ВЉЮФ
БэЙлвХДЋаоЪЮЃКЯывЊИФБфЛљвђБэДяЃПВЛжЛга DNAЃЌШОЩЋЬхвВПЩвдЃЁ_ MCE(MedChemExpress)
|
Section.01
БэЙлвХДЋбЇ
БэЙлвХДЋбЇ (Epigenetics) ЪЧжИбаОПЭЈЙ§ИФБфШОЩЋЬхЖјЗЧ DNA ађСаЖјВњЩњЕФЁЂПЩвХДЋЧвЮШЖЈЕФЛљвђБэДяБфЛЏЕФбЇПЦЁЃ
БэЙлвХДЋЕФЛњжЦЪЧЭЈЙ§ DNA МюЛљЕФЛЏбЇаоЪЮКЭШОЩЋЬхГЌНсЙЙЕФИФБфРДЕїНкЛљвђБэДяЁЃБэЙлвХДЋбЇЕФЭМЦзШевцИДдгЃЌЫќВЛНіАќКЌ DNA МзЛљЛЏЁЂзщЕААзУмТыЁЂЗЧБрТыRNA (ncRNA) КЭКЫаЁЬхЖЈЮЛЃЌЛЙгы DNA ађСаЙВЭЌЙЙГЩвЛИіЭъећЕФЕїПиЬхЯЕЁЃБэЙлвХДЋЭМЦзЕФЮЩТвгыЖржжШЫРрМВВЁЕФЗЂВЁЛњжЦНєУмЯрЙи[1]ЁЃ
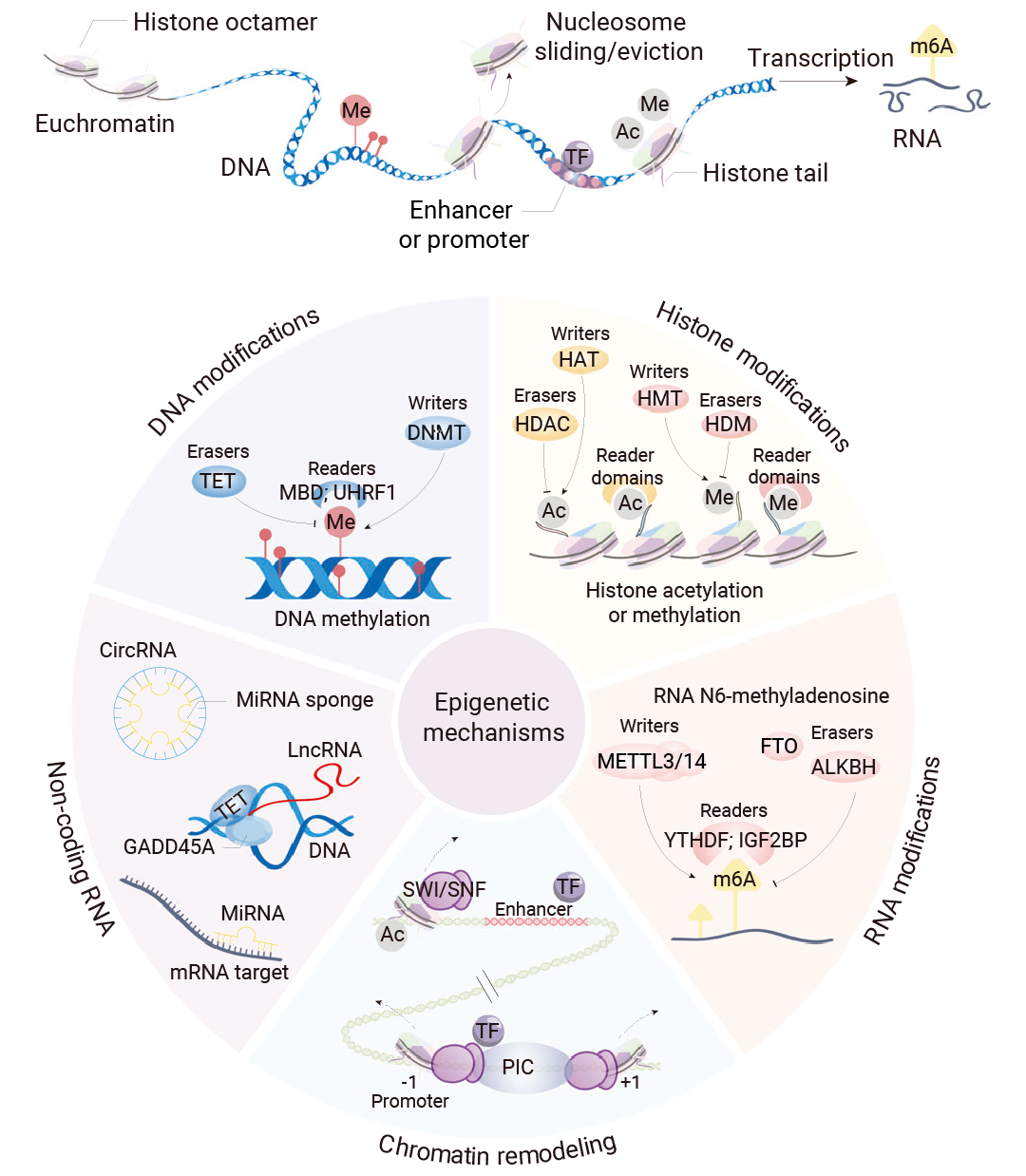
ЭМ 1. БэЙлвХДЋЛњжЦКЭЙуЗКбаОПЕФаоЪЮМАЦфаоЪЮУИ[2]ЁЃ
БэЙлвХДЋЕФЕїПи
БэЙлвХДЋаоЪЮЭЈЙ§Ш§РрЕїПивђзгЪЕЯжЃК (1) в§ШыИїжжЛЏбЇЛљЭХЕФЁАаДШыЦїЁБ (Writers) ЃЛ (2) ЬивьадЪЖБ№КЭНтЖС DNA ЛђепзщЕААзЩЯаоЪЮЕФЕААзжЪНсЙЙгђЃЌМДЁАЖСШЁЦїЁБ (Readers) ЃЛ (3) вдМАЧхГ§аоЪЮЕФЁАВСГ§ЦїЁБ (Erasers) ЁЃ
ОйИіР§згЃЌБэЙлЪЖБ№ЕААзНсЙЙгђ (Epigenetic Reader Domains) ЪЧЕААзжЪЩЯФмЙЛЬивьадЪЖБ№ВЂНсКЯЬиЖЈЕФБэЙлвХДЋаоЪЮБъМЧЕФЕААзжЪНсЙЙгђЃЌЪЧБэЙлвХДЋжаЕФЁАЖСШЁЦїЁБЃЌНЋБэЙлвХДЋБъМЧЕФЛЏбЇаХЯЂЁАЗвыЁБГЩЬиЖЈЕФЩњЮябЇЪфГіЃЌР§ШчЛљвђБэДяЕФМЄЛюЁЂвжжЦЁЂDNA Ы№ЩЫаоИДЛђШОЩЋжЪжиЫмЃЌНјЖјеаФМЦфЫћИДКЯЮяРДгАЯьШОЩЋжЪНсЙЙКЭЛљвђзЊТМ[2]ЁЃ
ЭМ 2. зщЕАЗвыКѓаоЪЮЕФЁАЖСШЁЦїЁБ[3]ЁЃ
змжЎЃЌЁАЖСШЁЦїЁБЁЂЁАаДШыЦїЁБКЭЁАВСГ§ЦїЁБаЭЌзїгУЃЌаЕїЯИАћБэЙлвХДЋЛњжЦКЭЛљвђБэДяЃЌВЂЧветаЉБэЙлвХДЋаоЪЮвђзггыЖржжМВВЁЃЌгШЦфЪЧАЉжЂУмЧаЯрЙи[2]ЁЃ
Section.02
DNA МзЛљЛЏЃК
ЛљвђЕФ"ГСФЌБъЧЉ"
DNA МзЛљЛЏ (DNA methylation) ЪЧжИдк DNA МзЛљзЊвЦУИ (DNMTs) ЕФДпЛЏЯТЃЌНЋМзЛљЬэМгЕН DNA ЩЯЕФЙ§ГЬЁЃDNA МзЛљЛЏЗЂЩњКѓЃЌВЮгыЛљвђвжжЦЕФЕААзЛђвжжЦзЊТМвђзгБЛФММЏгы DNA НсКЯРДвжжЦЛљвђБэДяЁЃ

DNA МзЛљЛЏжївЊЗЂЩњдкАћрзрЄ-ФёрбпЪ (CpG) ЖўКЫмеЫсЮЛЕуЃЌгШЦфЪЧЦєЖЏзгЧјгђФкЕФ CpG ЕКЁЃCpG ЕК (CpG Island) ЪЧЛљвђзщ DNA жаИЛКЌ CpG ЕФЧјгђЃЌМзЛљЛЏГЬЖШЕЭЃЌдкДѓЖрЪ§ВИШщЖЏЮяЛљвђзщжаЃЌCpG ЕКЮЛгкЛљвђЦєЖЏзгЩЯЃЌЪЙЛљвђОЭДІгкЁАД§УќЁБзДЬЌЁЃCpG ЕКЪЧЕїПиЛљвђБэДяЕФЁАПЊЙиЁБ[1][7]ЁЃ
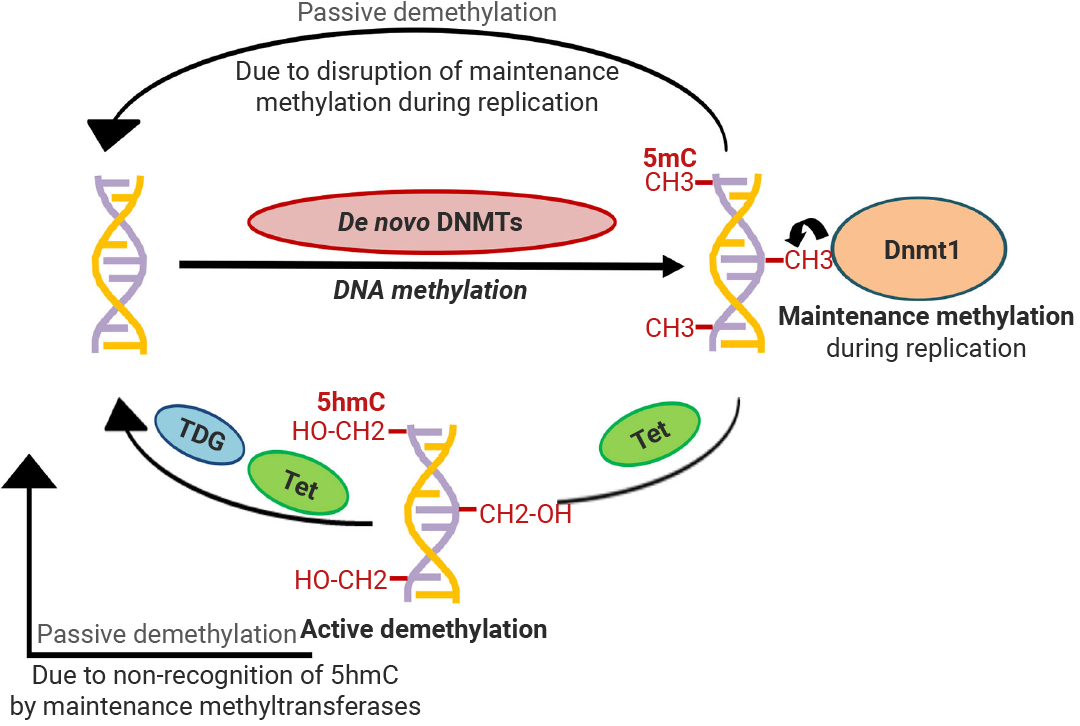
ЭМ 3. DNA МзЛљЛЏФЃЪН[8]ЁЃ
DNA ШЅМзЛљЛЏ (DNA demethylation) ПЩЗжЮЊБЛЖЏШЅМзЛљЛЏгыжїЖЏШЅМзЛљЛЏЁЃ
? БЛЖЏШЅМзЛљЛЏЪЧгЩгк DNMT1 ЙІФмБЛвжжЦЖјв§Ц№ЕФЃЌЗЂЩњдк DNA ИДжЦЙ§ГЬЁЃ
? жїЖЏШЅМзЛљЛЏЃКЭЈЙ§ОЕфЭЈТЗ TET-TDG-BER НЋ 5mC бѕЛЏВЂвЦГ§ПЩЪЕЯжжїЖЏШЅМзЛљЛЏЃЌВЛвРРЕгк DNA ИДжЦЁЃ
Section.03
зщЕААзаоЪЮЃК
ШОЩЋжЪЕФ"ЕїЙтПЊЙи"
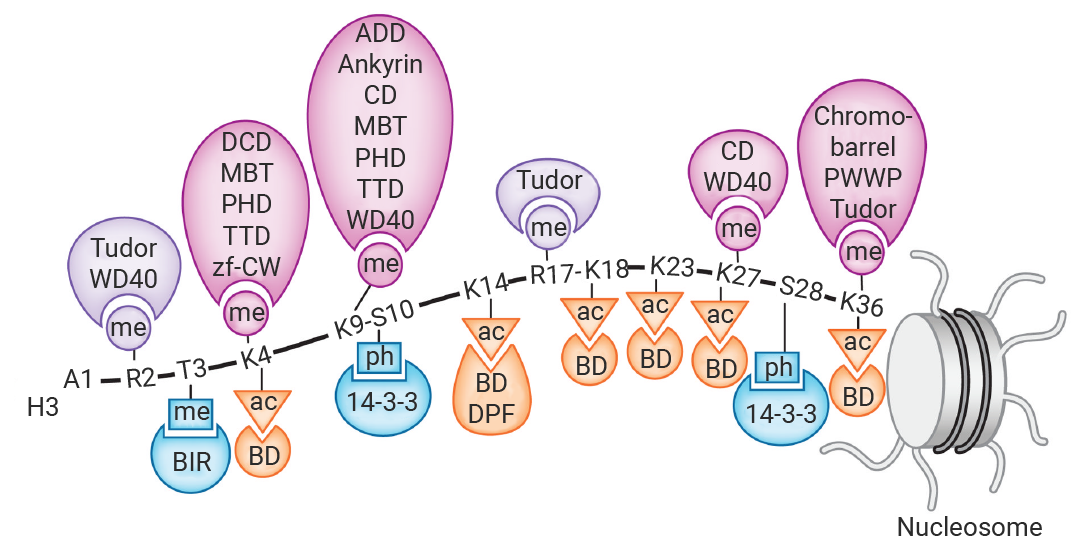
зщЕААз H2AЁЂH2BЁЂH3 КЭ H4 ИїСНИіЗжзгзщГЩАЫОлЬхЃЌDNA ВјШЦЦфЩЯаЮГЩКЫаЁЬхЃЌВЂгЩ H1 зщЕААзСЌНгДЎСЊЁЃИїКЫаФзщЕААзОљгавЛЬѕЗЧНсЙЙЛЏЕФАБЛљФЉЖЫЁАЮВАЭЁБбгЩьжСКЫаЁЬхЭтВПЃЌЮЊввѕЃЛЏЁЂСзЫсЛЏКЭМзЛљЛЏЕШЗвыКѓаоЪЮЬсЙЉЮЛЕуЁЃ
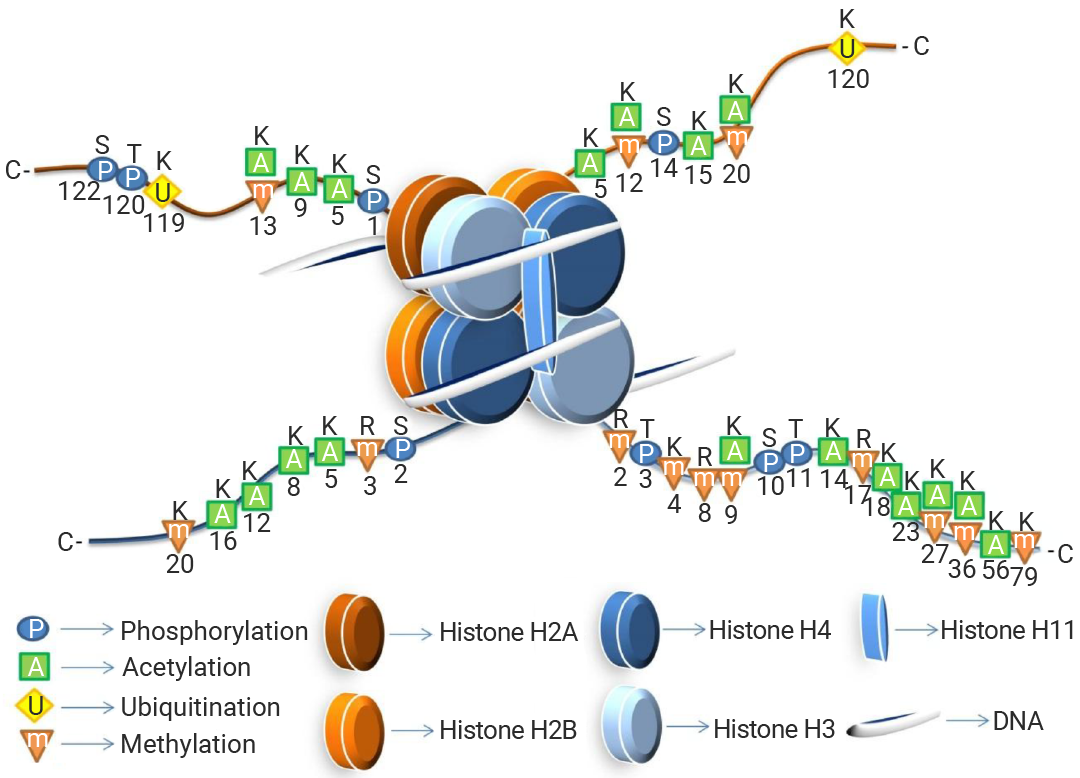
ЭМ 4. зщЕААзЕФНсЙЙЪОвтЭМ[9]ЁЃ
зщЕААзаоЪЮЕФжївЊЗНЪНга 2 жжЃКМзЛљЛЏКЭввѕЃЛЏЁЃ
зщЕААзМзЛљЛЏ
зщЕААзМзЛљЛЏ (Histone Methyltransferase) гЩзщЕААзМзЛљзЊвЦУИ (HMTs) ДпЛЏЃЌжИдкзщЕААз (ЬиБ№ЪЧ H3 КЭ H4) ЕФРЕАБЫсЛђОЋАБЫсВаЛљЩЯЬэМгвЛжСШ§ИіМзЛљЛљЭХЕФЙ§ГЬЁЃ
ИУЙ§ГЬДІгкЖЏЬЌЦНКтЃЌгЩМзЛљзЊвЦУИ (ЁАаДШыЁБ) КЭШЅМзЛљЛЏУИ (ЁАВСГ§ЁБ) аЭЌЕїПиЁЃзщЕААзМзЛљЛЏЕФЙІФмИДдгЃЌЦфЖдЛљвђБэДяЕФЕїПиШЁОігкОпЬхЮЛЕуЃКР§ШчЃЌH3K4ЁЂH3K36ЁЂH3K79 ЕФМзЛљЛЏМЄЛюЛљвђзЊТМЃЌЖј H3K9ЁЂH3K27 ЕФМзЛљЛЏЦ№вжжЦзїгУ[10]ЁЃ
ИљОн HMTs ДпЛЏЕФАБЛљЫсПЩЗжЮЊСНДѓМвзхЃКзщЕААзРЕАБЫсМзЛљзЊвЦУИ (KMTs) КЭЕААзжЪОЋАБЫсМзЛљзЊвЦУИ (PRMTs) ЁЃKMTs МвзхАќРЈ EZH2ЁЂG9aЁЂDOT1L КЭ SETD2ЃЛPRMTs МвзхдкВИШщЖЏЮяжаЙВга 9 ИіГЩдБ (PRMT1-9)ЁЃИљОнЬэМгЕФМзЛљЪ§СПЃЌРЕАБЫсВаЛљЩЯЛсаЮГЩЕЅМзЛљЁЂЖўМзЛљКЭШ§МзЛљЛЏЛљЭХ (Kme1ЁЂKme2ЁЂKme3) ЃЌЖјОЋАБЫсВаЛљЩЯЛсаЮГЩЕЅМзЛљКЭЖўМзЛљ (Rme1/MMAЁЂRme2) [10][11]ЁЃ
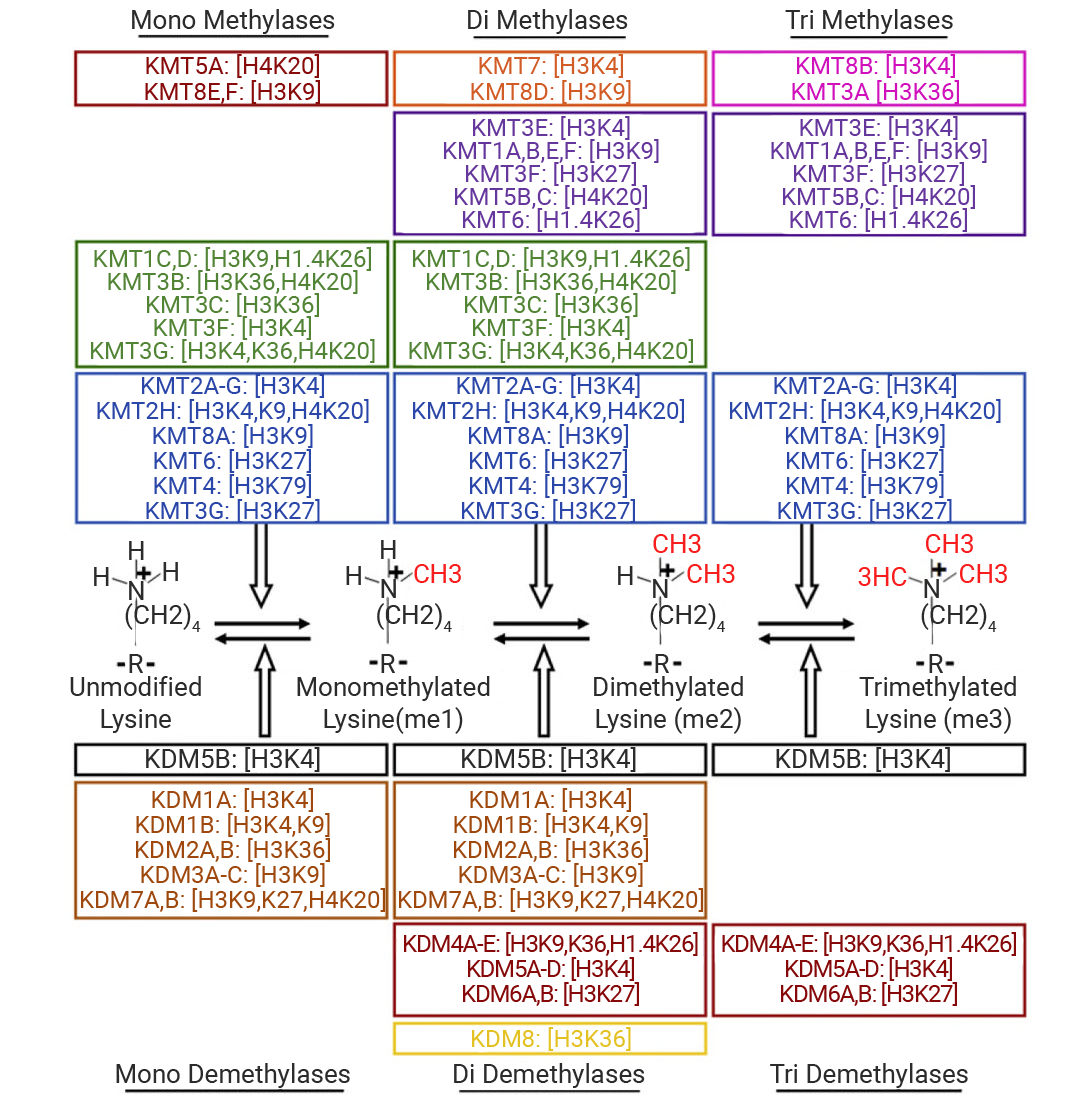
ЭМ 5.РЕАБЫсМзЛљзЊвЦУИ (KMTs) КЭРЕАБЫсШЅМзЛљЛЏУИ (KDM) МАЦфЖдгІЕзЮя[11].
МИКѕЫљгажївЊзщЕААзРЕАБЫсМзЛљЛЏЮЛЕуЕФМзЛљЛЏзДЬЌЖМЪЧПЩФцЕФЁЃзщЕААзШЅМзЛљЛЏгЩзщЕААзШЅМзЛљЛЏУИ (HDMs) ДпЛЏЁЃHDMs жївЊга LSD Мвзх (РЕАБЫсЬивьадШЅМзЛљЛЏУИ) ЃЌШч LSD1 (KDM1A)ЁЂLSD2 (KDM1B) КЭ JmjC Мвзх (Jumonji C НсЙЙгђЕААз) ЃЌШч KDM2ЁЂKDM3ЁЂKDM4 [12] [13] [14] ЁЃ
зщЕААзввѕЃЛЏ
зщЕААзввѕЃЛЏ (Histone Acetylation) ЪЧжИдкзщЕААз N ЖЫЮВВПРЕАБЫсВаЛљЩЯЬэМгввѕЃЛљЕФаоЪЮЃЌМѕШѕ DNA гызщЕААзжЎМфЕФЯрЛЅзїгУЃЌЕМжТШОЩЋжЪНсЙЙДгжТУмБфЕУЫЩЩЂЃЌDNA ИќШнвзБЛзЊТМвђзгКЭ RNA ОлКЯУИЕШзЊТМзщЗжНгДЅЕНЃЌДгЖјМЄЛюЛљвђзЊТМЁЃ
ИУЙ§ГЬгЩзщЕААзввѕЃзЊвЦУИ (HATs) ЁАаДШыЁБЃЌзщЕААзШЅввѕЃЛЏУИ (HDACs) ЁАВСГ§ЁБЁЃввѕЃЛЏ/ШЅввѕЃЛЏЪЧвЛИіЯрЖдПьЫйЕФЙ§ГЬЃЌдЪаэЯИАћбИЫйЯьгІЛЗОГаХКХ (ШчМЄЫиЁЂгІМЄЁЂгЊбјБфЛЏ) РДЕїећЛљвђБэДяЁЃзщЕААзввѕЃЛЏЫЎЦНЕФвьГЃЕїПигыЖржжМВВЁЯрЙиЃЌАќРЈбзжЂадМВВЁЁЂаФбЊЙмМВВЁЁЂЩёОЯЕЭГМВВЁКЭЖржжАЉжЂ[14]ЁЃ
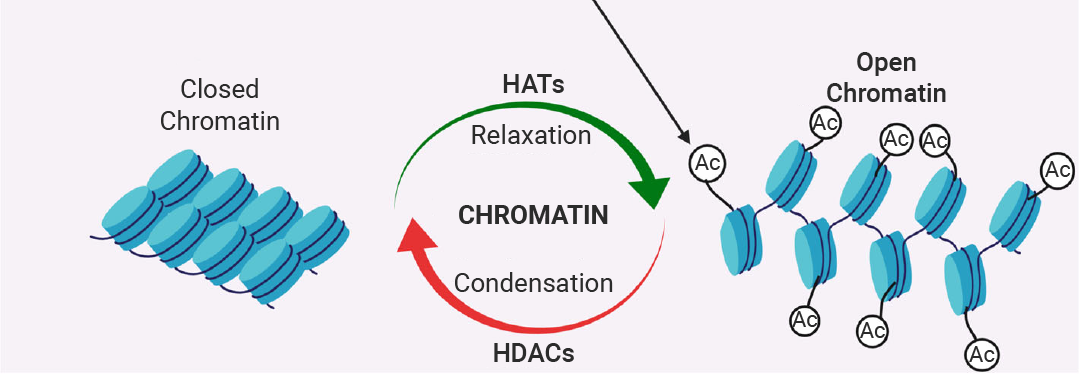
ЭМ 6. ШОЩЋжЪЪмСНзщУИ HATs КЭ HDACs ЕФЕїПи[14]ЁЃ
зщЕААзШЅввѕЃЛЏУИЭЈЙ§вЦГ§зщЕААзЩЯЕФввѕЃЛљЭХЃЌвжжЦЛљвђБэДяВЂЕїПиШОЩЋжЪНсЙЙЁЃдкШЫРржаЙВга 18 жж HDAC УИЃЌИљОнЦфгыНЭФИЭЌдДЮяЕФЯрЫЦадКЭЙІФмВювьЃЌЗжЮЊЫФИіжївЊРрБ№ЃК
I Рр (Class I)ЃКАќРЈ HDAC1ЁЂHDAC2ЁЂHDAC3 КЭ HDAC8ЃЛ
II Рр (Class II)ЃКЗжЮЊ IIa (HDAC4ЁЂHDAC5ЁЂHDAC7ЁЂHDAC9) КЭ IIb (HDAC6ЁЂHDAC10) ЃЛ
III Рр (Class III)ЃКвВГЦЮЊ Sirtuins (SIRT1-SIRT7)ЃЛ
IV Рр (Class IV)ЃКФПЧАНіАќКЌ HDAC11ЁЃ
HDACs ЕФЙІФмвьГЃгыЖржжМВВЁЯрЙиЃЌЬиБ№ЪЧАЉжЂЁЃдкаэЖржзСіжаЃЌHDAC ЛюадвьГЃЩ§ИпЃЌЕМжТвжАЉЛљвђБЛГСФЌ[14]ЁЃ
ВњЦЗЭЦМі
жизщЕФ DNMT1 ЕААз P26358 (P351-A1600) ЦЌЖЮЃЌгЩ E. coli БэДяЃЌДјга tag Free БъЧЉЁЃ
DNMT2ЕААз, Human (sf9, GST) (HY-P75711)
жизщЕФ DNMT2 ЕААзЃЌгЩ Sf9 insect cells БэДяЃЌДјга N-GST БъЧЉЁЃ
Histone H1ЕААз, Human (His) (HY-P74887)
жизщЕФзщЕААз H1 ЕААзЃЌгЩ E. coli БэДяЃЌДјга N-His БъЧЉЁЃ
Histone deacetylase 1/HDAC1ЕААз, Human (His-SUMO) (HY-P72262)
жизщЕФзщЕААзЭбввѕЃУИ HDAC1 ЕААзЃЌгЩ E. coli БэДяЃЌДјга N-SUMO, N-6*His БъЧЉЁЃ
HDAC6ЕААз, Human (His) (HY-P72224)
жизщЕФ HDAC6 ЕААзЃЌгЩ E. coli БэДяЃЌДјга N-6*His БъЧЉЁЃ
жизщЕФ Menin ЕААзЃЌгЩ E. coli БэДяЃЌВЛДјБъЧЉЁЃMenin ЕААзЪЧвЛжжФкЗжУкжзСівжжЦвђзгКЭзЊТМЕїНквђзгЃЌвВЪЧ KMT2A ЕААзЗЂЛгЙІФмЫљБиашЕФЁАУЊЕуЁБЃЌвдМЄЛю HOXA КЭ MEIS1 ЕШЛљвђЁЃ
ВЮПМЮФЯз
[1] Jin B, et al. DNA methylation: superior or subordinate in the epigenetic hierarchy? Genes Cancer. 2011 Jun;2(6):607-17.
[2] Biswas S,et al. Epigenetic tools (The Writers, The Readers and The Erasers) and their implications in cancer therapy. Eur J Pharmacol. 2018 Oct 15;837:8-24.
[3] Musselman, C., et al. Perceiving the epigenetic landscape through histone readers. Nat Struct Mol Biol 19, 1218ЈC1227 (2012).
[4] Kikuchi A, et al. Structural basis for activation of DNMT1. Nat Commun. 2022 Nov 21;13(1):7130.
[5] Okano M, et al. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell. 1999 Oct 29;99(3):247-57.
[6] Veland N, et al. DNMT3L facilitates DNA methylation partly by maintaining DNMT3A stability in mouse embryonic stem cells. Nucleic Acids Res. 2019 Jan 10;47(1):152-167.
[7] Moore LD, et al. DNA methylation and its basic function. Neuropsychopharmacology. 2013 Jan;38(1):23-38.
[8] Parveen N, et al. DNA Methylation Patterning and the Regulation of Beta Cell Homeostasis. Front Endocrinol (Lausanne). 2021 May 7;12:651258.
[9] Biswas S, Rao CM. Epigenetics in cancer: Fundamentals and Beyond. Pharmacol Ther. 2017;173:118-134.
[10] Yang C, et al. Histone methyltransferase and drug resistance in cancers. J Exp Clin Cancer Res. 2020 Aug 28;39(1):173.
[11] Black JC, et al. Histone lysine methylation dynamics: establishment, regulation, and biological impact. Mol Cell. 2012 Nov 30;48(4):491-507.
[12] Shi YG, et al. The discovery of histone demethylases. Cold Spring Harb Perspect Biol. 2013 Sep 1;5(9):a017947.
[13] Hyun K, et al. Writing, erasing and reading histone lysine methylations. Exp Mol Med. 2017 Apr 28;49(4):e324.
[14] Gujral P, et al. Histone acetylation and the role of histone deacetylases in normal cyclic endometrium. Reprod Biol Endocrinol. 2020 Aug 13;18(1):84.
[15] Mabe NW, et al. Pharmacological targeting of the cancer epigenome. Nat Cancer. 2024 Jun;5(6):844-865.
[16] Thakur RK, et al. The promise of menin inhibitors: from approval to triplet regimens. Hematology Am Soc Hematol Educ Program. 2025 Dec 5;2025(1):599-606.
[17] Liang J, et al. Ailanthone targets the KMT2A-MEN1 complex to suppress lung metastasis of osteosarcoma. Phytomedicine. 2025 Jan;136:156258.
https://blog.sciencenet.cn/blog-3536222-1536588.html
ЩЯвЛЦЊЃКFITC-Labeled CD70 ЕААзЪЧЛюЛЏ T КЭ B СмАЭЯИАћЩЯЕФБэУцПЙд_MCE(MedChemExpress)
ЯТвЛЦЊЃКСДУЙЧзКЭЫиДХжщЃККЫЫсгыЕААзбаОПЕФИпаЇЙЄОп _ MCE(MedChemExpress)

ШЋВПзїепЕФЦфЫћзюаТВЉЮФ
- • вЉЮябаЗЂЃКПЙОњвЉЮявЉаЇЃЈPDЃЉвЉДњЃЈPKЃЉЕФИјвЉТпМ_MCE(MedChemExpress)
- • AІТЕААзгыАЂЖћДФКЃФЌжЂ_MCE(MedChemExpress)
- • ЯИАћХрбјЃКРДдДЁЂРраЭЁЂХрбјЬѕМўЁЂГЃМћЮлШО_MCE(MedChemExpress)
- • вЉЮяЕнЫЭЃКПЙЬхХМСЊжЌжЪФЩУзПХСЃ (Ab-LNP) ЁЊ ЬхФк CAR-T ЕФЧАбиВпТд_MCE/MedChemExpress
- • РрЦїЙйШОЩЋЃКШчКЮбЁдёгЋЙтШОСЯ_MCE(MedChemExpress)
- • ПЦбажњЙЅ | жЪЦзМЖЕААзУИЃЌЕНЕзгаЖрЁАЖЅЁБЃП_ MCE(MedChemExpress)