博文
氢气缺乏体质之久坐中年代谢综合征
||
研究表明:氢气缺乏体质之久坐中年代谢综合征
摘要
背景:代谢综合征与细胞生物能量学早期损伤相关,传统体成分测量无法完全反映这些损伤。氢气由肠道微生物发酵内源性产生,可通过呼气检测,参与氧化还原与线粒体调控。呼气氢水平是否与代谢综合征患者细胞内代谢活性组织的保留相关,目前尚不明确。
目的:探讨久坐代谢综合征中年人群的呼气氢浓度,与基于细胞内体成分指标构建的综合细胞生物能量学表型之间的关联性。
方法:纳入28名久坐中年代谢综合征受试者(平均年龄51.2±7.9岁,女性19名),完成空腹呼气氢检测与多频生物电阻抗分析。通过主成分分析,整合体细胞质量、细胞内水分、全身总钾与糖原,构建复合细胞生物能量学表型。采用斯皮尔曼相关(带自助法置信区间)、泰尔-森回归、贝叶斯线性回归(校正年龄、性别、腰围),分析呼气氢与复合表型的关联;敏感性分析纳入去脂体重。
结果:单一主成分可解释细胞内指标98.6%的变异,提示高度一致的细胞生物能量学表型。呼气氢浓度与该表型呈正相关(ρ=0.43,P=0.021;BCa 95%CI 0.07~0.70)。泰尔-森回归证实稳健正相关(β=0.017/ppm氢;95%CI 0.002~0.046)。贝叶斯模型后验分布集中于正效应量,且独立于中心性肥胖。与之相对,呼气氢仅与去脂体重呈临界相关。
结论:在久坐代谢综合征中年人群中,呼气氢浓度可反映综合细胞内生物能量学表型,追踪细胞质量而非瘦体重数量。呼气氢可作为细胞生物能量学完整性的无创生物标志物,为表型导向的代谢干预提供潜在工具。
---
1. 引言
代谢综合征以中心性肥胖、胰岛素抵抗、血脂异常、低度全身炎症并存为特征,伴随多器官系统代谢健康进行性恶化[1]。除脂肪过量外,代谢综合征患者常出现细胞生物能量学损伤,包括线粒体功能障碍[2]、氧化还原失衡[3]、细胞内离子稳态紊乱[4]、底物储存能力下降[5]。这些异常会导致代谢活性组织质量与功能受损,即便无明显体重下降或肌肉减少症也可发生。
代谢综合征的体成分改变不仅限于去脂体重减少。新证据显示,早期改变优先影响细胞内代谢活性区室,其生化与分子变化早于临床症状[6],表现为体细胞质量、细胞内水分与钾含量降低[7],同时糖原储存受损[8]。这些特征反映细胞完整性与水化状态下降,可先于并可能驱动后续收缩组织丢失[9]。因此,能同时反映细胞内质量、水化、矿物质含量与能量储存的整合标志物,比单纯瘦体重粗测指标更能敏感反映代谢健康。此前尚无研究探讨内源性氢与代谢综合征综合细胞内生物能量学表型的关系。
氢气作为具有代谢调控潜力的生物活性气体,近年备受关注[10,11]。内源性氢气主要由肠道菌群发酵膳食底物产生,部分进入体循环并经呼气排出[12]。基础与临床研究表明,氢气可调控氧化还原稳态与线粒体功能,且不干扰生理信号传导[13,14],具备成为细胞生物能量学状态标志物或介导因子的潜力。呼气氢浓度可无创反映内源性氢生成与可利用度[15],但其与人体体成分、细胞代谢完整性及生物能量学的关系仍不明确。
在久坐代谢综合征人群中,呼气氢差异不太可能反映体力活动或运动诱导的肌肉肥大差异。相反,内源性氢生成差异可能与影响细胞水化、细胞内离子平衡及底物储存的基础代谢与菌群表型相关[16]。目前尚未系统研究呼气氢是否与该人群代谢活性细胞区室的保留相关。
据此,本研究旨在探讨久坐中年代谢综合征人群的呼气氢浓度,与多频生物电阻抗分析所得细胞内完整性指标的关联性。为捕捉体细胞质量、细胞内水分、全身总钾与糖原的共同生物学变异,采用主成分分析构建复合细胞生物能量学表型。我们假设:更高的呼气氢浓度与该整合细胞表型呈正相关,反映代谢活性水化细胞质量的保留,且独立于总体体型。本研究聚焦复合细胞内表型而非孤立体成分指标,旨在从机制层面揭示代谢综合征中内源性氢与细胞代谢健康的关系。
---
2. 研究方法
2.1 研究设计与受试者
本观察性研究纳入久坐中年代谢综合征人群。代谢综合征诊断依据公认标准,满足以下5项中至少3项:男性腰围≥102 cm、女性≥88 cm;甘油三酯升高(≥1.695 mmol/L);高密度脂蛋白胆固醇降低(男性<1.04 mmol/L、女性<1.30 mmol/L);血压升高(>130/85 mmHg);空腹血糖升高(>6.1 mmol/L)。纳入标准还包括久坐生活方式、签署书面知情同意书。久坐定义为未参与规律体育活动或运动训练,且评估前至少3个月体重稳定。排除标准:急慢性炎症性疾病、已知影响气体生成/吸收的胃肠道疾病、近期使用抗生素、吸烟、使用影响体内水分分布或肠道发酵的药物/补充剂。受试者通过社区招募(公开广告与外展活动),无临床转诊病例。所有受试者入组前签署书面知情同意。研究方案经机构伦理委员会批准(编号ABE-2025-05-383-13-02),遵循《赫尔辛基宣言》实施。
2.2 研究流程
所有测量在单次访视、标准化条件下完成。受试者需隔夜空腹(≥10小时),检测前24小时禁酒、避免剧烈运动,前一日避免可发酵食物。到达后排空膀胱,安静休息后开始测量。采用电化学燃料电池微处理器(LactoFAN2)检测空腹呼气氢浓度,评估内源性氢生成。按标准化呼气末采样流程坐位采集呼气样本:正常吸气、短暂屏气、完全呼气至采集装置。呼气氢浓度以百万分比(ppm)表示,所有分析均作为连续变量处理。尽管膳食纤维摄入是结肠氢生成的主要决定因素[17],但本研究未量化日常纤维摄入,可能导致呼气氢浓度的个体间差异。采用多频生物电阻抗分析(BIA)系统(BioScan 920)评估体成分,按厂家指南标准电极放置,仰卧位测量。设备施加多频电流(5 kHz、50 kHz、100 kHz、200 kHz),依据组织阻抗特性估算体成分与体液区室。所有BIA指标均按设备专有算法以绝对单位输出。重点关注体细胞质量(BCM)、细胞内水分(ICW)、全身总钾(TBK)与糖原,作为代谢活性细胞组织的指标。生物电阻抗估算的糖原为基于水化算法的间接指标,非直接生化检测结果。
2.3 细胞生物能量学表型构建
为独立于总体体型表征细胞内代谢活性区室,从BIA参数中构建复合细胞生物能量学表型,覆盖细胞完整性的互补维度。BCM主要反映代谢活性细胞数量,ICW反映细胞水化与体积依赖的代谢能力,TBK作为细胞内离子与酶质量的替代指标,糖原代表细胞内能量缓冲与代谢灵活性。这些变量为预先选定,并非仅反映体成分单一维度,而是整合细胞数量、水化状态与能量储备——即细胞内生物能量学质量的核心特征。主要复合指标排除去脂体重,以避免总体体型相关的非特异性影响。对标准化(Z分数)的BCM、ICW、TBK与糖原,基于相关矩阵进行主成分分析(PCA)。按Kaiser准则(特征值>1.0)与生物学可解释性(载荷一致正向、符合细胞结构与功能)保留主成分。第一主成分(PC1)捕捉上述维度的共同变异,定义为细胞生物能量学表型。采用回归法计算个体PC1得分,用于后续分析。为评估观察到的关联是否主要由瘦体重数量而非细胞内质量驱动,进行敏感性分析:(1)纳入去脂体重重复PCA;(2)单独分析呼气氢与去脂体重的关联。各分析结果一致,提示结果并非仅由体型驱动。收集生化变量以描述队列代谢特征,不纳入关联分析,以减少多重检验、聚焦体成分衍生细胞表型。
2.4 统计分析
鉴于样本量有限,统计分析侧重效应量估计、稳健性与生物学一致性,而非依赖零假设显著性检验。刻意限制模型复杂度以避免过拟合。通过可视化与描述性统计检查数据分布。采用斯皮尔曼秩相关分析呼气氢浓度与单项体成分参数、衍生细胞生物能量学得分的关联。为获得对异常值不敏感的稳健线性关系估计,额外进行泰尔-森回归。为进一步量化关联并校正关键混杂因素,构建贝叶斯线性回归模型,以细胞生物能量学得分为因变量、呼气氢浓度为主要预测因子。模型校正年龄、性别,敏感性分析额外校正腰围。为回归系数指定弱信息正态先验,结果以后验均值与95%可信区间呈现。对相关与回归估计采用非参数自助重抽样(5000次迭代)估计效应量的不确定性。图表采用散点图+局部加权平滑(LOESS)展示呼气氢与细胞结局的剂量反应趋势。为在贝叶斯框架下进一步量化呼气氢与细胞生物能量学表型关联的证据强度,采用Savage-Dickey密度比计算氢系数的贝叶斯因子。该方法比较零值(β=0)处的先验密度与对应后验密度,连续衡量支持备择假设(β≠0)而非零假设的证据强度。贝叶斯因子计算采用与贝叶斯模型中标准化回归系数相同的弱信息正态(0,1)先验。证据强度按通用阈值解读(BF10≈1~3:轶事证据;3~10:中等证据;>10:强证据)。所有分析采用IBM SPSS Statistics 24.0与R 4.0.0(Posit Cloud)完成。适用时采用双侧检验,但解读侧重一致性与生物学合理性。本研究为探索性研究,结果结合生物学合理性与各分析方法的一致性解读,而非严格统计显著性。
---
3. 研究结果
最终纳入28名久坐代谢综合征受试者(男性9名、女性19名),平均年龄51.2±7.9岁。空腹呼气氢平均浓度26.0±23.9 ppm(中位数19.5;四分位距7.5~38.0),平均腰围98.9±15.2 cm。体细胞质量、细胞内水分、全身总钾与糖原高度相关,合并为单一细胞生物能量学表型(PC1),呈近似正态分布(均值0.00±1.00)。研究人群描述性特征见表1。
表1 研究人群描述性特征(n=28)
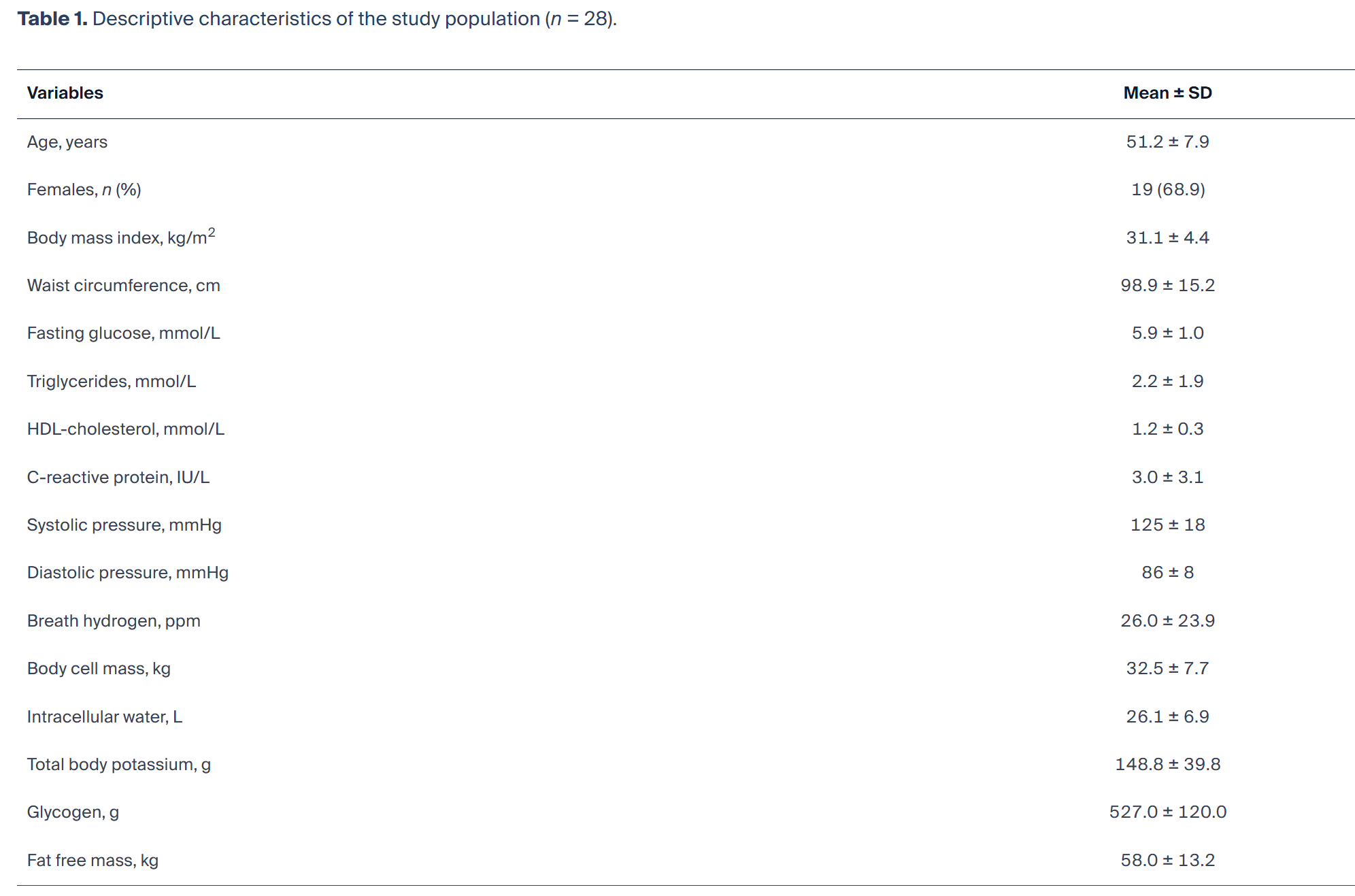
对BCM、ICW、TBK与糖原的PCA识别出单一主导成分,特征值3.94,解释总变异98.6%。所有变量在PC1上均呈强正向载荷(标准化载荷0.983~0.999),公因子方差均较高(0.966~0.999),提示各变量几乎全部变异均被复合得分捕捉。无其他成分满足保留标准。结果表明,细胞内质量、水化、钾含量与糖原储存构成高度一致的潜在结构,符合整合细胞生物能量学表型。纳入去脂体重重复PCA后,第一主成分仍占主导(特征值4.93),解释总变异98.6%,所有变量载荷同样强且正向。呼气氢与复合表型的关联在方向与量级上与主要分析可比,提示结果并非仅由总体瘦体重驱动。
呼气氢浓度与衍生细胞生物能量学表型得分呈正相关(ρ=0.43,P=0.021)。自助法偏差校正加速置信区间证实关联稳健(BCa 95%CI 0.07~0.70;5000次自助抽样)。该关联提示更高的内源性氢水平与更优的整合细胞质量、细胞内水分、钾含量及糖原储存相关。关联方向与量级在各单项成分变量中一致,支持呼气氢反映共同细胞内生物能量学核心,而非孤立体成分参数。与上述结果一致,泰尔-森回归显示呼气氢浓度与细胞生物能量学表型呈正相关(β=0.017/ppm氢,95%CI 0.002~0.046),置信区间不包含零,证实对潜在异常值稳健。相比之下,呼气氢仅与去脂体重呈临界相关(P=0.05),提示氢与细胞内生物能量学质量的关联强于瘦体重数量。
校正年龄、性别的贝叶斯线性回归模型显示,呼气氢浓度与细胞生物能量学表型呈正相关(后验中位数β=0.27,95%可信区间-0.13~0.67)(图1)。尽管可信区间跨越零,但后验分布集中于正效应量,与非参数及稳健回归分析一致。性别与细胞生物能量学表型呈正相关,年龄无独立关联。额外校正腰围的敏感性分析中,呼气氢与细胞生物能量学表型的关联方向保持一致(后验中位数β=0.24,95%可信区间-0.16~0.65),提示观察到的关系并非由中心性肥胖驱动。综上,贝叶斯结果支持呼气氢浓度与细胞生物能量学状态存在稳定正相关,同时合理反映小样本带来的不确定性。各分析框架下,呼气氢均持续追踪同一细胞内生物能量学维度。
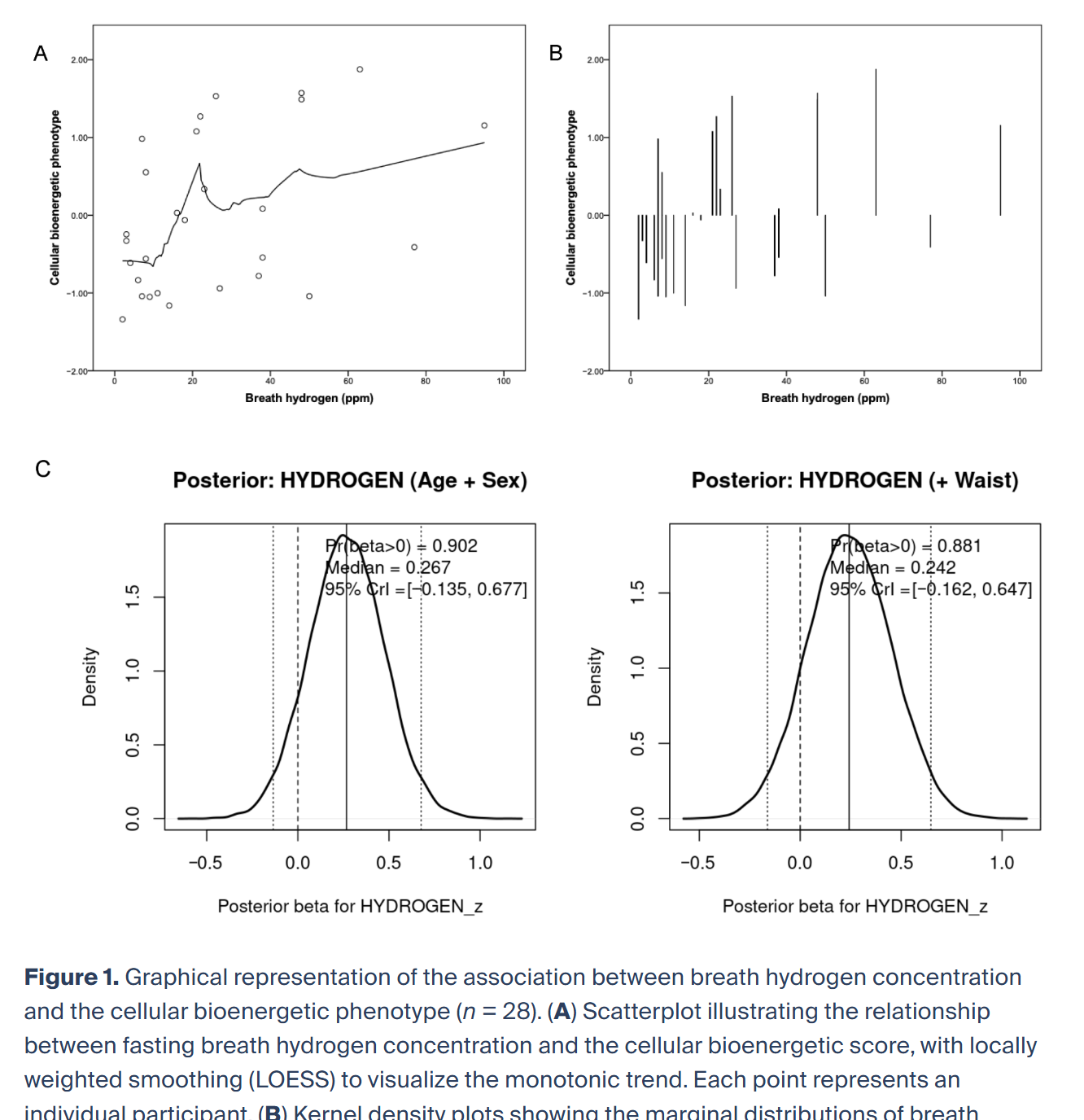
图1 呼气氢浓度与细胞生物能量学表型关联图示(n=28)
(A)空腹呼气氢浓度与细胞生物能量学得分的散点图,局部加权平滑(LOESS)展示单调趋势,每个点代表1名受试者。
(B)队列中呼气氢浓度与细胞生物能量学表型的边际分布核密度图。
(C)贝叶斯线性回归标准化氢回归系数的后验概率密度。左图为校正年龄、性别的后验分布,右图为额外校正腰围的后验分布。密度曲线经缩放,曲线下总面积为1。垂直实线为后验中位数,虚线为95%可信区间,点虚线为零值(β=0)。插图报告氢效应为正的后验概率(Pr[β>0])。两模型后验分布均集中于正值,提示呼气氢与细胞生物能量学表型呈正相关的后验概率较高。
贝叶斯因子分析显示,区分氢非零效应与零假设的证据有限。采用正态(0,1)先验的Savage-Dickey法,校正年龄、性别的模型BF10≈0.49,额外校正腰围的模型BF10≈0.41。该数值对应轶事证据,略支持零假设而非非零效应。因此,尽管后验分布集中于正效应量,且两模型中正相关概率均>0.88,但数据对竞争假设的区分证据有限,反映小样本固有的不确定性。
---
4. 讨论
本项针对久坐中年代谢综合征人群的观察性初步研究显示,呼气氢浓度与基于体细胞质量、细胞内水分、全身总钾及糖原构建的综合细胞生物能量学表型持续呈正相关。主要复合指标排除去脂体重,刻意构建以反映细胞内代谢活性组织特性,而非总体体型或瘦体重数量的非特异性变异。细胞内质量、水化、离子含量与糖原储存几乎完全汇聚为单一主导主成分(解释这些变量几乎全部变异),凸显这些细胞属性的强生物学相互依赖性,支持将其概念化为统一的生物能量学结构。
这种一致性在代谢综合征背景下尤为重要,该疾病中细胞水化与细胞内能量储存损伤常先于或伴随功能性去脂体重与代谢灵活性下降[6,18,19]。细胞内水分与钾含量丢失与细胞体积依赖的合成代谢信号减弱、线粒体功能受损相关[20],糖原储存能力下降则反映底物处理与能量缓冲受损[21]。因此,高度整合的细胞内表型可捕捉仅靠传统人体测量或生化标志物难以发现的代谢健康维度。复合得分的近似正态分布,进一步支持其作为该人群细胞生物能量学状态定量描述指标的适用性。
呼气氢浓度与细胞生物能量学表型的正相关提示,更高的内源性氢生成可追踪代谢应激状态下细胞完整性的保留。氢气被认为可影响氧化还原信号、线粒体效率与炎症状态[22,23],这些机制可合理支持细胞内钾梯度、细胞水化与糖原储存能力的维持。尽管本数据无法推断直接机制通路,但氢水平与多项细胞内指标的一致性,支持内源性氢反映或促进更具韧性的细胞生物能量学环境的假说。从转化角度看,本结果提示内源性氢生成低的代谢综合征患者,可能代表细胞生物能量学韧性降低的生物学亚组。此类患者补充外源性氢(最常见为富氢水或产氢化合物[24]),可成为支持细胞内生物能量学功能的靶向策略。
基础与早期临床研究显示,外源性氢可短暂提高全身氢可利用度,调控氧化还原敏感信号通路[25,26],可能模拟或增强内源性氢介导效应。若内源性氢可作为细胞生物能量学完整性标志物,那么在低氢表型中恢复氢可利用度,或有助于稳定代谢应激下的细胞水化、离子稳态与能量储存能力。重要的是,该框架不意味着外源性氢可直接增加瘦体重或逆转既定代谢综合征,而是提示其在支持代谢活性细胞区室的质量与功能完整性方面的作用,尤其在内源性氢生成受损的久坐人群中。与该解读一致,呼气氢仅与去脂体重呈临界相关,而与细胞内生物能量学指标的关联在各分析方法中更强、更稳定。因此,按基线呼气氢分层,可提高未来干预研究的精准度,评估低内源性氢个体是否从氢补充中获得更显著的生物能量学获益。该方法符合表型导向营养与代谢干预的新兴理念[27],或有助于解释既往氢相关试验中观察到的反应异质性。综上,本结果支持呼气氢不仅可作为细胞生物能量学状态的无创生物标志物,还可用于识别最可能从氢靶向干预中获益的个体。未来需开展随机对照试验,结合基线氢分层、细胞内生物能量学标志物纵向评估与线粒体功能直接检测,明确外源性氢是否可因果影响代谢综合征及类似疾病的细胞生物能量学完整性。
实验模型为这些关联提供了合理的机制基础。体外与动物研究显示,氢气选择性清除高活性氧化剂(尤其是羟自由基与过氧亚硝酸盐),减轻氧化损伤且不干扰生理氧化还原信号[14,28]。除直接抗氧化作用外,氢调控氧化还原敏感信号级联,包括激活Nrf2-Keap1通路、上调血红素氧合酶-1、抑制NF-κB依赖的炎症转录及类氢化酶线粒体氧化还原蛋白[29,30,31,32]。氢暴露还可提高线粒体膜电位、维持电子传递链复合物活性、改善ATP合成效率,部分通过稳定复合物I与V、减少ROS诱导的解偶联实现[22,33]。这些作用伴随细胞水化与离子稳态改善,与氢介导的Na+/K+-ATP酶活性及线粒体离子转运维持一致。在此背景下,本研究识别的整合表型(涵盖细胞内质量、水分、钾含量与糖原),可能代表此类线粒体与氧化还原调控过程的下游系统水平表现。因此,人体中呼气氢与该表型的关联,与基础研究中氢在应激状态下支持细胞能量效率、氧化还原平衡与代谢灵活性的结论一致。
mTOR信号提供了额外机制层面解释,该信号整合营养可利用度、细胞水化、氨基酸通量与能量状态,协调细胞生长、糖原代谢与代谢稳态。近期研究表明,mTOR复合物与细胞内离子平衡、线粒体效率紧密耦合,氨基酸依赖的ATP合酶组装与氧化磷酸化能力调控细胞生物能量学状态[34,35,36]。代谢疾病中mTOR信号紊乱,破坏糖原处理、细胞体积依赖的合成代谢信号与线粒体偶联,导致细胞内质量、水化与能量储备降低的表型。在此框架下,本研究识别的复合表型,可能反映mTOR调控的生物能量学通路的下游表现。因此,呼气氢与该表型的关联,符合内源性氢追踪或调控氧化还原敏感mTOR-线粒体偶联、维持应激状态下细胞内代谢完整性的模型。
重要的是,上述关系在非运动久坐人群中观察到,最大程度降低体力活动诱导的肥大或训练适应解释结果的可能性。将去脂体重纳入主成分的敏感性分析,得到几乎相同的潜在结构与呼气氢关联,提示结果并非仅由总体瘦体重更高驱动。相反,呼气氢似乎与代谢活性细胞区室的质量与完整性相关(涵盖水化状态、离子含量与能量储存),而非单纯瘦体重数量。
关联在多种分析框架下的稳健性,增强了该解读的可信度。采用带自助法置信区间的非参数相关,呼气氢与细胞生物能量学表型呈正相关;稳健泰尔-森回归中关联仍显著,置信区间不包含零。贝叶斯线性回归进一步显示,氢效应的后验分布持续集中于正值,即便校正年龄、性别与腰围,正相关的后验概率仍较高。尽管存在不确定性(小样本观察性研究预期内),频率法、稳健法与贝叶斯法的结果趋同,支持真实生物学梯度存在,而非虚假或异常值驱动效应。贝叶斯因子结果进一步 contextualize 这些发现。尽管后验分布集中于正效应量,贝叶斯因子仅显示轶事证据区分氢非零效应与零假设。该模式是小样本探索性研究的特征,效应估计方向稳定但精度不足。重要的是,这并不与各分析框架的一致性矛盾;反而强调本数据支持合理且生物学一致的关联,同时提示需更大样本、足够效能的研究以增强推断效力。在此背景下,呼气氢应被视为有前景但暂定性的细胞生物能量学状态标志物,需未来干预与纵向设计验证。
近期哺乳动物线粒体基因调控的比较分析,为本研究结果提供额外机制背景。多项研究显示,氧化磷酸化核编码组分的表达与协调存在显著组织特异性差异,尤其在氨基酸代谢与ATP合酶组装调控通路中[37,38]。这些数据表明,线粒体生物能量学能力并非仅由细胞器数量决定,而是由精细调控程序驱动,以细胞类型特异性方式耦合底物可利用度、氧化还原状态与ATP生成机制。重要的是,连接氨基酸通量与线粒体效率的保守调控基序,可稳定代谢活性组织的细胞内能量平衡与细胞水化。在此框架下,本研究捕捉的高度整合表型(涵盖体细胞质量、细胞内水分、钾含量与糖原),可能反映此类保守线粒体调控架构的下游表现。该表型与呼气氢的关联提示,内源性氢生成可能追踪或功能耦合于人体这些细胞内生物能量学程序。因此,呼气氢可作为线粒体-细胞偶联机制完整性的系统水平替代指标,该机制可维持代谢应激状态下的细胞内代谢韧性。
本结果可置于内源性气体递质(如一氧化氮NO、一氧化碳CO)的广义生物学背景下解读,后者是线粒体呼吸、血管张力与代谢信号的公认调控因子(详见综述[39])。代谢疾病中,NO生物利用度紊乱与血红素氧合酶来源的CO,与内皮功能障碍、氧化磷酸化受损及底物利用改变相关[40,41]。然而,与NO、CO不同,氢气并非宿主酶合成,主要来自微生物发酵,因此对宿主-菌群代谢耦合高度敏感。此外,氢无过量NO或CO的血管毒性与细胞毒性,不直接抑制细胞色素氧化酶,可调控氧化还原平衡且不抑制线粒体呼吸。这些特性使氢成为细胞生物能量学状态的互补(且可能更优)标志物:整合菌群活性与宿主代谢韧性,可经呼气无创检测,反映全身氧化还原-线粒体偶联,无其他气体介质固有的血流动力学混杂效应。
本研究存在若干局限性。样本量有限,观察性设计无法推断因果关系。体成分与细胞内指标来自多频生物电阻抗分析,为间接估算而非细胞区室直接测量[42]。结肠氢生成的主要决定因素[17]——日常膳食纤维摄入未评估,也未进行菌群谱分析;两者均可能导致内源性氢生成的个体差异。此外,性别分布不均衡(女:男≈2:1),可能影响主成分结构与单项变量权重,但校正性别的敏感性分析结果方向一致。本研究未探讨呼气氢与单项生化标志物的关联,设计专注于体成分衍生细胞生物能量学指标,减少小样本多重检验。因此生化变量仅用于描述性表征,而非机制推断。最后,结果可能无法外推至久坐中年代谢综合征人群以外,对年轻、体力活跃或临床不同人群需谨慎解读。
尽管存在上述局限性,细胞生物能量学表型的强内部一致性,及其与呼气氢在各分析方法中关联的稳定性,支持其作为细胞内代谢完整性整合标志物的实用性。本结果为未来干预研究提供依据,包括按基线呼气氢水平分层的富氢水补充或膳食调控,以明确内源性或外源性氢是否可因果影响代谢综合征的细胞生物能量学状态与体成分。纳入线粒体功能、细胞内离子处理与组织特异性糖原动态直接检测的纵向设计,对阐明本观察提示的机制关联尤为重要。
---
5. 结论
在久坐中年代谢综合征人群中,呼气氢浓度与反映细胞内质量、水化、钾含量及糖原储存的综合细胞生物能量学表型呈正相关。本研究构建独立于总体瘦体重的复合表型,证实内源性氢与代谢活性细胞区室的质量及完整性更密切相关,而非单纯瘦体重数量。该细胞内表型的一致性,及其与呼气氢在非参数、稳健与贝叶斯分析框架下关联的稳定性,支持氢气作为细胞生物能量学状态标志物的生物学意义。本结果提示,内源性氢生成的差异,可能反映代谢应激状态下细胞代谢韧性的差异。尽管无法推断因果关系,结果为未来干预研究提供依据,探讨调控氢可利用度(尤其基线氢水平低的个体)是否可支持代谢综合征的细胞内生物能量学完整性。因此,呼气氢可作为无创生物标志物,为表型导向的代谢干预提供依据。
https://blog.sciencenet.cn/blog-41174-1530406.html
上一篇:氢气吸入对脑血流的影响大【新进展】
下一篇:空气中充满DNA——科学家正在用它做这些事
