博文
我们不知道答案的125个科学问题(97)生命之树  精选
精选
||
97.会不会存在一个生物分类学家都认同的生命之树?
Will there ever be a tree of life that systematists can agree on?
题记:尽管科学家发展出更好的形态学、分子学和统计学的研究方法,但生命科学家们所得到的生物进化树并不统一。科学研究正在期待更多(但并非完全一致)的生命之树的共识。
1. 生命进化之树
达尔文的进化论发现了多样性生物之间存在某种关系,如果将这种形态学上的关系用图形结构表达出来的话,就能得到一棵生命之树(tree of life)。达尔文《物种起源》中发现所有已知的生物都可以通过共同的祖先联系在一起,就像一棵向外不断生长的树,叶子代表今天存在的各种生命有机体,树的树枝代表了这些有机体之间的进化关系,我们称之为生命之树。自达尔文时代以来,科学家们一直致力于在这棵树上绘制所有生命体的地图,以展示地球上所有形式的生命体是如何进化和相互联系的。1870年,德国科学家恩斯特·海克尔(Ernst Haeckel)第一个用“生命之树”描绘了植物和动物的进化关系,从那时起,科学家们就不断通过现代生物的DNA分子等数据不断添加各种物种、微生物等去更新、扩展和完善这棵生命之树。随着这棵生命树的成长,人类对地球上生命的理解也在不断增长(Nature Microbiology 9: 1906–1908, 2024)。然而,如果生命之树真的存在,那这棵隐藏在生物大数据背后高大而壮丽的树到底是什么样子?然而因为这棵树的许多部分和枝干到现在还不清楚并存在巨大争议,生命之树的形状到现在依然还没有一个统一的认识。
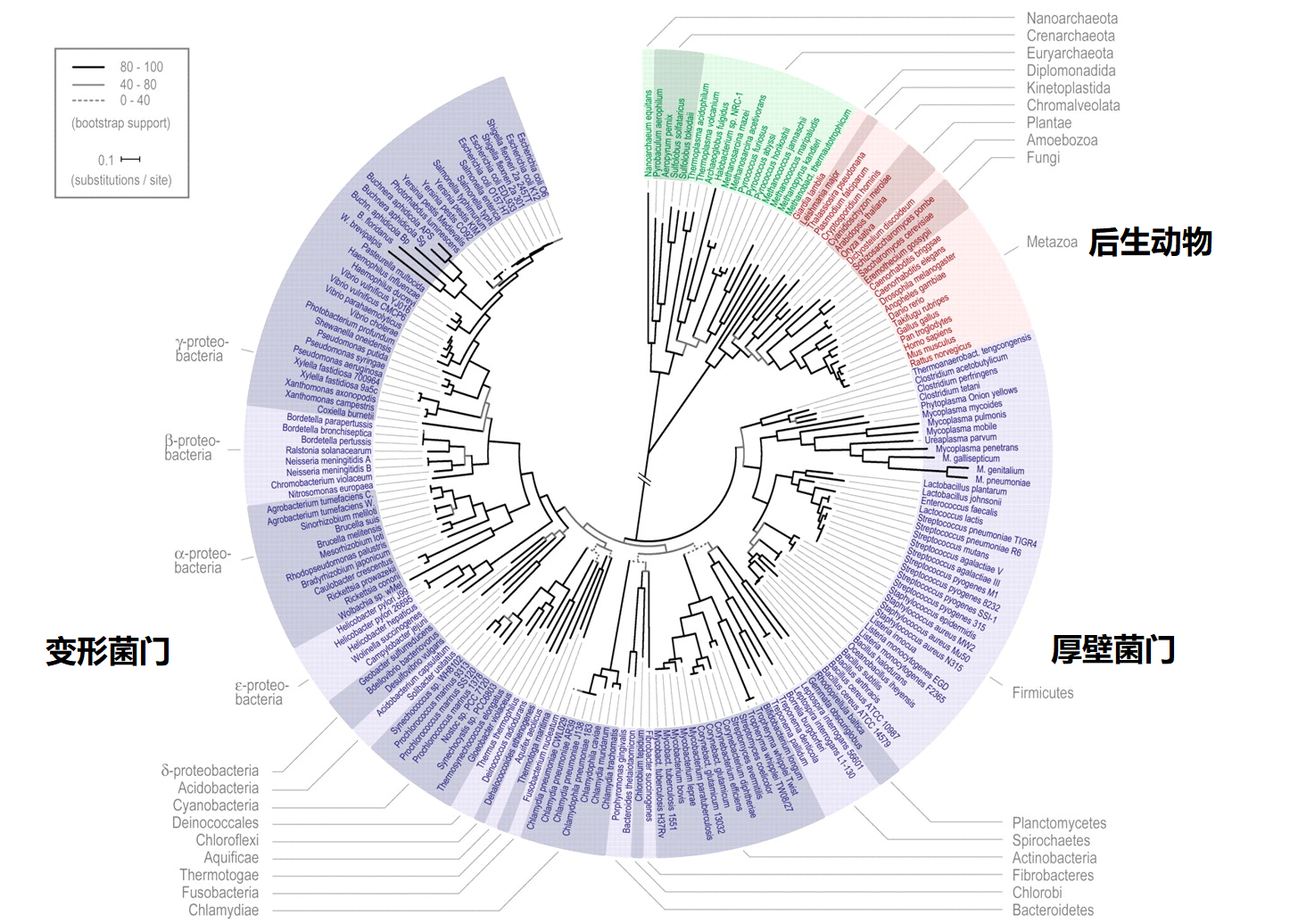
图1 2006年生物界的系统发育树
2006年欧洲海德堡分子生物学实验室(EMBL)利用新的自动算法生成了当时最为准确的生命之树(Science 311:1283-1287, 2006),如图1所示。图中通过对191个基因组已经完全测序的物种的31个通用蛋白家族进行了数据分析,明显显示了生命树由三个树干组成。之后,生物学家利用不同的方法对生命树进行了各个方面的完善和展示,2016年,给出了新的生命树,如图2所示(Nat. Microbiol 1, 16048, 2016)。图2是由生物分子信息构建的三域树之一,也是自基因组解析学发展以来首次发表的综合树。该生命树是通过核糖体RNA基因序列(生物的分子指纹)补充了原来生命树所忽略的树干:微生物部分(发现细菌树干应该分为两个子部分:细菌和候选门辐射类群),从而重塑了生命之树的整体结构,刷新了生命之树中对微生物部分的认知。
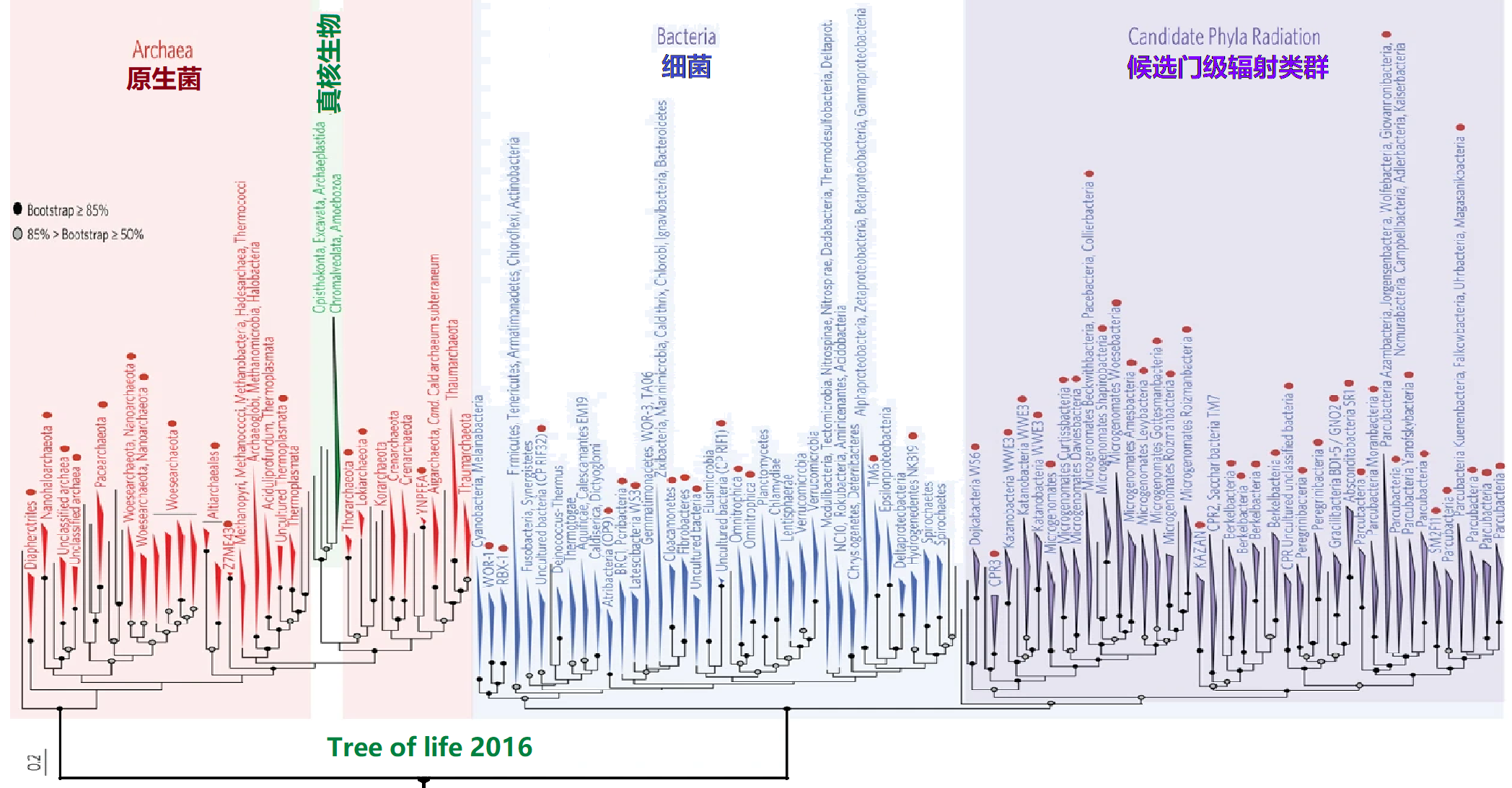
图2 2016年生物界系统发育三域树的改进版
2021年,给出了最新的生命树,总体的效果可以用如图3的树状图形来所示。图3是一张让人惊叹的生命之树,这张图上绘制了两百多万种物种之间的复杂关系,图中的每一个分支的尖端都代表一个不同的物种,而这些树枝追溯不同物种是如何从共同的祖先进化而来并沿着树枝如何向前发展,这张图最让你惊讶的地方有两个:(1)枝叶如此之多,表现出地球上生命的多样性是如此地丰富;(2) 树枝如此之复杂,表现了地球上生命之间的进化关系是如此地复杂。
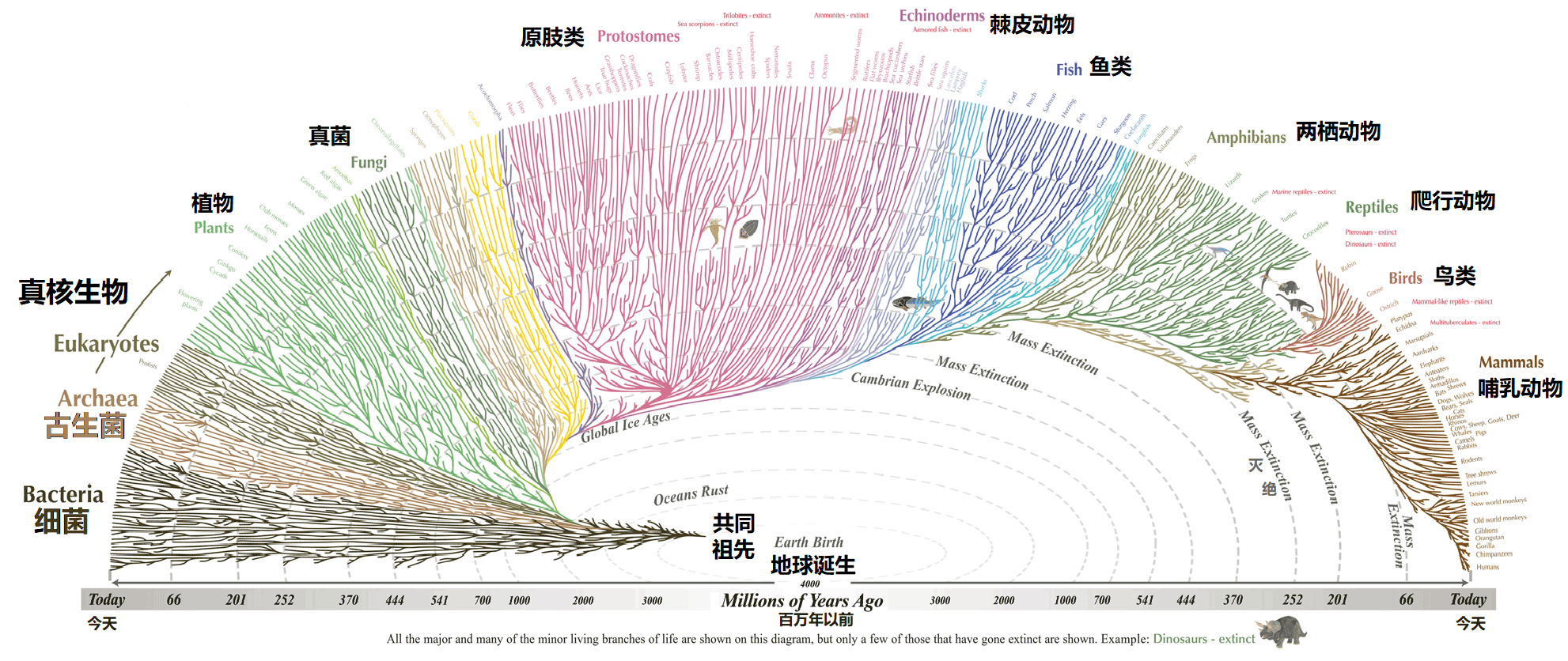
图3 复杂的生命之树
2. 生命树的发展历程
生命树模型可以非常清晰地展示地球上的每一种生物都是从大约35亿年前存在的一个共同祖先不断发展进化而来的。因为所有的生物都来自同一个祖先,所以它们存在一些共同的基因序列。随着祖先及其后代的不断繁衍,生命在一个被称为物种形成的过程中不断分化,这些分化就称为单一进化树枝上的分叉。随着达尔文的《物种起源》和恩斯特·海克尔对进化树的描绘,生命之树作为一种模型、隐喻或启发式的图像,在系统发育学中逐渐占据了重要的基础地位。
系统发育学是一门研究不同生物群体如何相互联系的学科,它是利用遗传学和其他证据来创建和解释进化史的图表,称为系统发育树(或生命树)。20世纪六七十年代兴起的系统遗传学,一直遵循着达尔文的树模型:生命树的分支是分开的,从来没有合并过。起初系统发育学以解剖学和胚胎学的机体特征为划分标准,并由此给出了动植物的分类树标准(就是把生物分成了等级森严、互不重叠的组,而种嵌在属里,属嵌在科里,科嵌在目里等等)。随后出现的分子测序方法彻底改变了系统发育学的发展。在20世纪60年代,对脊椎动物血红蛋白的比较研究使Zuckerkandl和Pauling提出,蛋白质的氨基酸序列以相对恒定的速率变化,可以用来估计物种进化分化的时间。他们提出的“分子钟”假说为分子系统发育学奠定了基础。要建立分子系统发育树,首先要对同源的核酸或蛋白质序列进行比对,比如给定一个描述氨基酸、RNA或DNA序列随时间变化的模型,人们就可以推断出一个代表所有序列之间关系的树。此后不久,随着RNA和DNA测序技术的发展,卡尔·沃斯(Carl Woese)提出以核糖体小亚基RNA基因序列为基础来构建生命之树的方法,开创了微生物系统发育的新时代。沃斯的方法揭开了生物学家了解甚少的微生物的进化史,以及揭示了真核生物中线粒体和质体的内共生起源,这导致了1990年沃斯及其同事在基于rRNA序列比较的基础上提出了生命树的基础模型:三域树(见图4),他将生命分为三个领域:细菌,真核生物和古生菌(新发现的谱系)。直到今天,三域树仍然是生命进化史中生物关系的标准形象。
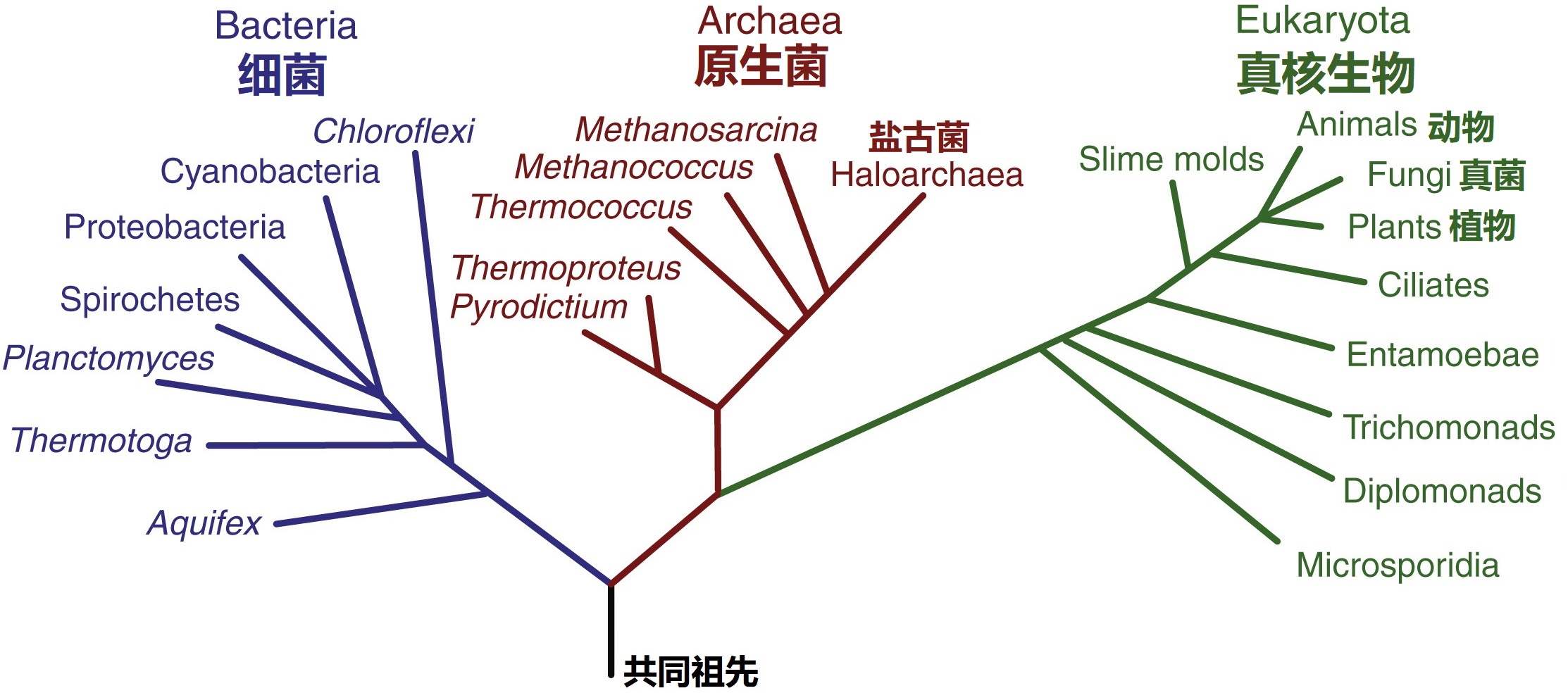
图4 三域树模型:细菌、古生菌和真核生物
随后在20世纪90年代末,随着全基因组测序和比较基因组学的兴起,人们依靠已测序的基因作为谱系的标记,很快发现同一基因组中的不同基因之间非常不同的树拓扑结构,甚至发现密切相关的生物体在基因含量上存在很大差异。比较基因组技术在原核生物中发现了广泛的横向基因转移,这是一种新的遗传进化现象:横向基因转移(HGT),即不同物种之间的遗传物质的交换过程(Nat. Ecol. Evol. 2: 904, 2018)。虽然生物体从父母那里继承了大部分基因,但在进化过程中,生物体可通过横向基因转移的过程与邻居交换基因而获得新的基因。基于基因转移的基因树与普通物种树的结构开始发生矛盾,因为不同物种的基因可以通过不同的进化途径进入生物体的基因组。虽然20世纪50年代以来,细菌中的种间重组就已为人所知,但当时没有大量的序列数据而无法完全了解其对系统发育的影响。然而广泛存在的转移基因的发现让树状生命树出现问题,基于此一些生物学家提出了生命进化史的另一种通用图形:生命网络。
根据网络模型,基因和基因组可以不断地交换和洗牌,它们的历史将交织在一个网状的网络中,无法清晰重建垂直树的图像。围绕横向基因转移对生命之树的影响,生命树结构的争论越来越激烈。反对生命树的人认为,基因的横向转移可能影响现存基因组中的所有基因,所以生命进化的树状结构不成立;而支持生命树的人则认为,基因横向转移的数量和比例并非是基因进化的主体,其影响垂直进化的程度不是很大。但无论如何,横向基因转移的存在改写了真正的、普遍的生命之树,但由于目前依然缺乏检测横向基因转移的准确方法,而且对于如何最优识别转移基因,仍然没有普遍的共识,这使得这场争论一时难以结束。
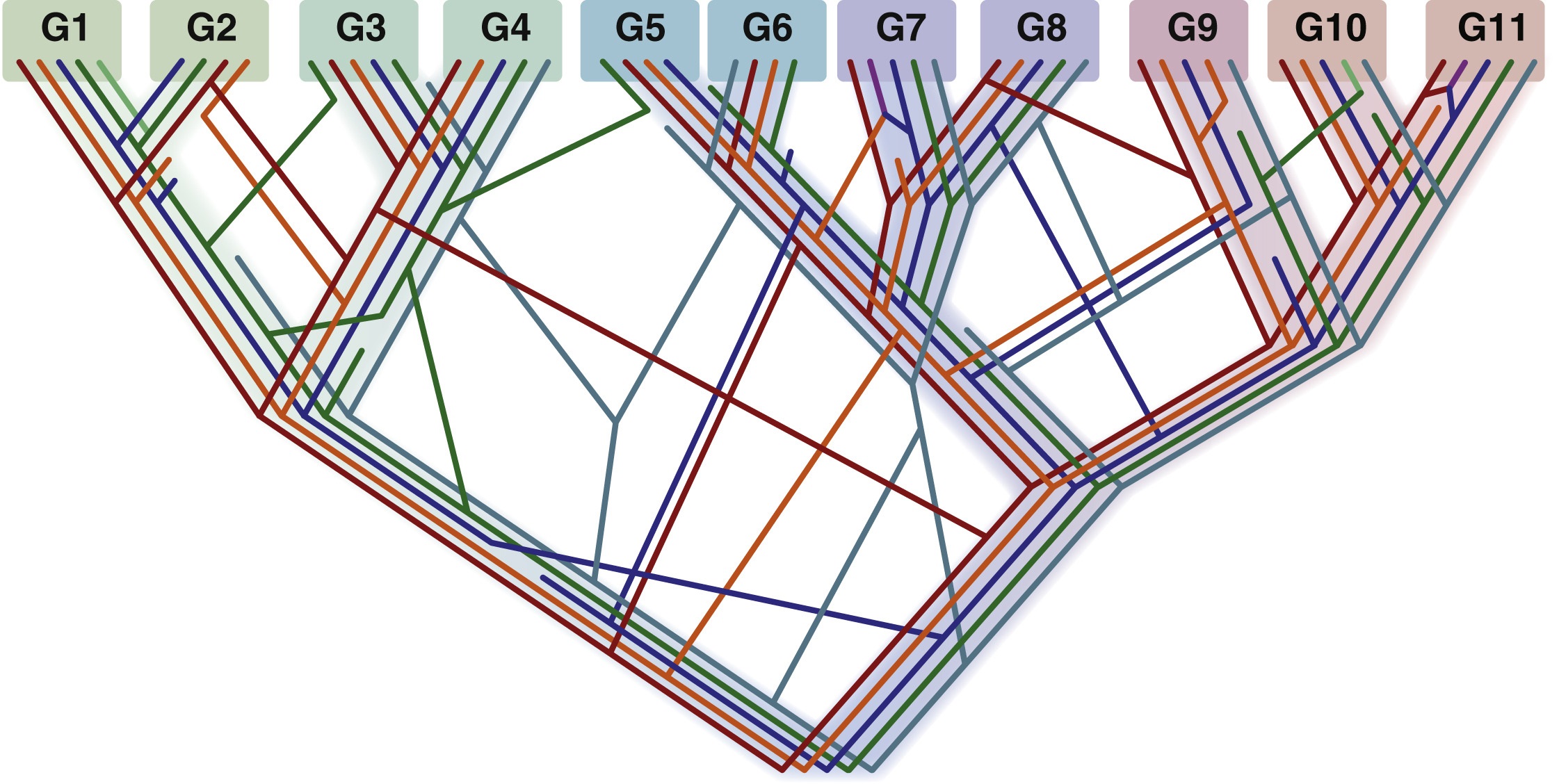
图5 生命之树的统计树模型
为了调和树和网的冲突,生命统计树模型认为树状垂直系统发育可能在相互冲突的基因树中表现为一种统计趋势(如图5所示)。图5中的彩色线条代表个体普遍的进化树特征,框代表现存物种的基因组(G)。虽然没有单一的基因树或几乎通用的树能捕捉到“真正的”生命之树,但它们的总和产生了一个清晰的树型结构,即生命进化的关系在统计的意义上体现了树的特征。
3. 生命进化的树网之争
生命树作为一种基础模型在系统发育学中发挥过重要作用,但转移基因的发现让有些人否认生命树的存在,他们认为生命之间的关系应该是一个交联的网络,使用树作为模型就预先假定了一种特殊的数据建模方法,这种图像对系统发育分析有严重的限制。但持这种观点的人基本上都不会彻底否定生物存在的进化史(生命树有根和垂直演化的枝条),生物界的进化过程都涉及遗传信息的传递,包括从亲代到后代的垂直继承,也包括所谓的水平传递,如通过杂交、重组、基因渗入、基因转移和基因组融合发生水平转移,所以与物种形成相关的垂直继承构造了一个树形的有机体进化史,而其他过程则在树中创造了网状结构。所以生命树基本上可以看成是一组相互连接的链,而网络是一组相互连接的树。而生命树到底是树状的还是网状的,这取决于垂直和水平遗传之间的平衡。从进化的角度来看,链代表了带有修饰的血统,是一个分类单元与其祖先之间的单一路径关系。树状结构代表这些链之间的相互关联,因为不同的物种形成是从任何给定的祖先开始,创造出许多可能的时间前进路径(这些链在时间上向后结合,存在遗传隔离)。而网络结构代表了这些树之间的相互联系,因为每个基因片段都有自己的谱系树,水平进化过程(杂交、重组、基因转移和基因渗入)在任何一个基因组的树中能创造许多可能的连接路径。因此,所有的系统发育严格来讲都是网络,但如果没有网状,那么任何给定的系统发育都将是树,如果没有物种形成,那么它也将只是链。所以生命树应该具有网络结构(如图6所示),而链和树都应该看成是网络的特殊情况。
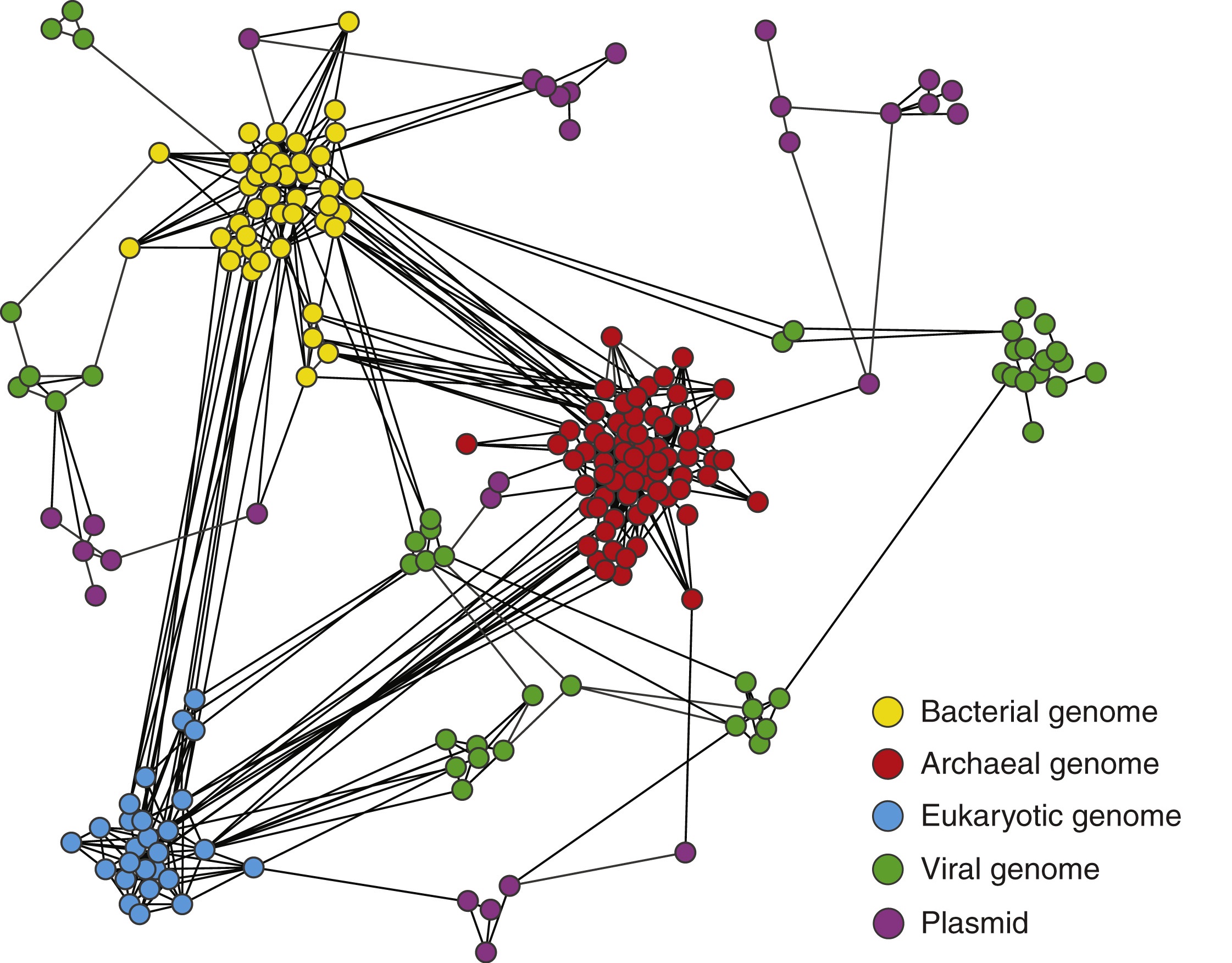
图6 基因组相关性所展示的生命网络结构
图6中的节点代表基因组,如果两个节点共享同源DNA序列,则之间由线连接在一起。细胞生命形式(古生菌、细菌和真核生物)和病毒的基因组,以及质粒和其他可移动的遗传元素,会聚集成离散的遗传世界,由世界内频繁的横向和纵向基因遗传所界定。这些区域之间的联系将整个生物界组织成了一个包罗万象的网络。分子系统遗传学发现细菌和古细菌的基因组在本质上是镶嵌的,它们通过横向(或水平)基因转移获得了来自近亲和远亲分类群的基因,通过原核生物中广泛的横向基因转移,关于生命作为一个整体是用树还是网络来代表已经达成了一个普遍的共识,即树和网络是共存的(Curr. Biol. 31(7): R314-R321, 2021)。数学上的树是一个无环路的连通图,而网络本质上与树具有相同的数学定义,它是存在环路的联通图。总体而言生命之树仍然表达了生物进化史的一个重要特征,垂直的系统发育一直而且仍然是一个实用和稳定的分类基础,在很大程度上由于基因横向转移所固有的多样性,形成了与生命树不相容的环状模式,将树整合成更广泛的相互作用网络,因此,将纵向和横向模式更广泛地统一到一个网络中,网络可以作为一种普遍理论的基础,将从分子到生物圈的生物相互作用的各个层面联系起来。
4. 生命树的结论
基因组规模的数据和新的统计系统发育方法的发展极大地帮助了生命之树草图的重建,并解决了生命树之中的许多分支问题。然而,在系统基因组学数据中相互冲突的进化史推断仍然是普遍存在的,这些问题阻碍了生物学家重建和解释生命树整体结构的能力。不完全谱系分选、基因水平转移、杂交、基因渗入、重组和趋同分子进化等生物学因素可导致不同于物种树的基因系统发育过程。在微生物世界中,细胞分裂、基因组复制和基因进化所导致的垂直遗传产生了树状结构,而细胞融合、基因融合和基因横向转移则引入了网状结构。所以某种基因元素通过细胞繁殖一旦整合到基因组中,会沿着树枝垂直遗传,直到发生转移,然后转移的基因继续进行垂直遗传,并再次发生转移。因此,生命树依然是存在的,而树之中存在的网状结构可以理解为发生在由垂直遗传过程决定的树枝上所产生的横向遗传支架,所以水平基因组学的概念是以垂直过程为前提的,纵向遗传和横向遗传应被理解为生物遗传进化的两种不同策略,它们共同导致了生物界无与伦比的适应性和复杂性。
https://blog.sciencenet.cn/blog-318012-1447607.html
上一篇:我们不知道答案的125个科学问题(96)性取向
下一篇:我们不知道答案的125个科学问题(98)物种的数量
