ВЉЮФ
Й§УєЬивьадBМЧвфЯИАћ  ОЋбЁ
ОЋбЁ
||
ЖдЙ§УєдЕФУтвпЗДгІЕФЬиеїЪЧ 2 аЭБэаЭКЭ T ИЈжњад 2 ЃЈTH2ЃЉ ЗжУкАзЯИАћНщЫи-4 ЃЈIL-4ЃЉЁЂIL-5 КЭ IL-13 ЕФЯИАћЁЃетаЉЯИАћвђзгМЄЛюЙ§УєдЬивьад B ЯИАћЃЌгеЕМЫќУЧЗжЛЏЮЊВњЩњМЧвф B ЯИАћ ЃЈBmemЃЉ КЭУтвпЧђЕААз E ЃЈIgEЃЉ ЕФНЌЯИАћЁЃМЧвф T ЯИАћКЭОпгаЙ§УєдЬивьадЕФ Bmem ЕФДцдкЪЙЕУдкЕкЖўДЮгіЕНЙ§УєдЪБМЬајЁАЙ§УєадЁБ2 аЭУтвпЗДгІЁЃетаЉУтвпЪТМўЕФЙиМќЪЧДцдкЖдЙ§УєдГЩЗжОпгаЬивьадЕФ IgEЁЃIgE гыЗЪДѓЯИАћКЭЪШМюадСЃЯИАћБэДяЕФИпЧзКЭСІ IgE ЪмЬх FcІХRI НсКЯЁЃЫцКѓЕФЙ§УєдБЉТЖгеЕМНсКЯЕФ IgE НЛСЊЃЌДЅЗЂЗЪДѓЯИАћКЭЪШМюадСЃЯИАћЭбПХСЃЃЌЪЭЗХАќРЈзщАЗдкФкЕФбзжЂНщжЪЁЃШЛЖјЃЌЗжУкаЭ IgE ЕФРДдДЩаВЛЧхГўЃЌвђЮЊ IgE Bmem МЋЮЊКБМћЃЌетПЩФмЪЧгЩгкЗвыЧЗМбЕМжТЕФ IgE БэУцБэДяЕЭЁЃдкБОЦкЁЖПЦбЇзЊЛЏвНбЇЁЗжаЃЌСНЯюбаОПБЈИцСЫвЛжж 2 аЭМЋЛЏ Bmem бЧШКЃЌИУбЧШКгаЭћзЊЛЛЮЊ IgEЃЌДгЖјИГгшЖдЖржжЙ§УєдЙ§УєЕФИіЬхЙ§УєдЬивьад ЁЃЪЙгУВЛЭЌЕФЛМепЖгСаКЭММЪѕЃЌOta ЕШКЭ Koenig ЕШСНепЖМЗЂЯжСЫетжж2аЭBmemШКЬхЃЌЦфЬиеїЪЧIgGЁЂCD23ЃЈЕЭЧзКЭСІIgEЪмЬхЃЌFcІХRIIЃЉЁЂIL-4ЪмЬхЃЈIL-4RЃЉКЭжжЯЕIGHEЕФБэДяЁЃ
дкЫћУЧЕФбаОПжаЃЌOtaЕШШЫМьВщСЫвЛзщЛЈЩњЙ§УєЖљЭЏЕФбЛЗBmemЁЃзїепЗЂЯжЃЌдкбЊЧхЛЈЩњЬивьадIgEХЈЖШИпЕФЖљЭЏжаЃЌCD23 BmemЦЕТЪдіМгЃЈ>100 kU+вЛИі/Щ§ЃЉгыЛЈЩњЬивьад IgE ЕЭ ЃЈ<5 kUвЛИі/Щ§ЃЉКЭЗЧЙ§УєИіЬхЁЃCD27 B ЯИАћЕФЕЅЯИАћ RNA ВтађЗжЮіНвЪОСЫЫљЮНЕФЁА2 аЭБъМЧЁБIgG1 Bmem ШКЬхЃЌЦф IL4RЁЂFCER2ЃЈБрТы CD23ЃЉКЭжжЯЕ IGHE ЕФЛљвђБэДядіМгЃЌвдМАИпЖШЭЛБфЕФ B ЯИАћЪмЬхЁЃдкЛЈЩњЬивьад IgE ИпЕФЛЈЩњЙ§УєЖљЭЏжаЃЌдкЛЈЩњЙ§Уєд Ara h 2 Ьивьад B ЯИАћжаМјЖЈГіЪеСВ Ig ЛљвђађСаЃЌетаЉ Ig ађСаРДдДгк 2 аЭ Bmem ШКЬхЁЃЫћУЧЕУГіЕФНсТлЪЧЃЌзЊТМжжЯЕIGHEЕФ2аЭBmemЯИАћАќРЈвЛИіЖРЬиЕФШКЬхЃЌИУШКЬхдкЛЈЩњЙ§УєЖљЭЏжаИЛМЏЃЌВЂгаПЩФмЗжЛЏЮЊВњЩњИпЧзКЭСІЕФIgEНЌЯИАћЁЃ
KoenigЕШШЫЕФБЈИцдкЙ§УєКЭЗЧЙ§УєИіЬхжаМјЖЈГі 2 аЭМЋЛЏ Bmem ШКЬхЃЌжжЯЕ IGHEЁЂFCER2ЁЂIL4R ЕФЛљвђзЊТМдіМгЃЌCD32 ЕФзЊТМIL13RA1ЕЋзЊТМНЯЕЭЁЃдкЫћУЧЕФЖгСажаЃЌЖдшыЪїЛЈЗлЛђЪвФкГОђ§Й§УєЕФГЩФъШЫБШЗЧЙ§УєИіЬхОпгаИќЖрЕФбЛЗIL-4RІСCD23 2аЭBmemЁЃетаЉ Bmem ИЛМЏСЫБэДя IgG1 КЭ IgG4 ЕФЯИАћЃЌВЂаЏДјИпЖШЖрбљЛЏЕФ Ig ЛљвђПтЃЌЕЋУЛгажЄОнБэУї V ЛљвђЪЙгУДцдкЧїЭЌЛђЦЋваЁЃЫфШЛЙ§УєдЬивьад Bmem гГЩфЕН 2 аЭ Bmem ШЫШКЃЌЕЋРДздЭЌвЛЙЉЬхЕФбЯжиМБадКєЮќЯЕЭГзлКЯжЂЙкзДВЁЖО 2 ЃЈSARS-CoV-2ЃЉ ПЙдЬивьад BmemЃЈДњБэ 1 аЭУтвпСІЃЉУЛгаЃЌетБэУї 2 аЭБэаЭгыЙ§УєадМВВЁЯрЙиЁЃДЫЭтЃЌ2 аЭ Bmem дкЩрЯТУтвпжЮСЦ ЃЈSLITЃЉ 1 ИідТКѓдкЬхФкЗжЛЏЮЊВњЩњ IgE ЕФНЌЯИАћЃЌЦфжаЙ§УєдГЃЙцдкЩрЯТИјвЉЃЌФПЕФЪЧМѕЩйЙ§УєЗДгІЃЌШч SLIT ЧА 2 аЭ Bmem гы SLIT Кѓ IgE жЎМфЕФПЫТЁЙиЯЕЫљЪОЁЃзїепЛЙБэУїЃЌдкВЛЭЌЭООЖЕФЙ§УєжТУєКѓЃЌаЁЪѓФЃаЭжагеЕМСЫЙ§УєдЬивьад 2 аЭ BmemЁЃЫфШЛВЛашвЊЩњЗЂжааФКЭбЛЗ IgEЃЌЕЋ IL-4 ЖдгкгеЕМ 2 аЭБэаЭжСЙиживЊЁЃетаЉЪ§ОнБэУїЃЌ2 аЭ Bmem ЪЧЙ§УєЕФжТВЁШЫШККЭ IgE ЕФЫожїЁЃ
етаЉЗЂЯжВњЩњЕФвЛИіживЊЮЪЬтЪЧЃЌ2аЭBmemЪЧШчКЮаЮГЩЕФЃЈЭМ1ЃЉЁЃKoenig ЕШШЫЯдЪОЙ§УєдЬивьадBmemЖЈЮЛгк2аЭBmemШКЬхЃЌВЂаЏДјИпЧзКЭСІБэУцIg;ЭЌбљЃЌOta ЕШЯдЪОЙ§УєдЬивьадBЯИАћИпЖШЭЛБфВЂзЊТМFCERКЭжжЯЕIGHEЁЃ2 аЭ Bmem ЕФаЮГЩВЛвРРЕгкЩњЗЂжааФЃЌЕЋдкТбЧхЕААзЙ§УєаЁЪѓФЃаЭжаШЗЪЕашвЊ IL-4 ЁЃIL-4 ЪЧЙ§Уєжа 2 аЭУтвпЗДгІЕФЧ§ЖЏвђЫиЃЌПЩФмЪЧЬхФкНЯИпЕФ IL-4 ХЈЖШЃЈЮоТлЪЧдкжТУєЦкМфЛЙЪЧдкЙ§УєЪБЕФ 2 аЭУтвпЗДгІЃЉЖМЛсЧ§ЖЏ 2 аЭ Bmem ЕФВњЩњЁЃСэвЛжж 2 аЭ Bmem БъжОЮя FcІХRII/CD23 ЕФБэДягыбЛЗ IgE ХЈЖШГЪе§ЯрЙиЃЌвђЮЊ IgE НсКЯПЩЮШЖЈ FcІХRII ВЂдіМгБэУц CD23 ЗсЖШЁЃОЁЙмШчДЫЃЌCD23 ЫЦКѕЪЧвЛжжЮШЖЈЕФ 2 аЭ Bmem БъжОЮяЃЌвђЮЊ Koenig ЕШШЫБэУї CD23 2 аЭ Bmem ШКЬхдк IgE ШБЯнаЁЪѓКЭвАЩњаЭаЁЪѓжаЕФЪ§СПЯрЫЦЁЃзмжЎЃЌетаЉЪ§ОнБэУїЙ§УєдЬивьад 2 аЭ Bmem ЪЧдкжТУєЙ§ГЬжааЮГЩЕФЃЌВЂЧвПЩвдЭЈЙ§ЮШЖЈЕФБъжОЮяРДЖЈвхЁЃ+
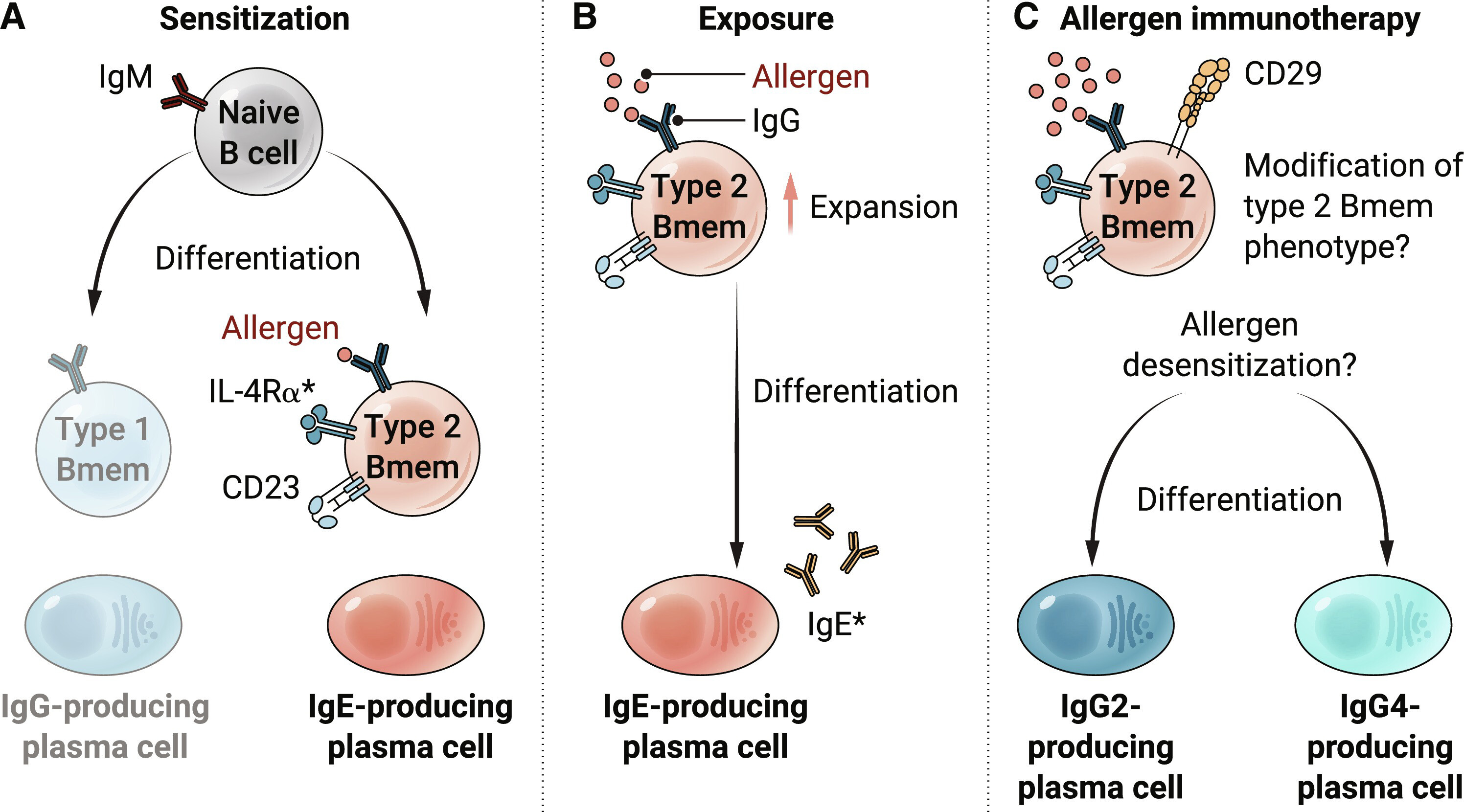
ЭМ 1.ВњЩњ IgE ЕФЯИАћЕФаЮГЩашвЊОпга 2 аЭБэаЭЕФЙ§УєдЬивьад BmemЁЃ
ЃЈAЃЉ Й§УєжТУєгыгзжЩ B ЯИАћЗжЛЏЮЊ 2 аЭ BmemЃЈзиЩЋЃЉЖјВЛЪЧ 1 аЭ BmemЃЈЛвЩЋЃЉвдМАВњЩњ IgE ЕФНЌЯИАћЕФДцдкгаЙиЁЃЃЈBЃЉ дкдйДЮБЉТЖЙ§УєдЪБЃЌ2 аЭ Bmem БЛМЄЛюЁЂРЉдіВЂзМБИЗжЛЏЮЊВњЩњ IgE ЕФНЌЯИАћЁЃаЧКХБэЪОПЩШмад IgE КЭ IL-4RІС ПЩЭЈЙ§ЕЅПЫТЁПЙЬхНјаажЮСЦадАаЯђжЮСЦЃЌвджЮСЦЙ§Ує ЁЃЃЈCЃЉ ЭЈЙ§ПижЦЩрЯТНгДЅЙ§УєдЃЈГЦЮЊ SLITЃЉНјааЙ§УєдУтвпСЦЗЈЃЌМйЩшЛсЕМжТ 2 аЭ Bmem ЕФРЉдіКЭЦфУтвпБэаЭЕФИФБфЁЃетаЉЯИАћЯждкФмЙЛЗжЛЏЮЊВњЩњ IgG2 КЭ IgG4 ЕФНЌЯИАћЃЌетгажњгкЙ§УєдЭбУєЁЃ
вђДЫЃЌЙ§УєжаЕФжТВЁад B ЯИАћШКЫЦКѕгаСНИівЊЧѓЃК2 аЭБэаЭКЭЖдЙ§УєдЬивьадЕФИпЧзКЭСІ B ЯИАћЪмЬхЁЃKoenigЕШШЫЕФЙлВьНсЙћжЇГжСЫетвЛЕуЁЃSARS-CoV-2 Ьивьад Bmem УЛга 2 аЭБэаЭ ЁЃДЫЭтЃЌ2 аЭ Bmem дкЗЧЙ§УєИіЬхжаЕФГіЯжЦЕТЪЕЭгкЙ§УєИіЬх ЁЃОЁЙмШчДЫЃЌШдШЛашвЊЛёЕУвђЙћЙиЯЕЕФе§ЪНжЄОнЃЌРэЯыЧщПіЯТЪЧЭЈЙ§дкСйДВЧАФЃаЭжаАаЯђКЭЯћКФ 2 аЭ BmemЃЌШЛКѓВтЪдгЩДЫВњЩњЕФЙ§УєЗДгІЁЃЛђепЃЌПЩвдЭЈЙ§ЦРЙРеыЖдЗЧжТУєШЫШКжаГЃМћЪГЮяЙ§УєдЃЈШчЛЈЩњЃЉЛђПеЦјЙ§УєдЃЈШчЮнГОђ§ЃЉЕФ Bmem БэаЭРДЛёЕУНјвЛВНЕФжЇГжжЄОнЁЃДЫЭтЃЌЙ§УєКЭЗЧЙ§УєИіЬхЕФ 2 аЭ Bmem ЬивьадМАЦфЖдФГаЉЙ§УєдЕФЧБдкПЫТЁРЉдіШдгаД§ВћУїЁЃ
ФПЧАЩаВЛЧхГўЪЧЪВУДЧ§ЖЏБэДя IgG ЕФ 2 аЭ Bmem ЗжЛЏЮЊВњЩњ IgE ЕФНЌЯИАћЁЃKoenig ЕШШЫЯдЪОЙ§УєдУтвпжЮСЦ 1 ИідТКѓЕФ IgE зЊТМБОдкжЮСЦПЊЪМЧАЖЈЮЛгк 2 аЭ Bmem ДиЁЃIL-13 гЩ T ТЫХнИЈжњЯИАћВњЩњЃЌЧ§ЖЏ B ЯИАћЕФЬхЯИАћГЌЭЛБфКЭИпЧзКЭСІЙ§УєдЬивьад IgE ЕФВњЩњЁЃЪТЪЕЩЯЃЌIL4R КЭ IL13RA1ЃЈЙВЭЌБрТы IL-13 ЪмЬхЃЉЕФБэДядк 2 аЭ Bmem жаЩЯЕїЃЌБэУїетаЉЯИАћПЩНгЪм IL-13ЁЃШЛЖјЃЌСїЪНЯИАћЪѕЮоЗЈШЗШЯБэУцБъжОЮяБэДяЃЌетгыЮвУЧжЎЧАЖдКкТѓВнЛЈЗлЙ§УєИіЬхЙ§УєдЬивьад Bmem ЕФЙлВьРрЫЦЁЃвђДЫЃЌашвЊНјвЛВНЕФбаОПРДНвЪОЧ§ЖЏ 2 аЭ Bmem ЗжЛЏЮЊВњЩњЙ§УєдЬивьад IgE ЕФНЌЯИАћЕФвђЫиЁЃ
ФПЧАЛЙВЛЧхГў 2 аЭ Bmem ЪЧЗёЗДгГСЫ TH2 ИіОпгаЖРЬизЊТМвђзгЕФЯИАћЃЌОіЖЈСЫЫќУЧЕФБэаЭКЭЙІФмЃЌЛђепетаЉЯИАћЪЧЗёЪЧОпгаПЩЫмадБэаЭКЭЙІФмЕФМЋЛЏ BmemЃЌРрЫЦгкЕїНкад B ЯИАћЁЃ 2 аЭ Bmem жаЕФЬиЖЈзЊТМвђзгЃЌБэУї Bmem МЋЛЏПЩФмдкЙІФмЩЯгы T ЯИАћВЛЭЌЁЃБэДя IL-10 ЕФЕїНкад B ЯИАћвВЪЧШчДЫЃЌетаЉЯИАћдкЙІФмЩЯЪЧУїШЗЕФЃЌВЛЙЙГЩЕЅИібЧШКЛђЕЅЖРЕФЦзЯЕЁЃНјвЛВНЕїВщЙ§УєКЭЗЧЙ§УєИіЬхжа 2 аЭ Bmem ЕФзнЯђзЊТМзщПЩвдИќЧхГўЕиСЫНтЦфЖЈвхКЭПЩЫмадЁЃ
Koenig ЕШШЫЭЈЙ§ЗжЮіНгЪмЙ§УєдУтвпжЮСЦЕФЛМепбљБОЃЌМьВщЙ§УєдБЉТЖЖд 2 аЭ Bmem ЕФгАЯьЁЃЙ§УєдУтвпСЦЗЈЪЧЮЈвЛПЩгУЕФЙ§УєМВВЁИФЩЦСЦЗЈЁЃЭЈЙ§ГжајЖЈЦкЩрЯТЛђЦЄЯТзЂЩфЙ§УєдЃЌ2 аЭУтвпСІЛсЗЂЩњБфЛЏЃЌДгЖјЪЙЛМепЖдЙ§УєдЭбУєЁЃЫфШЛСЦаЇПЩвдДяЕН80%ЃЌЕЋетЪЧвЛИіЪ§дТжСЪ§ФъЕФГЄЦкЙ§ГЬЁЃKoenig ЕШШЫЯдЪОЃЌшыЪїЛЈЗлSLIT1ИідТКѓЃЌЙ§УєдЬивьад2аЭBmemЦЕТЪдіМгЁЃживЊЕФЪЧЃЌ1ИідТШдДІгкжЮСЦЕФдчЦкЃЌЭЈГЃдкЭбУєПЊЪМжЎЧАЁЃвђДЫЃЌSLIT Жд 2 аЭ Bmem Ъ§КЭУтвпБэаЭЕФгАЯьЩаВЛЧхГўЁЃЮвУЧжЎЧАБэУїЃЌЖдКкТѓВнЛЈЗлЙ§УєЕФИіЬхНјааЮЊЦк 4 ИідТЕФ SLIT СЦГЬПЩМѕЧсЙ§УєжЂзДВЂИФБф Bmem ЖдКкТѓВнЛЈЗлЕФжївЊЙ§Уєд Lol p 1 ЬивьадЕФБэаЭЁЃЭЈЙ§ЕЅЯИАћ RNA ВтађЃЌдк SLIT 4 ИідТКѓРЉдіСЫ 2 Иі Lol p 1 Ьивьад Bmem ДиЃЌЦфжавЛИіДиЕФЬиеїЪЧ IGHEЁЂFCER2ЁЂIL4R КЭ IL13RA1 ЩЯЕїЃЌБэУїЙ§УєдЬивьад 2 аЭ Bmem РЉдіЁЃетОЭЬсГіСЫвЛИіЮЪЬтЃЌМД 2 аЭ Bmem ЪЧжТВЁЕФЛЙЪЧБЃЛЄадЕФЁЃживЊЕФЪЧЃЌSLIT КѓЕФ 2 аЭ Bmem ДиЛЙЯдЪО IGHG4 КЭ ITGB1 ЩЯЕїЃЌКѓепБрТы CD29ЃЈећКЯЫи ІТ1ЃЉВЂИКЕїПиБэУц Ig аХКХзЊЕМЁЃашвЊНјаазнЯђбаОПвдШЗЖЈИУбЧШКЕФБэаЭБфЛЏЪЧЗёгажњгкФЭЪмадЛђМВВЁЁЃ
СэвЛИіЧАОАЪЧгУЩњЮяжЦМСАаЯђ 2 аЭ BmemЁЃКЯКѕТпМЕФКђбЁвЉЮяЪЧЖШЦеРћгШЕЅПЙЃЌетЪЧвЛжж IL-4RІС зшЖЯПЙЬхЃЌОпгаЫЋжизїгУЃЌвђЮЊЫќПЩзшжЙ IL-4 КЭ IL-13 гыЦфЪмЬхНсКЯЁЃЕкЖўжжКђбЁвЉЮяЪЧАТТэжщЕЅПЙЃЌетЪЧвЛжжПЙIgEПЙЬхЃЌПЩгыIgEЕФFcЦЌЖЮНсКЯЃЌДгЖјзшжЙIgEгыЗЪДѓЯИАћКЭЪШМюадСЃЯИАћЩЯЕФFcІХRIНсКЯЁЃМИЯюбаОПвбОШЗЖЈЃЌдкНгЪметСНжжПЙЬхжЮСЦЕФИіЬхжаЃЌЙ§УєжЂзДКЭЙ§УєадЯјДЗЂзїгаЫљМѕЩйЃЌЕЋЕНФПЧАЮЊжЙЃЌИпАКЕФГЩБОМѕЛКСЫЙуЗКЩуШЁЕФЫйЖШЁЃСэвЛжжжЮСЦЭООЖЪЧНЋЙ§УєдУтвпСЦЗЈгыАаЯђIL-4RЛђIgEЕФЩњЮяжЦМСЯрНсКЯЃЌетПЩвдНјвЛВНМѕШѕ2аЭBmemЃЌВЂЭЈЙ§ИФБфЛђЯћГ§жТВЁадIgEЕФРДдДРДЬсЙЉГЄЦквцДІЁЃ
змжЎЃЌOta ЕШШЫКЭ Koenig ЕШШЫСНепЖМШЗЖЈСЫдкЙ§УєадМВВЁЛМепжаРЉеЙЕФ 2 аЭ Bmem бЧШКЁЃОЁЙмИУбЧШКБэДяБэУцФЄ IgG1ЃЌЕЋЫќвВБэДя FcІХRIIЁЂIL-4RІС КЭжжЯЕ IGHE зЊТМБОЃЌЪЙЦфдк 2 аЭЛЗОГжаМЄЛюКѓгаЭћЗжЛЏЮЊВњЩњ IgE ЕФНЌЯИАћЁЃвђДЫЃЌЙ§УєИіЬхжажТВЁад IgE ЕФРДдДЫЦКѕЪЧ B ЯИАћЃЌИУ B ЯИАћОпга ЃЈiЃЉ ЖдЯрЙиЙ§УєдГЩЗжЕФИпЧзКЭСІКЭ ЃЈiiЃЉ ОпгаЧаЛЛЕН IgE ЧуЯђЕФ 2 аЭБэаЭЁЃЛЛОфЛАЫЕЃЌЫќашвЊСНИіЁЃ
https://blog.sciencenet.cn/blog-41174-1421434.html
ЩЯвЛЦЊЃКбЊЙмжмЮЇГЩЯЫЮЌЯИАћПЩФмЪЧEDЕФЛљДЁ
ЯТвЛЦЊЃКЁАБШЖћЁЄИЧДФЮЪЬтЁБЃКвкЭђИЛЮЬДШЩЦМвЪЧЗёХЄЧњСЫШЋЧђНЁПЕбаОПЃП
