博文
花色的演变|从色彩缤纷的球果最初演变至人类世演变  精选
精选
|
【内容提要】
花色的演变是自然引人注目的现象,其奥秘涉及色彩本质和不同观察者视角下的色彩差异。花色的分子和生化基础主要基于色素合成途径,其演变过程可从起源、宏观进化、微观进化及人类影响四个时间维度进行审视。
鲜艳的花色有助于吸引传粉者,提高被子植物的繁殖成功率,而现存裸子植物的生殖结构色彩对比不明显。开花植物祖先在吸引传粉昆虫和减少食草动物之间进行权衡,对现存裸子植物颜色的进化和生物学意义,球果苞片上的红色可能是花青素色素积累的结果,最初作为视觉线索进化,可能在蜜蜂色觉进化之后,但早于其他花色素功能的积累。
花和颜色
颜色由两个部分组成。首先,它需要某些而不是所有波长的光进行不同的反射。一张白纸没有其他颜色,因为它对所有波长的反射都相同并且很强烈,而黑色键盘也没有其他颜色,因为它对所有波长的反射都相同但反射效果很差,反而吸收了它们。然而,一朵红色的花(图 1A)如毛茛科欧侧金盏花(Adonis annua)会吸收蓝色、黄色和绿色波长,仅反射红色波长。那么它是红色的吗?这取决于颜色的第二个组成部分:具有存在能够感知不同波长的生物,特别是感知红色能力的生物。对于人眼来说,这朵花确实是红色的,因为我们的眼睛含有受红光刺激的光感光器以及其他受绿光和蓝光波长刺激的光感受器。但是,对于像蜜蜂这种的动物来说,在光谱的红色部分几乎没有或没有光感受能力,尽管在绿色、蓝色和紫外线区域有很好的感知力,这种花很可能没有颜色,而只是黑色。对颜色的两个组成部分都需求,因此在自然世界中光波长的不同反射有可能被用作一种信号系统,相同的反射光谱对不同的接收者意味着不同的色彩意义。
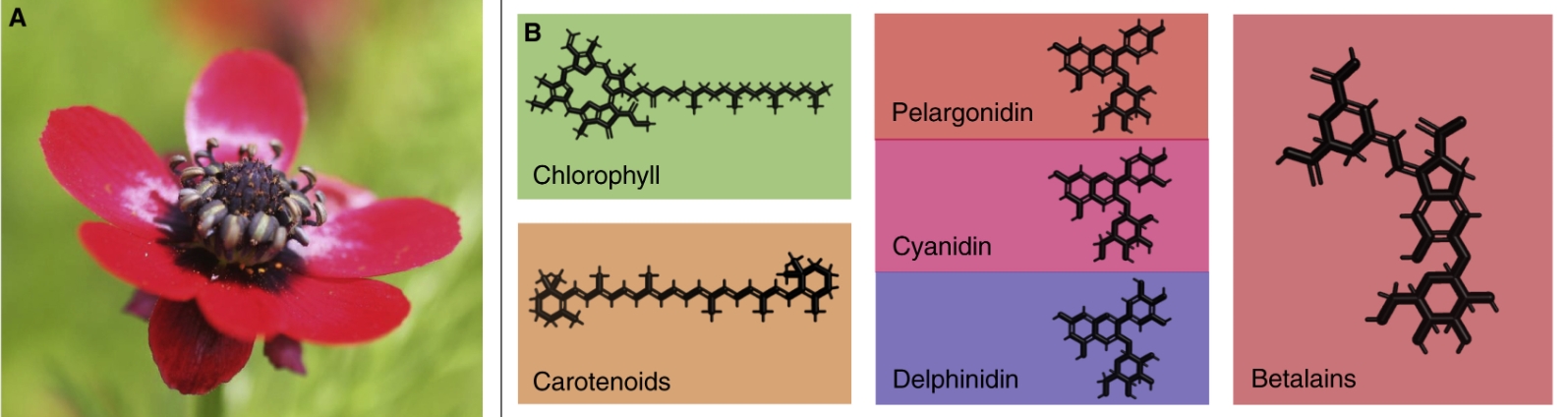
图1 花色的特性
(A) 欧侧金盏花的红花吸收蓝光和绿光,只反射和透射红光。(B) 花色素的主要类别包括绿色的叶绿素、黄橙色的类胡萝卜素、花青素(包括橙色鹅掌楸素、品红色青花素和蓝色鹅掌楸素)和黄红色甜菜碱。
花是如何着色的?
令人惊讶的是,花的各种颜色仅由叶绿素、类胡萝卜素、花青素和甜菜碱等少数几种不同的色素产生,此即色素色(图 1B)。绿色叶绿素和黄色/橙色至红色脂溶性类胡萝卜素都存在于质体中,在光合作用中具有重要功能。相比之下,水溶性的花青素是一类类黄酮,储存在液泡中,在细胞紫外线防护中发挥作用。花青素生物合成途径的分支可产生橙色的天竺葵素(pelargonidins)、品红色花青素(cyanidins)和蓝色的飞燕草素(花翠素,delphinidins)。大多数其他类黄酮在可见范围内(几乎)是无色的,但有助于形成紫外线图案。最后,石竹目(Caryophyllales)特有的黄色到红色的甜菜碱来自于酪氨酸,与花青素的存在相互排斥。人们对这些色素的生物合成途径进行了深入的研究,进化过程中的持续修补似乎使有限的遗传元素产生了巨大的多样性。
特别是花青素的生物合成在不同的植物中以各种方式进行修饰,从而可以精确地微调花色色调。WDR-bHLH-MYB 转录因子复合物及其调节的通路基因的时空表达变化可以重新定向花青素通路分支之间的代谢物通量,并且多个组织特异性转录因子的亚功能化可以实现花的模式。在它们生物合成后,pH 值变化、分子堆积和化学修饰(例如添加糖苷侧基或与金属离子螯合)可以进一步改变花青素化合物的色调。最后,花色素沉着可以与花瓣细胞的生物物理特性相互作用,产生额外的光学效果,例如天鹅绒般的表面和花的彩虹色。
花色的起源和深度演变
现代五颜六色的花瓣来源于叶状结构,被认为在早期起到了保护花内部生殖器官的作用。松果的鳞片也起着类似的作用有时也会有颜色。松柏类如西班牙冷杉(Abies pinsapo)等物种的球果(图 2A)之所以呈现醒目的红色,是因为花青素色素积累在鳞片中,吸收了除红色波长以外的所有光线。这些风媒授粉结构的颜色被认为是为了防止强光照射、增加发育种子周围的温度,甚至是为了防止草食动物的侵害。因此,被子植物和裸子植物的共同祖先的保护性不育器官以及最早的花朵周围的不育器官,可能已经是鲜艳的颜色,但其原因与吸引传粉者无关。然而,被子植物花和裸子植物的球果并不一定是同源结构,源自相同祖先形式。关于花的起源,以及相应的彩色花瓣的起源,存在着相当多的争论。

图2 花色的起源和宏观演变
(A)西班牙冷杉(Abies pinsapo)的红色球果(拍摄于剑桥大学植物园)。(B)无油樟(Amborella trichopoda)淡奶油色花(拍摄于剑桥大学植物园)。(C)由熊蜂授粉的狗面花属Mimulus lewisii(粉红色,左)和蜂鸟授粉的Mimulus cardinalis(红色,右)的正面和侧面图。这两个物种之间花色的相互渐渗诱导了传粉媒介的转换(图片由 Kelsey Byers 博士提供)。
在发育过程中,花瓣明然与叶状结构同源,叶状结构应该是绿色的。一个简单的变化,例如叶绿素缺失,就会产生白色的花瓣(因为纤维素细胞壁是所有波长的扩散器)。这可能对传粉媒介的吸引有一些好处,因为即使是全白的花瓣也能与绿色植被形成对比,特别是在近距离时,当 “绿色对比度”(目标和背景的相对反射率,特别是在绿色通道中)在昆虫颜色处理中变得不那么重要。早期分化的被子植物谱系通常会产生颜色较淡的花,例如无油樟(Amborella trichopoda)的奶油色花(图 2B),或许多睡莲的白色花。然而,在其中一些类群中也发现了更强烈的颜色,包括橙色、粉红色和一些蓝色,这表明在被子植物类群中,通过吸收在光保护、光合采光或应激反应中发挥着其他作用的色素,花色的演化相对较早。
当我们考虑到主要的授粉群体昆虫对颜色的感知能力时,花色的快速多样化可能并不足为奇了。对昆虫眼中的感光色素视蛋白的编码基因的进化历史进行的分析表明,所有昆虫和甲壳类动物的盘古祖先很可能拥有编码多种不同视蛋白的基因,因此能够区分长、中和短波长的光。尽管对昆虫和花的起源时间存在争议,但人们普遍认为,昆虫比被子植物早一亿多年。对于这些对颜色敏感的动物来说,景观中出现的第一批彩色花一定显得非同寻常,为植物和动物之间的信号传递开辟了新的可能性。
花色宏观演变
纵观现存花的无穷形态及其传粉者,似乎可以很直观地将被子植物的快速发展——达尔文的“可憎之谜”——归因于植物与传粉者之间信号传递的变化。植物与传粉媒介的相互作用显然不是被子植物成功的唯一原因,但可以说是今天看到的被子植物多样性的主要原因。与其他花信号如气味、形状和奖励一起,花色在这种持续多样化的过程中发挥着重要作用。花色的变化可以使以前不感兴趣的动物类群看到花朵,从而将这些动物招募为新的传粉者。这已在实验中得到证明,例如,在透骨草科狗面花属(Mimulus)中,大黄蜂授粉的Mimulus lewisii具粉红色花和蜂鸟授粉的Mimulus cardinalis具红色花,它们的花色相互渗入降低了其相应的本地传粉媒介的访问率,但反而引发了其他物种的传粉者的访问(图 2C),从而诱导了传粉媒介的转换。
一旦在特定分类群的植物和动物之间建立了授粉相互作用,花色变化就可以进一步加速植物多样化,因为它会引发从一个传粉媒介谱系到另一个传粉媒介谱系的转变,植物和传粉媒介之间的共同进化种族,或者从许多到少数或只有一个传粉媒介物种的花专业化。
人类试图检测由此产生的花卉多样性中的模式,导致了授粉综合征的定义,即在由同一动物顺序授粉的多个植物物种中独立进化的特定花性状组合。花的颜色是这一分类的核心组成部分,不同的授粉综合症颜色与相应传粉媒介组中的光感受器敏感性和光信号处理相匹配。例如,蜜蜂授粉植物的蓝色和黄色花朵、与鸟类色觉相匹配的红色花朵,以及黄昏和夜行性飞蛾物种光顾的白色紫外线反射花朵。授粉综合征的有效性和有用性已被广泛争论。例如,对 耧斗菜属(Aquilegia)、钓钟柳属(Penstemon)和矮牵牛属(Petunia)的辐射研究提供了支持,在这些研究中,仅通过观察花朵就可以预测相应的蜜蜂、鸟类或飞蛾传粉者,有时甚至可以预测到物种水平。
然而,在大约三分之二的植物类群和环境中,授粉综合征充其量只是指示性的,有许多不太特化的花朵由不止一个目的的动物授粉的例子。最近的一项元研究进一步得出结论,颜色是传粉媒介预测中最无信息量的花性状之一,很可能是因为它经常受到不同传粉媒介和食草动物群体的对抗性选择。因此,虽然花色变化是传粉媒介介导的植物宏观进化的重要驱动力,但花色的大规模系统发育模式与传粉媒介多样性之间的关系很少是线性的。
花色微进化
在整个进化生物学的历史中,花色一直是研究多样化和物种形成过程的模型性状。与许多其他表型变异的例子不同,花的颜色变化很容易感知和量化,它们通常直接影响植物的生殖适应性,分子研究受益于花色素沉着的保守和充分理解的遗传基础。花色变异在文献中突出的另一个可能原因是,这种类型的变异比其他性状的变异更常见。这可以用色素生物合成途径的不稳定性来解释。由于大多数花色素是次级化合物,在初级代谢中没有关键作用,因此潜在途径基因和转录因子的突变通常几乎没有有害的多效性作用。缺乏内在致死性使颜色突变体能够在种群中持续存在,直到开花阶段,从它们突然受到传粉者和其他外在生物和非生物因素的不同选择压力开始。
这个过程的结果可以分为三种主要情况(图 3A)。在第一种也可能是最有可能的情况下,新的颜色变化再次消失,要么是由于遗传漂变,要么是由于选择性劣势,例如对传粉媒介的能见度或吸引力降低,或对食草动物的能见度增加。还有研究表明,花青素颜色的完全丧失,一种频繁的花色突变,使配子暴露在更高剂量的紫外线辐射下,从而降低它们的生存能力,尤其是在高海拔栖息地(图 3B)。

图3 花色微进化
(A)新的花色形态要么由于选择性劣势而再次消失(左图),要么因形态替换或传粉媒介转移而导致生态物种分化(中图),要么产生稳定的多态性种群(右图)。(B)(近)完全丧失色素通常会带来选择性劣势,拟蝶唇兰(Psychopsis papilio)。(C)花的颜色变化,如花粉红色的草甸鼠尾草(Salvia pratensis)个体,可以成为传粉媒介介导的生态物种分化的基础。(D)波动和平衡选择可将花色多态性锁定在微进化状态,如在阿尔卑斯手参(Gymnadenia rhellicani)过量表达系统中(所有图像均由 Roman Kellenberger 拍摄)。
在第二种情况下,新的花色赋予了选择优势。如果当前的传粉者喜欢它,它可能会随着时间的推移取代种群中的祖先颜色变化。如果它介导了向新的传粉媒介物种的转变,这会导致两种颜色形态之间的不同选择。这两个例子最终都可能导致生态物种形成并完成从微观进化到宏观进化的过渡(图 3C)。花色的种间差异也可能源于单独的遗传漂变或花色位点与选择中的其他遗传元素的联系,但目前的文献表明这些过程起着次要作用。
第三种情况相当奇特。在这种情况下,新的花色也赋予了选择优势,但作用于整个种群的选择阻止了任何一种颜色形态取代另一种颜色,从而将花色多态性锁定在微进化状态。导致这种情况的主要过程是选择强度和方向的空间或时间变化,例如,兰花阴阳掌裂兰(Dactylorhiza sambucina)中令人惊叹但有争议的负频率依赖性选择,以及由两个或多个拮抗选择力相互抵消而导致的平衡选择,例如,黄花帝王花(Protea aurea)中的拮抗多效性,普通牵牛花(Ipomoea purpurea)中的杂合子优势,以及在高山兰花阿尔卑斯手参(Gymnadenia rhellicani)中的优势,图 3D)。
人类世的花色演变
除了传粉者、不受欢迎的食草动物和其他动物将它们用作庇护所、狩猎场或以其他方式,花还吸引了另一种类型的访客——智人的崇拜者。尼安德特人墓葬中的花粉、埃及墓葬中的花卉图画以及来自希腊、罗马、中国、玛雅和其他古代文明的带有花卉图案的文物表明,花卉与人类之间的关系是泛文化的,可以追溯到很久以前。这令人费解,特别是因为除了潜在的药物应用外,鲜花不会为人类提供任何明显的回报。然而,心理学实验表明,花会引起人类的即时和长期积极情绪,而且由于我们配备了出色的三色视觉,因此花的颜色在此过程中起着重要作用。
对于灵长类动物和早期狩猎采集者来说,鲜艳的颜色与有价值和美味的高营养水果和其他食物来源有关。这导致一些科学家推测,花在人类的情感奖励中占据了一个进化生态位。无论这种关系的起源如何,它都深深地影响了双方。17 世纪的“郁金香狂热”就是一个例子,如今花卉栽培是一个价值数十亿美元的大型产业(图 4A),为从出生到结婚再到葬礼的关键生活事件提供产品。人类尝试使用园艺技术(如嫁接、组织培养和转基因)生产新的高价值花卉作物,这导致了在野外永远无法生存的花卉颜色品种的出现和保存。突出的例子包括嫁接的观赏玫瑰(图 4B)、病毒感染诱导的带有条纹花瓣的郁金香,或来自不同大陆的多个兰花属之间的杂交五颜六色的兰花(图 4C)。这种人工产生的花卉颜色变化与人类引起的栖息地加速破坏和野花及其传粉媒介的灭绝形成鲜明对比。我们只能希望,从我们过去和现在与花朵的关系中获得的知识和技术可以用来保存花色自然进化的多样性。

图4 园艺中的花色 Floral colour in horticulture
(A)花瓣颜色不规则的蝴蝶兰栽培品种。这些植物通过组织培养大量繁殖,并在世界各地的园艺中心和超市出售。(B)现代条纹玫瑰品种,是广泛的长期育种计划的结果。(C) × Renanthopsis ‘Mildred Jamson’ 是兰科火焰兰属(Renanthera)和蝴蝶兰属(Phalaenopsis)之间的属间杂交种(所有图像均由 Roman Kellenberger 拍摄)。

图5. 通过人类眼睛(1-6)、紫外线(7-12)和蜜蜂眼睛(12-18)使用假彩色摄影看到的花朵颜色变化,说明花朵颜色变化被忽视的潜在原因和研究系统选择的偏差
由生物授粉的被子植物,通常会具有颜色鲜艳、对比强烈的花,这有助于吸引传粉者,从而提高物种的繁殖成功率。这种五颜六色的花与四类现存裸子植物多彩生殖结构形成鲜明对比,它们大多是靠风媒授粉,而苏铁类和买麻藤目主要由昆虫授粉,昆虫吸食授粉滴表面的液体。这篇综述研究了现存种子植物和化石种子植物花色的可能进化途径和神秘线索,探讨了祖先开花植物如何克服在吸引传粉昆虫和减少食草动物之间存在的不可避免的权衡问题,并讨论了从一些现存裸子植物出现的颜色可能的进化和生物学推论。一些现存针叶树种的球果苞片上的红色是花青素色素积累引起的,它们的主要功能可能是在特定环境条件下帮助保护生长的植物组织。因此,花瓣的色彩所提供的视觉线索最初可能首先是作为一种次要效应进化的,可能发生在蜜蜂色觉进化之后,但发生在一系列不同花色素随后的功能积累之前(图6-8)。

图6 球果红色肉质结构
(A)山岭麻黄(Ephedra gerardiana),隶属于买麻藤目(Gnetales)麻黄科(Ephedraceae) ,成熟的肉质圆锥形苞片,最初是绿色,然后是黄色,然后变成红色。(B)欧洲红豆杉(Taxus baccata),红豆杉科(Taxaceae),种子周围有红色肉质假种皮。(C)台湾穗花杉(Amentotaxus formosana),红色肉质假种皮,圆锥苞片也是彩色的。(D)罗汉松属Podocarpus oriarius,罗汉松科(Podocarpaceae),每颗种子下方有肉质假种种皮,从黄色到成熟时红色。(E-G)丽江麻黄(Ephedra likiangensis),麻黄科(phedraceae),处于授粉阶段球果苞片肿胀和肉质之前,球果苞片在侧翼呈黄色,在中央区域呈绿色。(F)扫描电子显微镜照片显示种子周围的圆锥苞片(去除外苞片)。气孔和圆锥形细胞都存在于暴露区域 (scale=0.5 mm)。(G)(F)中圆锥苞片表面的细节,显示锥形细胞和两个气孔,保护细胞深陷在表面以下(不可见)(刻度 = 20 μm)。图片 A-D 由 Martin Gardner 提供,其余部分由作者提供。

图7 针叶树雌球果
针叶树雌球果(有时称为排卵球果或雌球果)。(A - D)欧洲落叶松(Larix decidua, 松科)。(A) 2019 年 5 月,邱园皇家植物园的树上球果,显示出突出的红色苞片。(B) 圆锥体正中纵切面的扫描电子显微镜照片,显示突出的苞片掩盖了较小的卵形鳞片(比例尺 = 1 毫米)。(C) 苞片近轴(上)表面的细节,显示两排气孔包围中脉,苞片其余部分相对平坦的细胞(比例尺 = 200 μm)。(D) (C) 的细节(比例尺 = 50 μm)。(E-G)青杄(Picea wilsonii, 松科)。(E) 2019 年 5 月,邱园皇家植物园雌球果,显示球果具有淡红色的卵形鳞片。(F) 卵形鳞片扫描电子显微镜照片:背面(外)表面有扁平的表皮细胞,显示气孔的新月形区域(比例尺 = 1 mm)。(G) 卵形鳞片的扫描电子显微镜照片:近轴(内)表面有平坦的表皮细胞,缺乏气孔(比例尺 = 1 mm)。
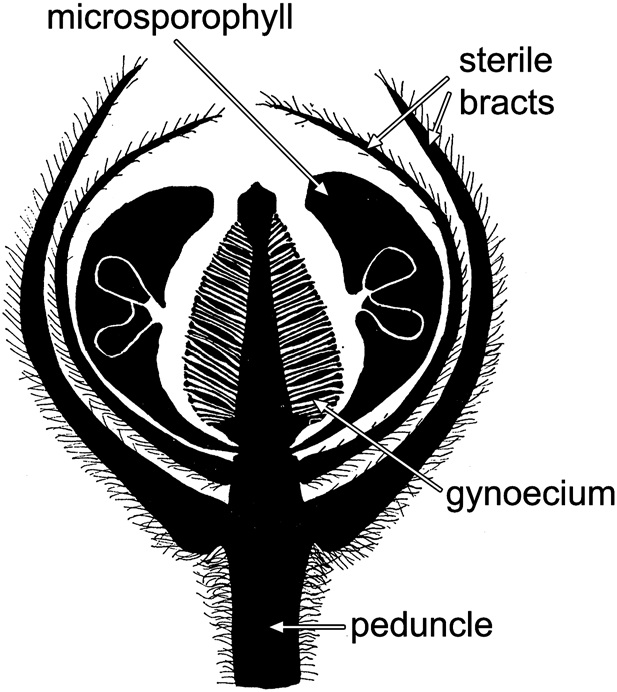
图8 本内苏铁目小威廉姆逊花纵切面图
延伸阅读
1.Bradshaw, H. ∙ Schemske, D.W. Allele substitution at a flower colour locus produces a pollinator shift in monkeyflowers. Nature. 2003; 426:176-178
2. Dellinger, A.S. Pollination syndromes in the 21st century: where do we stand and where may we go? New Phytol. 2020; 228:1193-1213
3. Grotewold, E. The genetics and biochemistry of floral pigments Annu. Rev. Plant Biol. 2006; 57:761-780
4. Huss, E. ∙ Bar Yosef, K. ∙ Zaccai, M. Humans’ relationship to flowers as an example of the multiple components of embodied aesthetics Behav. Sci. 2018; 8:32
5. Henze, M.J. ∙ Oakley, T.H. The dynamic evolutionary history of pancrustacean eyes and opsins Integr. Comp. Biol. 2015; 55:830-842
6. Narbona, E. ∙ del Valle, J.C. ∙ Arista, M. ...Major flower pigments originate different colour signals to pollinators Front. Ecol. Evol. 2021; 9, 743850
7. Rudall, P.J. ∙ Bateman, R.M. Defining the limits of flowers: the challenge of distinguishing between the evolutionary products of simple versus compound strobili Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2010; 365:397-409
8. Rudall, P.J. Colourful cones: how did flower colour first evolve? J. Exp. Bot. 2020; 71:759-767
9. Sapir, Y. ∙ Gallagher, M.K. ∙ Senden, E. What maintains flower colour variation within populations? Trends Ecol. Evol. 2021; 36:507-519
10. Spaethe, J. ∙ Tautz, J. ∙ Chittka, L. Visual constraints in foraging bumblebees: flower size and color affect search time and flight behavior Proc. Natl. Acad. Sci. USA. 2001; 98:3898-3903
11. Wessinger, C.A. ∙ Rausher, M.D. Lessons from flower colour evolution on targets of selection J. Exp. Bot. 2012; 63:5741-5749
https://blog.sciencenet.cn/blog-38998-1459871.html
上一篇:色素色| 在绿色画布上色素描绘花的色彩
下一篇:格特鲁德·杰基尔|以花园边界色彩及轮替改变20世纪园艺方式
