博文
意识是什么?----第二章 信息流(2)
||

2.3 生物细胞内的信息流
2.3.1 酶介导的信息流
生命体与非生命体统一于物理层面,分道与化学层面。这里的物理层面是指元素水平上的差异,而化学层面主要体现在组成生物体的一些化学物质(如酶)会具有一定的活性,其结构会出现一定程度的变动,表现为一种构象时具有一定的活性,而另一种构象时却表现为另一种活性状态。
能流进化论认为,细胞的基本组成单位是由酶作为能量传递介体的能量传递单元,即“能量供体→酶→能量受体”。细胞内的能量传递单元相互影响,相互制约,驱动了能量供体中的电子向能量受体方向传递,同时将能量供体中的部分化学能转化为热能。
在一个特定细胞中,物质分子的布朗运动,以及不同种类物质分子之间的相互作用,使得特定微环境中以酶作为能量传递介体的能量供体和能量受体的浓度时刻发生着变化,而且这种变化具有随机性,进一步导致特定微环境中“能量供体→酶→能量受体”这一能量传递单元的不稳定性,进而导致所属体系结构和功能的不稳定。
细胞生物学的发展已经表明,生物细胞的结构和功能是相对稳定的,这就说明,一个结构和功能不稳定的非生命体系是不可能演化成细胞生命体的。反过来讲,细胞生命体的出现表明,特定的酶与其能量供体和能量受体组成的能量传递单元是相对稳定的,这便要求该酶具有能及时、快速的识别其能量供体和能量受体的能力,即酶与其能量供体和能量受体之间的信息交流具有特异性和快速识别的特性。在能量流的驱动下,酶逐渐演变为只识别特定能量供体和能量受体的能量传递介体。
那么,酶与其能量供体和能量受体是如何进行信息交流的呢?又是如何保障特异性和高效性的呢?以下将从酶的结构和分布两个方面来解析:
1)结构方面的策略
酶是具有催化活性的蛋白质,其结构组成主要是氨基酸。组成蛋白质的氨基酸都是电中性的,在电性方面是相对稳定的,但其也包含着两个相对不稳定的因素:一个是容易失去电子的氨基,一个是容易获取电子的羧基。那么,为什么氨基酸会有这两个相对不稳定的共性?
氨基和羧基让不同的氨基酸可以相互连接,即一个氨基酸的氨基与另一个氨基酸的羧基通过脱水缩合形成肽键。在电荷分布上,肽键令一个氨基酸的氨基和另一个氨基酸的羧基相对单独的氨基和羧基而言更加稳定。尽管如此,新合成的产物仍然存在一个电荷相对不稳定的氨基和羧基。然而,就电荷分布的稳定性而言,相互连接的氨基酸越多,不稳定部分所占的比例就越低,进而使得整个产物的电荷稳定性增加。物质的电子分布越稳定,就越适合自然界对组成物质电子稳定性的需求,就有存在的必要,也就有出现的可能。
可以把酶看成是一根一头接水龙头,一头接水桶的水管。组成酶的每一个氨基酸相当于一根水管,氨基酸通过肽键连接而成的多肽链可以看作是由不同的水管连接而成的新的水管。如果一直是线性的排列,随着多肽链长度的增加,电子的分布受环境的影响就越大,不稳定性也就随之增加,这必然不符合物质对稳定性的需求。也就是说,如果氨基酸是线性排列,随着多肽链的延长,多肽链上的电子分布就会发生从不稳定到相对稳定再到不稳定的变化。物质对稳定性的需求决定了多肽链不可能以线性的形式存在。事实上,多肽链只是酶的一级结构,在其合成过程中,多肽链会通过α-螺旋、ꞵ-折叠、ꞵ-转角、无规则卷曲等形式变成二级结构,该二级结构进一步折叠变成球形、椭球形等,这些变化的结果都是令酶分子在整体上实现电子分布的相对更好的稳定。
2)分布方面的策略
如果细胞内所有的酶和能量供体、能量受体等是随机分布的,那么,“能量供体→酶→能量受体”这一能量传递单元的不稳定性就会增加,这必然违背自然界在能量流方面的需求,是要被自然界所淘汰的。《隐藏的动力:生物在自然界中的价值》一书中的“生命起源”一章有说明,在自然界能量流方面需求的驱动下,不同的能量传递单元之间,特别是在存在“共享能量供体”、“共享能量受体”或者“共享能量传递介体”时,会向着最有利于能量传递的方向演变(注意,并不是不存在不利于能量传递的演变,而是这种演变在自然选择过程中被淘汰了)。伴随着这个过程,能量供体之间、能量受体之间、能量传递介体之间以及能量传递单元之间也出现了反映它们之间关系的两种信息,即“竞争”与“合作”。有利于能量传递的能量传递单元之间便是“合作”关系,同一功能位的能量传递单元之间便是“竞争”关系。事实上,细胞中的酶并不是随机分布的,它们被各种细胞器分组,提高了它们与能量供体和能流受体的接触几率。例如,真核细胞中参与三羧酸循环的酶均位于线粒体内,电子传递链上的酶则位于线粒体的内膜上,这种分布利于电子从能量供体乙酰辅酶A经NADH向O2方向传递。
能流进化论将处于同一个细胞体系中的物质按其在能量流中的作用分为两类:一类是中央能量控制系统;一类是辅助系统。中央能量控制系统主要负责能量的获取、传递、储存、分配等,是非生命态体系向生命体演变过程所依附的“地基”。该系统可有效提高一个体系的能量传递及利用效率,在介导能量供体中的能量进行释放和传递的同时,也维持着自身结构和功能的稳定。在细胞内,参与糖酵解途径、三羧酸循环、磷酸戊糖途径、电子传递链、脂肪酸代谢等过程的物质分子都属于这个系统,它们在能量的释放和传递过程中发挥着重要作用。
辅助系统的功能主要体现于抑制或削弱其竞争者的竞争能力,以及维持自身系统的稳定性或抵抗来自竞争者的攻击等。辅助系统让所属细胞体系的能量传递能力的稳定性和持续性得到保障,主要包括遗传信息、调控信息等。
综上所述,生物细胞内的信息按信息载体可分为:能量供体信息、能流受体信息、能量传递介体信息、辅助信息(如遗传信息、调控信息等)等。按物质或系统之间的关系可将细胞内的信息分为:竞争信息和合作信息。基于此,生物细胞内的信息流主要包括:1)能量流上的信息流,即“能量供体⇋能量传递介体⇋能量受体”;2)DNA、RNA和蛋白质之间的遗传信息流;3)调控因子与被调控因子之间的调控信息流;4)存在竞争或合作关系的物质分子或系统之间的信息流。
2.3.2基于能流和信息流的电子传递链构想
电子传递链是一系列电子载体按对电子亲和力逐渐升高的顺序组成的电子传递系统。所有组成成分都嵌合于线粒体内膜或叶绿体类囊体膜或其他生物膜中,而且按顺序分段组成分离的复合物,在复合物内各载体成分的物理排列也符合电子流动的方向。其中线粒体中的电子传递链是伴随着营养物质的氧化放能,又称作呼吸链。
1961年英国生物化学家米切尔(P. Mitchell)提出了电子传递的化学渗透假说。他认为电子传递链像一个质子泵,电子传递过程中所释放的能量,可促使质子由线粒体基质移位到线粒体内膜与外膜间空间形成质子电化学梯度,即线粒体内膜外侧的H+浓度大于内侧并蕴藏了能量。当电子传递时,质子(H+)被泵出到内膜外测,在H+浓度梯度的驱动下,H+通过FoF1 ATP酶中的特异的H+通道或“孔道”流动返回线粒体基质时,由H+流动返回所释放的自由能驱动FoF1 ATP酶催化ADP与Pi偶联生成ATP。此假说假设在电子传递驱动下,H+循环出、进线粒体,同时生成ATP,虽能解释氧化磷酸化过程的许多性质,但仍有许多问题未能完全阐明。
能流进化论认为,生物体以及组成生物体的细胞均是能量传递介体,其价值是介导食物(能量供体)中的能量流向能量受体(也称电子受体,指环境中用以接收细胞代谢过程产生的流经电子传递链的电子的氧化性物质,如氧气),以驱动自然界能量分布的相对均衡。由于能量供体在细胞内的代谢过程是由一系列的氧化还原反应组成,而氧化还原反应的本质是电子的得失与转移,因而能量进化论还认为,细胞的功能单位是由提供电子的还原性物质、介导电子传递的催化酶和接收电子的氧化性物质组成的能量传递单元。总之,能流进化论认为,能量供体中的部分电子经组成细胞的一系列的能量传递单元传递给能量受体,在这个过程中,在自然条件下难以释放和传递的化学能被转化为相对更容易在自然界释放和传递的热能。基于此,作者认为电子传递链在化学渗透假说的基础上做如下修订,可能更合适。
原假说观点1. 线粒体内膜上的呼吸链同时起质子泵的作用,可以在传递电子的同时将质子从线粒体基质腔转移到膜间腔。
新构想认为线粒体内膜上的呼吸链同时起质子泵的作用,在泵出的同时,促进了电子向下游复合体的流动。区别在于,原假说认为电子的传递是质子泵出的原因,而新构想认为电子的传递是质子泵出的结果。
我的观点:呼吸链上的还原酶复合体具有两个功能:一是辅助底物脱氢;二是将脱下的H+转移到内膜外侧。还原酶的脱氢功能众所周知,不多解释。先探讨两个问题:1)电子传递链为什么需要脱氢酶,为什么需要脱氢?2)H+是如何转移到线粒体内膜外侧的,是什么力的作用驱动了这一过程?
如果将葡萄糖、细胞和氧气看成是一个系统,在细胞呼吸作用的影响下,葡萄糖被转化为水和二氧化碳。从能流的角度看,细胞的呼吸作用可以看作是葡萄糖中的电子经细胞传递给氧气,细胞就是一个电子传递介体。既然如此,细胞内参与呼吸作用的所有的化学反应的主要矛盾就应该是驱动电子的流动,电子传递链自然也不例外,即发生在电子传递链上的生化反应的主要目的就是驱动电子的流动。明确了这个关系,那么,在质子与电子的关系中,就不应该有所争议,即质子的泵出是为电子在电子传递链上的传递服务的,而不是化学渗透假说中所说的电子的传递是为了质子的泵出。
正如河道与水流的关系,河道是为水流服务的,水也会选择合适的河道传递。如果河道堵塞,其介导水流的能力降低,那么水就会选择其它更合适的河道。电子的传递也是如此,它们也会选择更合适的电子传递通道,例如,当在硫还原地杆菌的培养液里同时加入延胡索酸和水铁矿两种电子受体时,在电子传递链上传递的电子就有两种选择:一是传递给延胡索酸;一是传递给水铁矿。延胡索酸可在细胞内接收电子,而水铁矿只能在细胞外接收电子,因而相比较而言,电子应该优先选择延胡索酸,实验结果也是如预期所示。当利用抑制剂如鱼藤酮、安密妥、杀粉蝶菌素等阻断电子传递链时,细胞就需要其它的电子传递途径,如果不存在这样的途径,那细胞最终只能走向死亡。
再者,我们知道化学反应本质上是电子的得失或转移,提供电子的化合物我们称之为电子供体,接收电子的化合物我们称之为电子受体。电子得失或转移的前提是两个化合物之间存在电势差(ΔE)。换句话说,如果两个反应物之间存在足够的电势差,那么电子供体中的电子就有传递至电子受体的可能。我们知道,很多化学反应需要在高温、高压等条件下才会发生,对于整个系统相对稳定的生物体而言,需要特殊条件的化学反应是不允许的。换言之,生物体内发生氧化还原反应的两个化合物之间,电子应该相对比较容易的发生转移。然而,如果两个化合物之间的电子转移太容易发生,那么细胞内整体电子的流速必然很快,进而导致能量难以及时的利用,势必造成能量的浪费。能量进化论认为,生物体在介导能量流动的同时,还需要从中获取维持自身能量传递能力的能量。这便出现了一对矛盾,即电子的转移既要相对比较容易的发生,又要控制其发生的速度,以利于生物从中获取维持自身能量传递能力的能量。那么,生物体是如何来解决这对矛盾的呢?
生物体的策略(作者的观点):
假设电子供体为A,电子受体为B,二者电势差为ΔE。在ΔE条件下,电子不能从A转移至B。这个时候,脱氢酶出现了,它先将A的一个H+脱去,导致A带负电,进而增加了A与B之间的电势差,变为ΔE’,此时,A携带的电子就相对比较容易的转移给B 了。因此,我认为脱氢酶是控制两个化合物之间电子流动的开关,其通过对电子供体的脱氢,改变电子供体与电子受体之间的电势差,保证了电子从电子供体往下游电子受体传递的可能。
既然两个化合物之间存在合适的电势差是它们之间能发生电子传递的前提,那么,通过脱氢酶增加两个化合物之间电势差就应该是化合物之间发生电子传递的原因。
对于前述第二个问题,H+是如何转移到线粒体内膜外侧的?是什么力的作用驱动了这一过程?我的观点是,脱氢酶具有两种功能,一是夺氢功能,即破坏底物(电子供体)中H+的稳定性,使其与自身的亲和力更强;二是递氢功能,即把底物中稳定性较差的H+传递给天然受体,如烟酰胺腺嘌呤二核苷酸(NAD+)、烟酰胺腺嘌呤二核苷酸磷酸(NADP+)、细胞色素等。此外,当某一脱氢酶在一特定环境中不存在天然受体时,该脱氢酶会与另外一种有天然受体的脱氢酶组成超脱氢酶复合体。已有研究证明,电子传递链上的脱氢酶复合体不是独立存在的,而是组成一个超脱氢酶复合体,通过结构解析,也发现这些脱氢酶复合体之间存在结合位点(Guo, R., et al., 2017. Architecture of human mitochondrial respiratory megacomplex I2III2IV2. Cell 170, 1247–1257)。
基于以上猜想,当脱氢酶与底物(电子供体)结合时,电子供体中的H+与脱氢酶有相对更高的亲和度,在H+偏向脱氢酶的瞬间,电子供体和电子受体之间利于电子传递的电势差形成。由于电子的转移速度要远远快于H+在脱氢酶和电子供体之间的转移速度,电子一旦转移至电子受体,根据化合物电荷守衡原理,电子供体与H+之间的结合力会进一步减弱。在这种情况下,被脱氢酶夺走的H+将难以重新回到电子供体。对于脱氢酶而言,虽然其对H+具有一定的亲和力,然而,为了维持自身的电荷守恒,其不可能接受这个H+,结果便是, H+以游离态存在。
游离态的H+在脱氢酶复合体上就存在两个方向上的移动可能:一是留在线粒体基质,一是流入线粒体内膜与外膜之间的间隙。大量直接或间接的实验证明,膜表面能够滞留大量H+,并且在一定条件下质子能够沿膜表面迅速转移,表明H+在流动方向的选择中选择了后者。那么,是什么力推动了H+选择后者的呢?这个问题会在后面回答。
原假说观点2. 线粒体内膜上的ATP合酶复合体也能可逆地跨线粒体内膜运送质子,一方面利用水解ATP的能量将质子从基质腔转移到膜间腔,另一方面当膜间腔存在大量质子使线粒体内膜内外存在足够的电化学H+梯度时,质子则从膜间腔通过ATP合成酶复合物上的质子通道进入基质,同时驱动ATP合成酶合成ATP。
新构想认为,呼吸链上的各种脱氢酶复合体以及ATP合酶复合体,均具有质子交换膜的性质,既可允许质子泵出,又可允许质子泵入。对于ATP的合成,是电子经ATP合酶流出所产生的电能转化为储存在ATP中化学能的过程,而不是由H+因内膜两侧的浓度差和电势差,经ATP合酶流入线粒体基质过程所产生的动能转化为储存在ATP中化学能的过程。线粒体内膜两侧的H+梯度为电子的流动提供了可能,氧气是在内膜外侧接收电子和H+结合生成H2O,氧气的存在直接驱动了质子的流动,间接驱动了电子的流动。此外,新构想认为,ATP合酶的Fo复合体相当于一台电动机,电子在其中流动的过程也就是电能转化为这台电动机机械能的过程。F1复合体是合成和水解ATP的区域,在这里Fo复合体产生机械能进一步转化为储存在ATP中的化学能。这个过程就像水力发电站与电视机之间的逻辑关系:一种逻辑是将水的动能转化为电能,有了电能,电视机才能运行(类比H+的泵出驱动电子的流动,电流的电能经ATP合酶转化为储存在ATP中的化学能);另一种逻辑是,电能转化为水的动能,有了水的动能,电视机才能运行(类比原假说中的电子的流动驱动H+的泵出,H+在ATP合酶中流动时的动能转化为储存在ATP中的化学能)。看看我们日常家电的运行,我觉得没有人会认可第二种逻辑。
再者,原假说认为,是H+通过Fo复合体时的动能转化为储存在ATP中化学能。如果H+经ATP合酶流入线粒体基质过程所产生的动能可以转化为储存在ATP中化学能,那么,H+被脱氢酶复合体泵出时的动能为什么就不能直接驱动ATP的合成?或者说,为什么是“质子泵→H+流出质子泵时的动能→H+流入ATP合酶时的动能→ATP”,而不是“质子泵→H+流出质子泵时的动能→ATP”?况且,在ATP合酶复合体区域,还存在一个因H+浓度差而形成的电势差,为质子通过Fo复合体也提供了驱动力。在前面质子的泵出过程缺少这个电势差的辅助,这便意味着最初质子从线粒体基质泵入内膜和外膜间隙时,需要耗费更多的能量。原假说的情况就变成,细胞耗费更多的能量就是为了合成一小部分储存在ATP里的能量。可以想象为这样一种场景,一个人利用抽水机将地面的水抽到高出,然后再利用一台机器为抽到高出的水流向地面时提供一个驱动力,使水留下时的动能经发电机产生电能。这于情于理都说不过去,更何况细胞内的各种调控是如此精细,怎么可能会出现这种浪费能量现象?
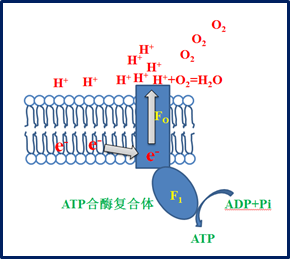
如果按照我提出的氧气是在内膜外侧接收电子和H+结合生成H2O,问题就解决了。氧气在内膜外侧接收电子,同时不断的消耗H+, H+消耗的同时也为脱氢酶脱氢过程产生的自由H+往内膜外侧的流动提供了牵引力,正是该牵引力的存在, 使得H+在选择继续留在线粒体基质中还是流入内膜外侧的选择中选择了往内膜外侧流动(这也回答了前面预留的问题)。
新构想有几个优点:
1)减少了水和氧气在线粒体内膜上的进出。既然电子传递链的最终产物是水,如果是在线粒体基质中合成水,为了维持最终的渗透平衡,合成的水最终还是要排出来,既然如此,为何当初就不能直接在内膜外侧合成呢?虽然水和氧气是膜通透性的,但减少在内膜上的进出更利于生物体的渗透平衡。
2)进一步解释了生物体为什么需要水。我们都知道生物体的存活需要水,一个原因就是因为生物体内的很多代谢反应需要水的存在和参与,在新构想中,生物体内水的流动是内膜外侧H+流动的原动力,同时,电子传递链上生成的水也为水在生物体中的流动起到了推动作用。
对于耗氧生物而言,氧气是最终的电子受体,然而对与很多厌氧生物而言,它们所需要的很多电子受体是难溶性的金属氧化物,这些金属氧化物颗粒比较大,很难进入细胞内,这就需要生物体将电子传递到细胞外,交给外面的电子受体,这个过程就是胞外电子传递,也称作胞外呼吸,具有胞外呼吸特性的微生物成为胞外呼吸菌。如果按原假说,是质子浓度梯度,即质子在ATP合酶中的流动是ATP合成的原因。这便出现一对矛盾,电子往外胞外传递,质子却往胞内传递,那么最终结果是微生物带正电了,这也违背了电荷守恒原则。
https://blog.sciencenet.cn/blog-3479210-1369265.html
上一篇:意识是什么?----第二章 信息流(1)
下一篇:意识是什么?----第三章 生命体介导的信息流的演变(1)
全部作者的精选博文
- • 再谈科研思路
- • 科研的思路应该是什么?
- • 面点统一的高等教育培养模式
- • 我眼中的王老师