的大多数酶亚基的证据。考虑到存在WLP,这可能表明LUCA具有有机异养生长的能力,也可能具有自养生长的能力。糖异生在将碳固定与通过五磷酸核糖途径的核苷酸生物合成联系起来方面将是重要的,LUCA似乎存在该途径的大多数酶。我们没有发现LUCA是光合作用的证据,几乎所有氧气光系统和非氧气光系统的组件的后验概率都很低。
我们强烈支持ATP合酶的存在,特别是亲水V/A1亚单位的A(K02117,PP=0.98)和B(K02118,PP=0.94)亚单位组分,以及跨膜V/A0亚单位的I(亚单位a,K02123,PP=0.99)和K(亚单位c,K02124,PP=0.82)亚单位。此外,如果我们放宽抽样阈值,我们还推断出F1型β亚单位(K02112,PP=0.94)的存在。这与许多之前的研究一致,这些研究将ATP合酶亚单位映射到了LUCA。
我们对硝酸钾同化途径(铁氧还蛋白-硝酸钾还原酶,K00367,PP=0.69;铁氧还蛋白-亚硝酸钾还原酶,K00367,PP=0.53)和硫酸盐还原途径(硫酸腺苷转移酶,K00957,PP=0.80,和K00958,PP=0.73;亚硫酸盐还原酶,K00392,PP=0.82;磷酸腺苷磷硫酸还原酶,K00390,PP=0.56)的存在获得了适度支持,可能是为了促进氨基酸生物合成,我们推断出了37个部分完整的途径。
我们在LUCA的基因组中发现了19个第1类CRISPR-Cas效应蛋白家族的支持,包括I型和III型(cas3, K07012, PP=0.80, 和K07475, PP=0.74; cas10, K07016, PP=0.96, 和K19076, PP=0.67; 以及cas7, K07061, PP=0.90, K09002, PP=0.84, K19075, PP=0.97, K19115, PP=0.98, 和K19140, PP=0.80)。Cas1和Cas2的缺失可能表明LUCA编码了一个早期的Cas系统,具有提供基于RNA的免疫反应手段的能力,通过切割(Cas6/Cas3)和结合(CSM/Cas10)RNA,但缺乏完整的免疫系统位点CRISPR。这支持了CRISPR-Cas免疫的效应阶段是从RNA感应的信号转导进化而来的观点,基于蛋白质的RNA结合模块的相似性58。这与细胞生命在LUCA时期已经卷入与病毒的军备竞赛的观点一致59,60。我们的结果表明,一个早期的Cas系统是现存细胞生命的祖先免疫系统。
总的来说,我们的代谢重建表明LUCA是一种相对复杂的生物体,类似于现存的古细菌和细菌。基于在LUCA之前Sec和ATP合酶基因的古老复制,以及这些系统的关键组分的高后验概率(PP),膜结合的ATP合酶亚单位、参与肽聚糖合成的基因(mraY, K01000; murC, K01924)和细胞骨架类似肌动蛋白的蛋白质MreB(K03569)(补充数据3),LUCA很可能拥有现代原核生命的核心细胞器。这可能包括磷脂膜的基本成分,尽管我们的分析并未最终确定其组成。特别是,我们恢复了以下涉及醚和酯类脂质合成的酶(烷基二羟基丙酮磷酸合成酶、甘油3-磷酸和甘油1-磷酸)以及甲瓦龙酸途径的组分(甲瓦龙酸5-磷酸脱水酶(PP=0.84)、羟甲基戊二酰辅酶A还原酶(PP=0.52)、甲瓦龙酸激酶(PP=0.51)和羟甲基戊二酰辅酶A合成酶(PP=0.51))。
与之前对LUCA基因内容的估计相比,我们发现与参考文献7的共识数据集有81个重叠的COG基因家族,与参考文献6的数据集有69个重叠的KOs。之前研究之间的关键点一致包括存在信号识别颗粒蛋白ffh(COG0541, K03106)7用于质膜的蛋白质定位和递送,大量的氨基酰-tRNA合成酶用于氨基酸合成和糖酵解/糖异生酶。
参考文献6推断LUCA为一种使用WLP进行碳固定的热原厌氧自养生物,基于单一酶(CODH)的存在,并同样暗示LUCA能够利用固氮酶进行氮固定。我们的重建与参考文献6一致,认为LUCA是使用WLP进行碳固定的厌氧自养生物,但我们推断出的存在一个比之前获得的更完整的WLP。我们没有找到固氮酶或氮固定的有力证据,且重建对于LUCA的最佳生长环境并不明确。
我们使用了概率方法来重建LUCA——即,我们根据基因家族沿总体物种树演化的模型,估计每个基因家族存在于LUCA中的概率。这种方法与分析系统发育存在-缺失特征或那些使用过滤标准(如广泛分布或高度垂直演化家族)来定义可能在LUCA中存在的现代基因高置信子集的分析不同。我们的重建将更多基因映射到LUCA——尽管每个基因的概率较低——比以前的分析得到的结果是现代原核生物范围内的LUCA基因组大小估计。结果是一个不完全的细胞生物图像,它是原核级别的而非原核生物前体,并且与今天的原核生物相似,可能作为一个生态系统的一部分存在。作为抽样现存原核生命的共同祖先,LUCA是我们可以通过系统基因组学重建的最古老的节点,但如图3所示,它已经是进化历史上一个高度创新时期的产品,在这一时期大多数细胞核心组分都已建立。按照定义,我们不能使用系统基因组学重建LUCA的同时代生物,但我们可以基于重构的LUCA提出关于它们生理学的假设,其特征直接暗示了与其他原核代谢相互作用的可能性。
四、LUCA的环境、生态系统和地球系统背景
推断LUCA使用WLP有助于限定其可能生活的环境和生态。现代产乙酸菌可以在H2(和CO2)上自养生长,或在包括醇类、糖类和羧酸在内的广泛替代电子供体上异养生长。这种代谢的灵活性是它们现代生态成功的关键。无论是自养还是异养,产乙酸作用的能量产出和生长效率都较低(尽管使用还原性乙酰辅酶A途径进行能量生产和生物合成减少了生物合成的能量成本)。这与能量受限的早期生物圈一致。
如果LUCA作为有机异养的产乙酸菌发挥作用,那么它必然是一个包含自养生物的生态系统的一部分,这些自养生物提供了有机化合物的来源(因为在早期地球上有机物分子的非生物来源通量很小)。另一方面,如果LUCA作为化学自养的产乙酸菌发挥作用,原则上它可以独立于非生物的H2(和CO2)来源生活。然而,LUCA不可能孤立存在,因为它的化学自养生理代谢的副产品会为其他代谢类型的共生创造生态位(如图3d所示)(如图3d所示)。这包括LUCA本身作为有机异养生物生长的可能性。
化学自养的产乙酸型LUCA可能占据了两个主要的潜在栖息地(图3e):第一个是深海,其中热液喷口和海底蛇纹石化作用提供了H2的来源。与此一致,我们发现支持逆转录酶的存在(PP=0.97),这是超嗜热原核生物的标志酶6,63,64,65,如果早期生命存在于海洋表面,则不会期望出现这种情况(尽管逆转录酶的进化很复杂;见补充信息中的“逆转录酶”)。第二个栖息地是海洋表面,大气层提供了来自火山和变质作用的H2源。实际上,我们检测到了孢子光解酶的存在(COG1533, K03716, PP=0.88),在现存生物中修复因紫外线(UV)辐射诱导而在孢子DNA中发生的亚甲桥胸腺嘧啶二聚体。然而,这个基因家族也出现在既不形成内生孢子也不生活在可能对其DNA造成UV损伤的环境中的现代分类群中,因此并非暴露于UV环境的独有标志。以前的研究通常倾向于认为LUCA生活在深海环境中,因为那里早期生命更能免受LHB事件的影响。然而,如果LHB事件不如最初提出的那样强烈,或者仅仅是采样偏差,这些论点就站不住脚了。另一种可能性可能是LUCA栖息在浅层热液喷口或温泉中。
在这些生态系统中,早期地球上的氢流量可能比今天高出几倍(由于内部热源更大)。今天的火山活动产生约1×10^12摩尔H2/年,蛇纹石化作用产生约0.4×10^12摩尔H2/年。根据当前的H2流量和已知的H2逃逸率与空间的关联,预测非生物大气中的H2浓度约为150ppmv。化学自养的产乙酸菌会在局部降低H2浓度(无论是在表面还是深层生态位),但它们的低生长效率确保了H2(和CO2)仍然可用。这以及产生的有机物质和乙酸将为其他代谢类型创造生态位,包括产甲烷作用(图3d)。
基于热力学考虑,CH4和CO2预计是最终的代谢产物,一小部分初始消耗的氢以有机物形式。CH4向大气的通量将为表面海洋的光化学H2再生和相关生产力提供燃料(图3e)。现有模型表明,由此产生的全球H2循环系统非常有效,以至于到达表面的H2供应通量可能超过火山对大气的H2输入至少一个数量级,这意味着这样的生物圈的生产力得到了相应的提升。光化学再循环到CO也会支持消耗CO的生物的表面生态位。
在深海栖息地,可能会有一些局部的电子再循环(图3d),但由于高度不溶的H2和CH4向大气层的定量损失以及在CH4光化学转化为H2后返回最小,全球性的深度再循环将是最小的(图3e)。因此,LUCA的表面环境可能成为主导(尽管由此产生的有机物的再循环可以通过海洋深度传播;“深层异养生态系统”图3e)。包括产乙酸型LUCA和产甲烷菌在内的早期化学自养生物圈的全球净初级生产力可能在~1×10^12到7×10^12摩尔C/年之间(比今天低约3个数量级)。
要支持这样一个生物圈所需的营养供给(例如,氮)需要与有机物埋藏通量中损失的相平衡。地球表面的氧化还原平衡规定,氢向空间的损失和电子/氢的埋藏必须共同平衡电子/氢的输入。考虑到当前的H2输入和上述净初级生产力的估计,这表明最大埋藏通量在~1012摩尔C/年的数量级,按照当代的化学计量学(碳氮比约为7),这可能需求超过1011摩尔N/年。雷电将提供亚硝酸钾和硝酸钾的来源70,这与LUCA推断的亚硝酸盐和(可能的)硝酸盐还原途径一致。然而,它只有3×109摩尔N/年的量级。相反,在一个全球性的氢气循环系统中,大气中较高处的光化学反应产生的HCN被沉积并在水中水解为氨,将可利用的氮供应增加了数个数量级,达到约3×1012摩尔N/年(参考文献71,72)。这条HCN途径与最早可能的生物物质(3.8–3.7亿年前)异常轻的氮同位素组成一致(参考文献73),尽管那大大晚于我们推断的LUCA的时代。这些考虑表明,所提出的LUCA生物圈(图3e)将是能量或氢限制而非氮限制。
五、结论
通过概率性处理基因存在,我们的重建将许多更多基因(2,657个)映射到LUCA,比以前的分析结果多,并且得出了LUCA基因组大小(2.75Mb)的估计,这在现代原核生物的范围内。结果是对一个细胞有机体的描述,它是原核级别的而不是前原核的2,并且很可能作为一个生态系统的组成部分存在,使用WLP进行产乙酸生长和碳固定。我们不能使用系统发育学来重建这个早期生态系统的其他成员,但我们可以根据LUCA的代谢输入和输出来推断它们的生理学。从生命起源到LUCA时期早期群落的进化如何进行仍是一个未解决的问题,但推断的LUCA年龄(约4.2亿年前)与地球和月球的起源相比表明,这个过程需要一段令人惊讶地短的地质时间间隔。
https://blog.sciencenet.cn/blog-41174-1442270.html
上一篇:CAR-T疗法对胶质母细胞瘤的成功案例
下一篇:氢气药理作用和给药途径【综述】
