博文
色素色| 在绿色画布上色素描绘花的色彩  精选
精选
|
【内容提要】开花植物的显著特征在于其花色彩鲜艳,而花色彩主要是由花器官细胞内所含色素的积累形成的。花呈现的非凡色彩主要依赖于四类色素:叶绿素、类胡萝卜素、花青素和甜菜碱。类黄酮拥有数千种不同的化合物,是种类最多、分布最广的色素群;包括有色花青素、橙黄酮和查耳酮,以及许多类黄酮化合物,如黄酮和黄酮醇,人类看不见这些化合物,但大多数传粉昆虫却能看见,因为这些化合物能吸收紫外线。花色可能只由一种色素产生同色系,也可能由几种色素在相同或不同的花器官中积累而形成极其复杂的色彩图案。
引言
花的色彩彻底改变了艺术、历史和科学。从克劳德·莫奈(Claude Monet, 1840-1926)在吉维尼(Giverny)以色彩为灵感绘画,到维多利亚时代的花语(Language of Flowers),花色被用来传递爱情和效忠的隐秘信息,花瓣的调色在欧洲历史的许多方面都发挥了有影响力的作用。在科学领域,孟德尔用花色揭示遗传规律,赖特用花色作为群体遗传学理论发展的实证案例,mRNA 分子引起的花色变化使紫色牵牛花变为白色。
克劳德·莫奈(1840~1926 年),法国画家,印象派风格的开创者、领导者和坚定不移的倡导者。在莫奈的成熟期作品中,他发展了对同一主题进行系列重复研究的方法,并随着光线或兴趣的变化更换画布。这些系列作品经常成组展出--例如,他的麦垛(1890/91 年;常被称为干草堆)和鲁昂大教堂(1894 年)。莫奈在吉维尼的家中创作了睡莲池塘,这也是他最后一系列画作的灵感来源。20 世纪下半叶,莫奈的知名度急剧上升,他的作品在世界各地的博物馆展出,吸引了破纪录的参观者,以他的艺术形象为主题的商业产品也广受欢迎。


人类对花色的迷恋源于底层色素的多样性驱动的。花的四轮结构都可以着色,从花萼到雌蕊。尽管有色花蜜和有色花粉是植物学中的新奇事物,但花瓣上的彩绘才是吸引最多注意力的。花瓣的颜色种类繁多,与典型的绿色背景形成鲜明对比,这是植物邀请传粉者来访的诚实信号,也是大多数开花植物繁衍生息的关键步骤。
颜色感知存在于观察者的思维,是花瓣反射的光波长、接受者的视觉系统和特定光环境背景的产物,是距离、亮度、花瓣表面形态和视觉背景共同作用的综合效果。人类只能感知电磁波谱中大约 400~700 nm 范围的可见光。在这个范围内,人们区分了从紫色到红色的一系列颜色。虽然花瓣细胞的形状可能会改变反射光的波长,从而改变感知的颜色(即结构色),但花的颜色主要由各种色素的类型和数量控制,这就是为什么花色主要被认为是色素性的原因。色素是一种化合物,当暴露在光线下时,可以吸收某些波长并反射其余波长。最终,色素吸收的特定波长取决于其化学结构。
仅四类色素就能产生非凡的花色多样性
在开花植物中发现的大多数色素属于四大类:叶绿素、类胡萝卜素、类黄酮和甜菜碱。众所周知,叶绿素(chlorophylls)是主要光合色素,吸收蓝色和红色波长的光,在花中相对罕见,主要局限于少数类群,例如马兜铃属(Aristolochia)、大戟属(Euphorbia)、贝母属(Fritillaria)、常春藤属(Hedera)和蜂兰属(Ophrys)的某些物种。第二,类胡萝卜素(carotenoids)在开花植物中无处不在,属于类异戊二烯类的有机色素,主要吸收蓝色波长的光,具有典型的三峰吸收光谱。类胡萝卜素可将花涂染成黄色到橙色,很少红色,是菊科和豆科许多黄色花物种的主要花瓣色素。第三,最多样化的色素是类黄酮(flavonoids),这种水溶性多酚几乎在所有维管植物都有发现。花青素(Anthocyanins)主要吸收绿色波长的光,反射紫色、蓝色、粉红色和红色。其他类黄酮,如儿茶素(catechins)、黄酮醇(flavonols)、黄酮(flavones)、异黄酮(isoflavones)和黄酮(flavanones),只在紫外光谱部分吸收;因此,这些“吸收紫外线的类黄酮”对人类是不可见的,但许多传粉者如蜜蜂、苍蝇、蝴蝶和大多数鸟类都能分辨其色彩。类黄酮和查尔酮,以及一些糖基化黄酮醇,在紫外-蓝光谱范围内吸收,产生黄褐色到黄色。第四,甜菜碱(betalains)是含氮化合物,可产生黄色至粉红色和红色,仅存在于石竹目的某些科,例如仙人掌科(Cactaceae)和紫茉莉科紫茉莉(Mirabilis jalapa)。有趣的是,产生甜菜碱的物种并不产生花青素(anthocyanins);除其他因素外,这两种色素的相互排斥可能是由于对共同底物的竞争引起的。除了这些主要的色素群外,还有一些在野生植物中极为罕见的化合物,例如菊科红花(Carthamus tinctorius)中的醌查尔酮,某些鸢尾属和龙胆属某些物种中的克山酮。
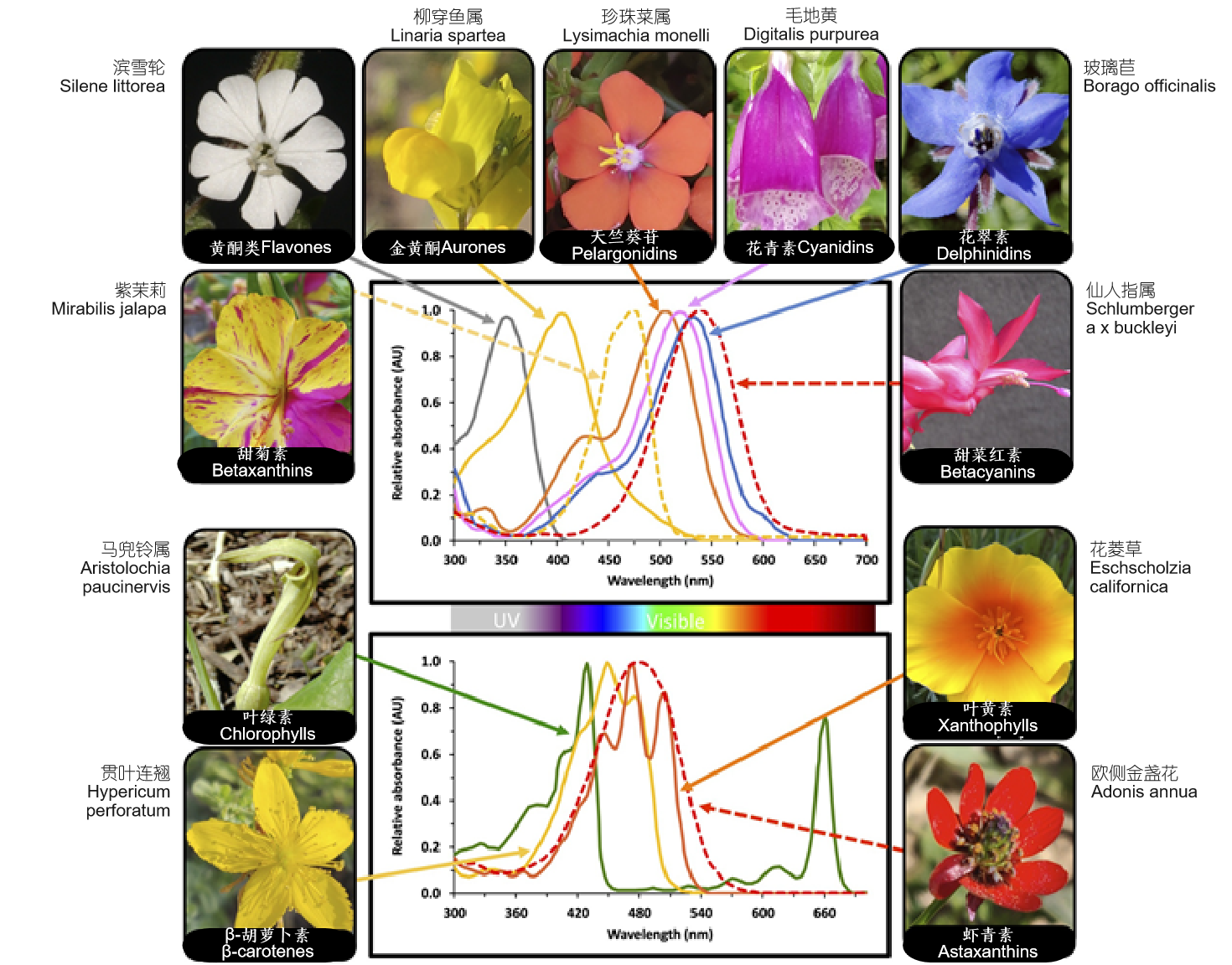
主要花色素的吸收光谱和含有这些色素的花卉案例
花中的色素:单独的、混合的还是分层?
色素的特定类型或各种色素的混合比例的变化会影响花最终的颜色(即色调),而化合物浓度的变化主要影响颜色强度(即颜色饱和度或纯度)。花的颜色可从简单的单色到极其复杂的颜色图案,就像兰花、鸢尾、马利筋或鱼尾草中经常出现。通过精确地调节花瓣不同部位(例如斑眼、条纹、镶边)的色素合成,可以产生复杂的颜色图案。

(a)兰科叶蜂兰(Ophrys tenthredinifera),(b)鸢尾科阴阳兰(Moraea sisyrinchium)和(c)半日花科松露花(Tuberaria guttata)。字母表示按丰度顺序列出的各花器官积累的主要花色素群。ant-花青素;fla-吸收紫外线的黄酮类化合物;chl-叶绿素; car-类胡萝卜素。
相似的花色可以通过独特类型的色素积累或通过多种色素的组合而产生。例如,花瓣中的红色可能是由于独特的红色花青素、红色胡萝卜素或红色类胡萝卜素的存在,或者是由于紫色花青素与橙色类胡萝卜素的组合。色素通常位于花瓣解剖结构的表皮层和叶肉层。由于不同细胞器中的积累,同一细胞中不同色素类型的组合可能会发生。类胡萝卜素和叶绿素是脂溶性分子,分别储存在花瓣细胞的染色质体和叶绿体中。类黄酮和甜菜碱是水溶性分子,主要在液泡中积累;因此,花青素和其他吸收紫外线的类黄酮可以混合在同一个液泡中。由于没有彩色色素,白色花瓣通常反射所有波长的可见光;然而,这些花瓣通常含有高浓度的吸收紫外线的类黄酮,例如黄酮或黄酮醇。同样,花蜜指南(即引导传粉者获得奖励的空间线索)通常对人类不可见,但对某些昆虫可见,因为它们是由吸收紫外线的黄酮醇和黄酮的积累引起的。
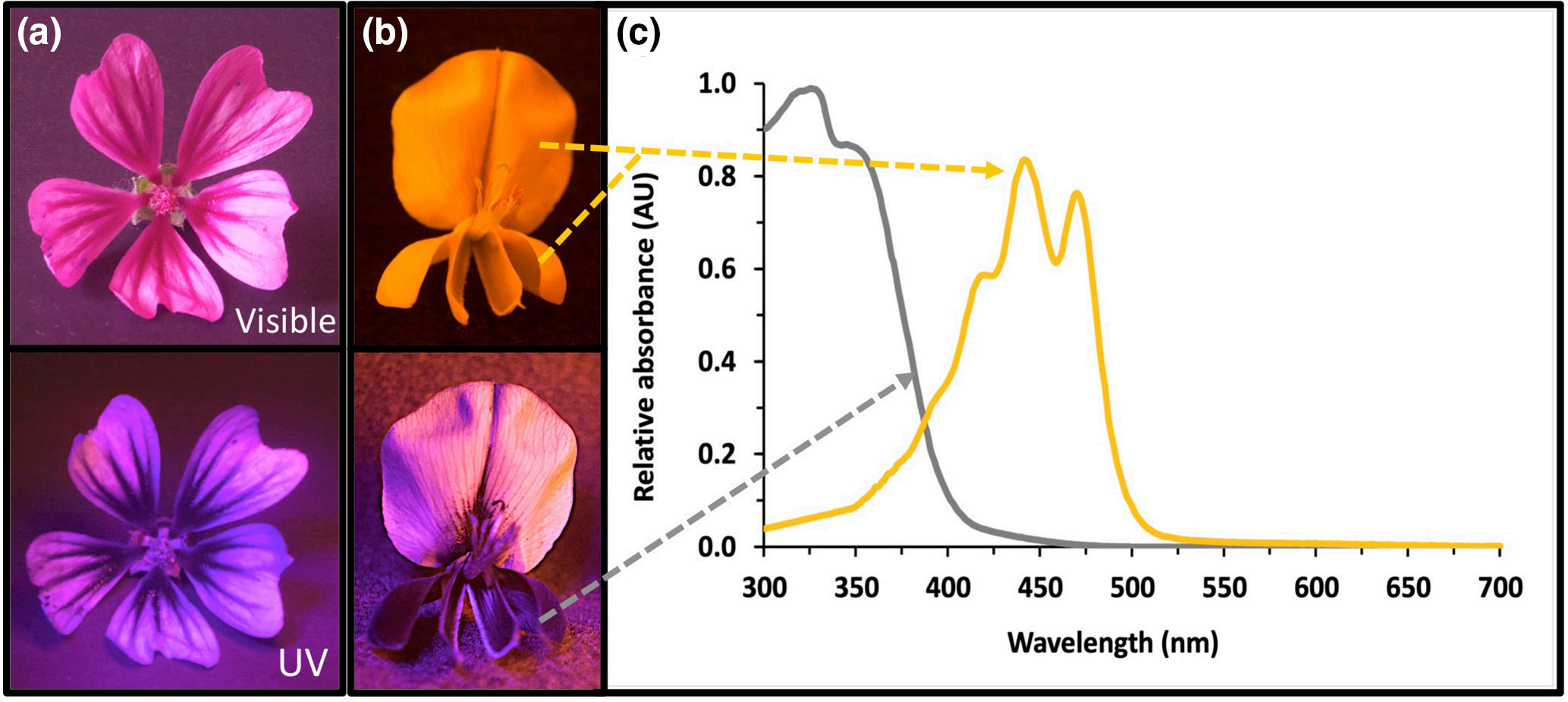
(a)锦葵科锦葵(Malva cathayensis),花瓣上有人类可见的深粉色放射线;在紫外线摄影中,由于存在吸收紫外线的类黄酮,放射线颜色更深。(b)豆科 鹰爪豆(Spartium junceum)的所有花瓣都显示出黄色类胡萝卜素;在紫外线摄影下,旗瓣可反射紫外线,而龙骨瓣和翅瓣可吸收紫外线,这是因为含有类黄酮而吸收紫外线。(c)旗瓣(黄线)和翅膀(灰线)提取物的吸光度光谱。
花青素作为研究花色的模型
花青素(Anthocyanins)一词,来自希腊语的 anthos(花)和 kyanos(蓝色),无疑是提供最多种类花色调的色素。花青素可以产生可见光谱中的大多数颜色,包括一些最稀有的颜色,如绿松石色,例如,高山龙舌凤梨(Puya alpestris)或黑色(例如,角锹兰属Streptanthus niger)。花青素是花青素的糖基化形式,花青素是核心色素结构。维管植物中描述了 600 多种花青素,其中大多数来源于六种类型的花青素:矢车菊素、飞燕草素、天竺葵素、芍药素、矮牵牛素和锦葵素。大多数花青素通过添加各种糖基化和酰化进行后续的化学修饰。此外,金属或其他类黄酮(如黄酮和黄酮醇)可以充当共色素(与花青素结合以稳定和增强其颜色的弱着色物质),进一步增加颜色的多样性——通常会增强着色。而且,如果这还不够修饰,花青素的颜色取决于溶液的 pH 值;在酸性条件下,占主导地位的黄酮阳离子呈红色,但在较高的 pH 值下,由于喹啉类形式,颜色变为蓝色。因此,细胞的 pH 值至少在理论上可以改变花青素的化学结构,从而产生颜色的剧烈变化。最后,光线进入并反射回观察者的表皮细胞形状会进一步影响着色。因此,由各种花青素类型与多层装饰以及颜料所在的物理和化学环境相结合,创造了几乎无限的调色板。
花朵不断变化的颜色
大多数被子植物物种都表现出独特的花色,无论环境如何,这种花色在个体内部和个体之间都是稳定的。然而,有时同一物种的花色会随着时间的推移而改变颜色。因此,花瓣的颜色可能会随着花的生命周期而改变,变得更深甚至改变颜色。例如,由于花青素降解的活跃过程,大花鸳鸯茉莉(Brunfelsia pauciflora)的花(昨天、今天和明天)从深紫色变为纯白色。在一些研究案例中,这种颜色变化可能与传粉媒介的吸引力有关(在许多红木森林草本植物(如延龄草)授粉后白色花瓣呈粉红色)。十字花科堇娘芥(Moricandia arvensis)在春天开出反射紫外线的淡紫色花,但到夏天则转为吸收紫外线的白色花朵,而不是单朵花变色。

(a)大花鸳鸯茉莉(Brunfelsia pauciflora)(茄科Solanaceae)在花的生命周期中会改变花瓣的颜色,从深紫色到淡紫色再到纯白色(昨天、今天和明天)。(b)萝卜(Raphanus sativus)(十字花科Brassicaeae) 表现出花瓣颜色多态性,植物根据花青素和类胡萝卜素的独立存在(+) 或不存在(–) 而产生四种可能类型的花中的一种。
另一种现象是种群内个体之间或种群间的花色变化。这种变化可能是离散的,产生两个或多个不同的颜色变化(又名花色多态性,如 Lysimachia arvensis),或者变化可能更像是连续的渐变(如 Silene littorea 和 Parrya nudicaulis)。在色素生产方面,花色多态性可能是由色素浓度的变化引起的,导致花色强度的梯度或完全缺乏色素(例如,Silene littorea 的深粉色到浅粉色再到白色花瓣)。花色多态性也可能是由多种色素类型的层变化引起的,从而产生不同颜色的花朵。例如,在 Raphanus sativus 中,花瓣的颜色取决于花青素和类胡萝卜素的存在与否。由于每个色素组都是独立控制的,花青素和类胡萝卜素的混合和匹配会产生四种主要的颜色变化:青铜色(花青素和类胡萝卜素)、粉红色(只有花青素)、黄色(只有类胡萝卜素)和白色(不含两种色素)。有趣的是,并非所有类型的花色多态性都同样频繁。在伊比利亚半岛和不列颠群岛的植物区系中,涉及色素损失(即白色)的花色多态性比色素类型变化引起的花色多态性更常见。

牛顿用棱镜准确地展示了什么是颜色

牛顿试验棱镜。根据 J.A. Houston 的一幅画雕刻,约 1870 年

发光蘑菇邮票



众神之食 ,露丝·皮特,自《北欧众神的悲剧》,1893 年
延伸阅读:
Narbona E, del Valle JC, Whittall JB (2021) Painting the green canvas: how pigments produce flower colours. Biochem (Lond) 43 (3): 6–12.
https://blog.sciencenet.cn/blog-38998-1459367.html
上一篇:结构色 | 花瓣表皮结构对南非植物花色的贡献
下一篇:花色的演变|从色彩缤纷的球果最初演变至人类世演变
