博文
植物认知的范式革命——从神经中心主义到生态互动视角
|
像植物一样思考:行为生态学与植物认知本质的视角
【核心概览】本文打破传统认知研究的人类中心主义桎梏,基于认知生物学与生态心理学理论,论证认知是生物体与环境互动的动态过程。通过梳理植物在感知、学习、记忆、亲缘识别、空间定向及通讯等方面的实验证据,提出以"可供性"理论为核心的整合研究框架,重新定义植物作为行为主体的认知能力。
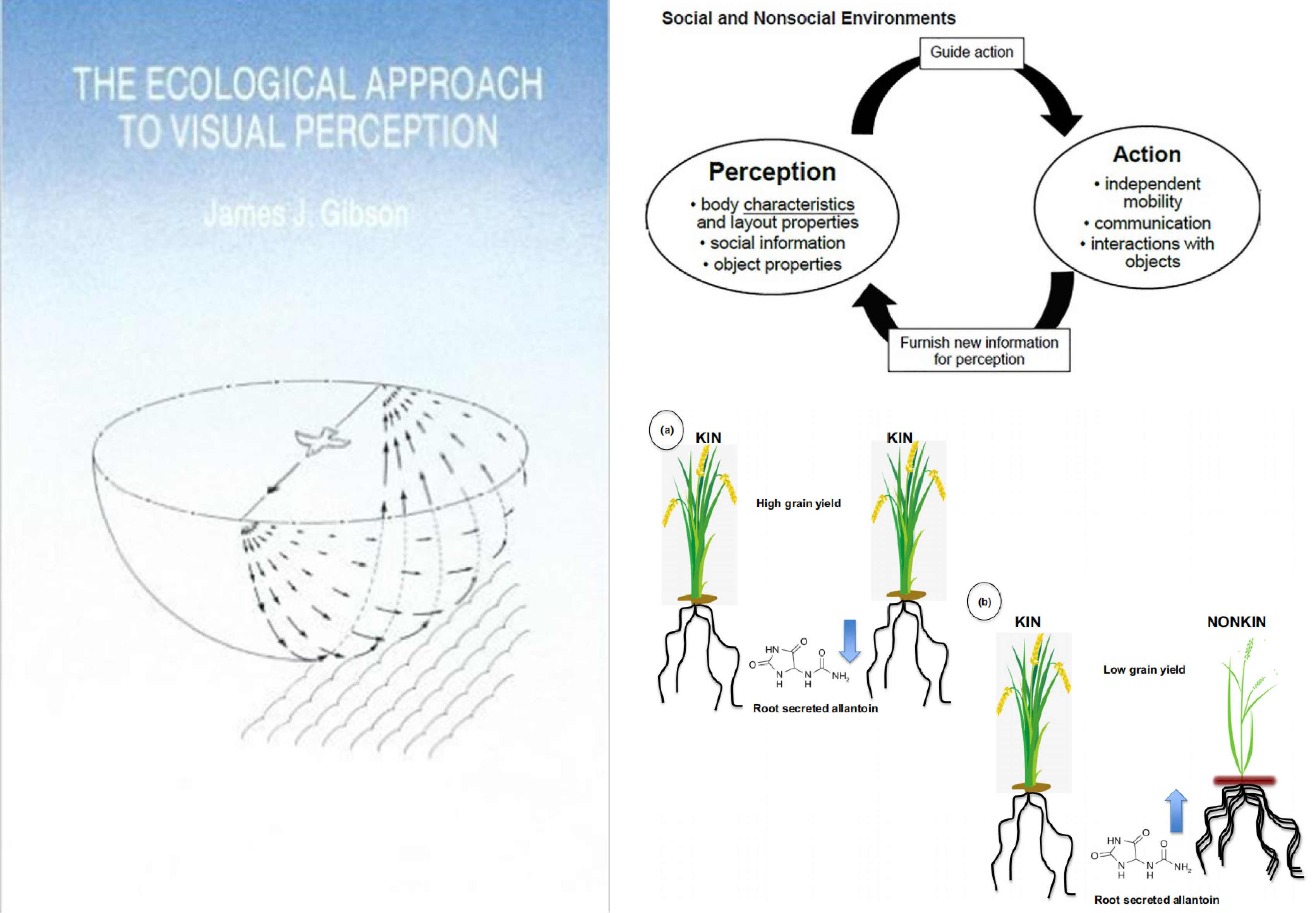
图1 认知研究范式转变示意图
左:传统神经中心主义认知模型(以大脑为核心);右上:吉布森生态心理学的感知-行动耦合系统;右下:水稻亲缘识别实验示意图(亲缘植株通过减少尿囊素分泌降低根系竞争)(Huang et al., 2018)
1 引言:感知与认知——生命系统的基本属性
感知环境并做出适应性响应是所有生命体的核心生存技能。生物体通过多样化的信号转导系统处理环境信息,这一过程远比简单的"刺激-反应"模式复杂——从细菌的趋化性到哺乳动物的决策行为,感知与认知能力构成了生命演化的重要驱动力。
感知作为认知的基础,不仅包含信息接收,更涉及对复杂环境信号的组织与解释。这种能力从根本上塑造生物体的行为策略,是生命系统区别于非生命物质的本质特征。演化压力促使生物体发展出与生态位相匹配的感知模式,以获取环境中的关键信息。
正如生物学家冯·于克斯屈尔(von Uexküll)提出的"环境界(Umwelt)"理论,每个物种都通过独特的感知滤镜构建主观生存世界。这种感知主观性意味着认知研究必须超越人类中心视角,才能真正理解其他生物的认知本质(Tacca, 2011)。
近25年来,认知科学逐渐突破人类中心框架,开始关注非人类生物的认知能力(Yoerg, 1991; Shettleworth, 2001)。从动物的数字认知(如计数、算术能力)到社会学习行为,研究表明认知能力广泛存在于生物界(Brannon & Roitman, 2003; Vallortigara et al., 2010)。值得注意的是,植物同样展现出复杂的社会学习能力,这为认知研究开辟了全新视角。
与试错学习等非社会学习不同,植物的社会学习通过感知邻近植株的信号实现环境信息高效传递。这种能力具有重要生态意义:通过亲属识别减少种内竞争(如水稻通过尿囊素调节根系生长);形成群体防御机制(如蚜虫侵害时释放挥发性信号);提升后代适应环境的能力,体现了分布式智能在植物界的独特表现。
从生命本质看,所有有机体都需通过感知-认知系统将环境信号转化为行为响应。植物认知研究的核心挑战在于:突破动物认知范式的桎梏,揭示无神经系统生物的认知机制,建立统一的生命认知理论框架。

图2 生物认知多样性示例:从昆虫到植物的社会学习策略
社会性昆虫的低成本认知策略(A-E):黄蜂面部识别、蜜蜂舞蹈通讯、白蚁集体筑巢等,展示了无复杂神经系统生物的智能行为。这些案例为理解植物认知提供了类比参考——植物虽无神经组织,却通过细胞信号网络实现类似的环境适应与群体协调(Lihoreau et al., 2012)。
2 植物认知研究的理论基础与范式突破
2.1 传统认知范式的人类中心主义局限
传统认知研究以人类大脑为模板,将神经系统视为认知的必要条件,认为复杂计算过程(预期、记忆、决策等)必须依赖神经组织。这种神经中心主义范式导致植物认知研究长期被忽视——因缺乏中枢神经系统,植物的适应性行为被简单归因为"被动响应"。
人类中心主义认知评估标准常陷入拟人化误区:通过类比人类行为解释非人类生物认知。神经影像学研究显示,人类大脑对类人动作的反应强于非人类动作(Oberman et al., 2007),这种认知偏差使得植物的缓慢行为与分散式信号处理难以被纳入认知研究范畴,成为植物认知研究的核心方法论障碍。
这种对类人类动作的认知依赖,导致那些以不同方式实现类似功能的物种被排除在外——尤其是缺乏神经系统且行动缓慢的植物,长期被认为不具备认知能力(Griffin, 1976; Warwick, 2000)。
2.2 有机体-环境互动视角:马图拉纳的认知生物学
突破人类中心主义的关键,是将认知重新定义为有机体与环境互动的动态过程。智利生物学家温贝托·马图拉纳(Humberto Maturana)提出,生物体与其生态位是相互定义的整体系统,认知即产生于这种"结构耦合"的互动领域(Maturana & Varela, 1970)。在他的理论中,认知被理解为维持有机体与环境持续互动的功能组织,而非局限于神经组织的属性。
这种视角下,认知是生命系统的基本过程而非专属属性。当认知被视为生物体维持生存的适应性机制时,人类与植物的认知能力可置于系统发育连续谱中考察(Lyon, 2005)。神经系统只是扩展了互动范围,并非认知的必要条件——这为植物认知研究提供了理论合法性。
图3 马图拉纳认知生物学的"结构耦合"模型
核心概念"结构耦合":认知产生于生物体与环境的递归互动过程,而非预先存在于神经系统中。该模型揭示:认知是所有生命的本质属性;生物体通过互动建构自身的"环境界";神经系统仅为认知过程的一种实现方式(Maturana & Varela, 1970; Lyon, 2005)。
3 植物认知的实验证据:从行为到机制
传统观点认为植物行为是刻板的、基因编程的被动响应,与动物行为的复杂性形成鲜明对比。这种偏见导致植物认知研究长期滞后——即便观察到学习等行为,也被简单归因于生理反应而非认知过程。
近年实验证据已明确证实:植物是具备复杂认知能力的动态行为主体。通过对感知、学习、记忆等行为的系统观测,植物展现出远超传统认知的环境适应策略,以下为关键实验发现。
3.1 学习与记忆:植物的行为可塑性
近年研究已积累了植物认知的有力证据,揭示植物具备远超传统认知的行为复杂性。代表性研究包括含羞草的习惯化学习(Gagliano et al., 2014)、植物的亲缘识别(Dudley & File, 2007)及挥发性信号通讯(Karban & Shiojiri, 2009)等,为植物认知提供了实验支撑。
Gagliano团队对含羞草(Mimosa pudica)的经典实验显示,植物能通过学习形成稳定记忆:经反复无害刺激后,含羞草会停止叶片闭合反应,且这种习惯化记忆可维持数周(Gagliano et al., 2014)。Dudley等(2007)则发现,植物能通过根系信号区分亲缘与非亲缘个体,通过调控资源分配减少种内竞争,体现了主动的行为决策能力。
3.2 亲缘识别与社会行为:信号机制与生态策略
植物通过精准的信号机制实现亲缘识别:根系分泌的尿囊素等化学信号可调控种内互动——亲缘植株间减少根系竞争,提高群体产量(Yang et al., 2018);地上部分则通过挥发性有机物传递防御信号,如蚜虫侵害时,邻近植株接收信号后会提前合成防御化合物(Karban et al., 2013)。这种社会行为体现了植物在资源分配与风险应对中的适应性认知策略。
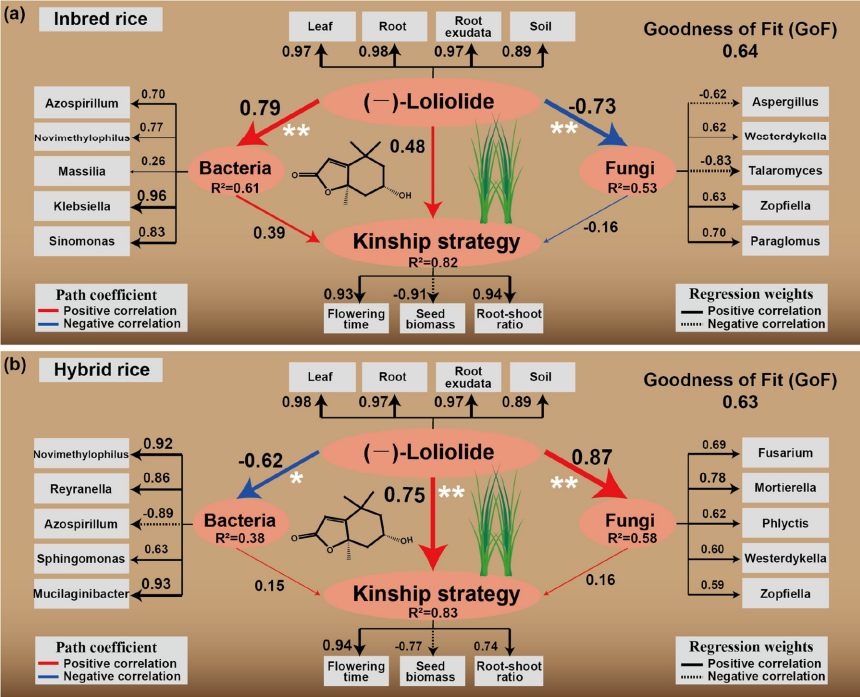
图4 不同亲缘关系水稻品种对混种土壤微生物调控的影响(a 自交水稻,b 杂交水稻)
PLS-PM 路径模型图,分别展示 (–)- 洛利内酯调控土壤微生物对常规稻(a)和杂交稻(b)亲缘策略的作用机制;箭头宽度代表路径系数大小,蓝色 / 红色为正 / 负效应,标注了关键细菌 / 真菌菌属、路径系数、决定系数(R²)和拟合优度(GoF);结果显示常规稻中该物质对细菌调控作用更强,杂交稻中对真菌调控作用更强,且杂交稻中该物质对亲缘策略的直接效应更高。Chen, X., Xu, Y., Ding, G.-C., Meiners, S. J., & Kong, C.-H. (2026). Chemically and microbially mediated kinship strategies in rice cultivar mixtures. Plant, Cell & Environment, 49(1), 51–63。
3.3 空间定向与多模态环境感知
空间定位是植物生存的核心能力。尽管缺乏动物的运动器官,植物通过整合多种感知信号,形成对环境的"位置地图",实现资源获取与风险规避的精准决策。
植物的空间定向依赖双系统整合:自我中心定向(通过本体感受监测自身姿态,Bastien et al., 2013)与异我中心定向(利用外部信号:光梯度引导向光性、土壤机械阻力调控根系生长,Semchenko et al., 2012)。此外,植物还能感知生物信号,如通过真菌网络传递的化学信息(Simard et al., 1997)、昆虫振动(Appel & Cocroft, 2014)及邻近植株的视觉信号(Gianoli & Carrasco-Urra, 2014),构建多维环境认知。
3.4 多模态信号感知:声音、振动与环境互动
植物具备复杂的声学感知能力:既能检测环境中的声音振动(如昆虫啃食产生的机械波,Appel & Cocroft, 2014),也能主动发出咔哒声进行种内通讯(Gagliano et al., 2012)。研究推测,某些植物可能通过类似回声定位的机制,利用声波反射感知周围环境结构(Gagliano, unpublished)。在空间导航中,植物通过自我中心定向(本体感受调节生长姿态)与异我中心定向(整合光、化学信号等环境线索)的协同,构建动态环境认知。这些发现表明植物的核心认知能力与动物存在趋同演化,但当前对其分子机制与神经替代系统的研究仍需深化,这是未来植物认知领域的关键突破方向。
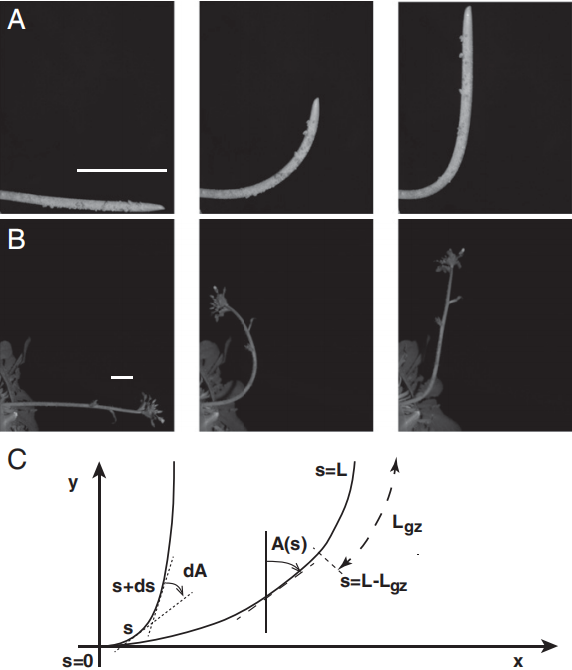
图5 植物向重力性反应的动态形态特征
A、B为向重力性反应的延时摄影:小麦胚芽鞘(A)与拟南芥花序(B)水平放置后向上弯曲的动态过程(标尺=1cm);C为弯曲形态的几何建模,通过倾斜角(A(s))和曲率(C(s))参数量化器官生长方向的精准调控,揭示植物通过自我感知实现空间定向的机制(Bastien et al., 2013)。
4 整合框架:环境可供性理论与植物认知研究
基于上述实验证据,植物作为认知主体的地位已得到充分支持。当前核心科学问题应聚焦于:植物如何通过独特的生理机制实现认知功能?其认知过程与动物存在哪些趋同与分化?
马图拉纳的认知生物学理论为解答这些问题提供了关键视角:认知是有机体与环境动态互动的涌现过程(Maturana, 1970)。这一观点推动我们超越结构决定论,从系统互动层面理解植物认知。
核心在于将感知视为探索环境"可供性"的主动过程——生物体通过持续评估环境提供的生存机会(如资源获取、风险规避),动态调整行为策略。这种感知-行动耦合机制,使植物能在缺乏神经系统的情况下实现适应性决策。
可供性理论(affordance theory)将环境特征定义为有机体可感知的互动机会(Chemero, 2008),为跨物种认知研究提供了统一框架。该理论已成功解释动物的环境适应:如鸟类通过评估树枝直径(功能可供性)选择筑巢位置(von Bayern et al., 2009),猴子利用工具的物理属性解决取食问题(Nelson et al., 2011)。
蝗虫的研究为此提供了重要类比:它们通过感知自身体型与障碍物的相对大小(可供性评估),决定是否穿越障碍并协调运动(Ben-Nun et al., 2013)。这提示植物可能通过类似机制——整合自身生理状态与环境信号,实现资源分配与空间导航的精准调控。
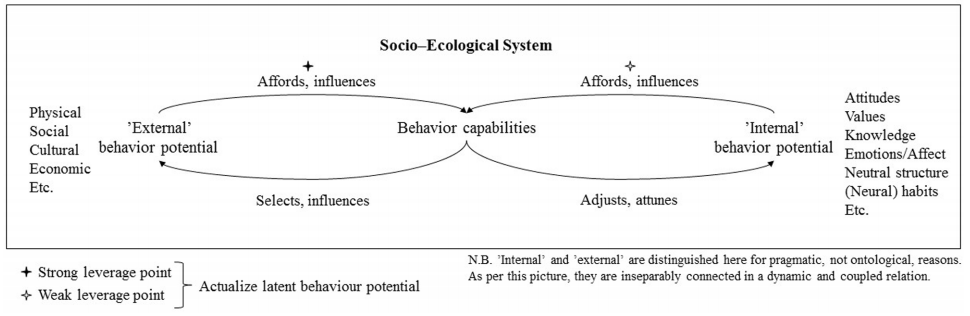
图6 有机体-环境耦合的动态认知系统模型
动态认知系统模型展示:有机体与环境通过持续互动塑造认知行为。以攀援植物为例,支撑物的直径、质地等物理特征(环境可供性)与卷须卷曲能力(生物体属性)的匹配,指导植物的攀爬决策。该模型摒弃传统主客二分,强调认知是系统互动的涌现结果(Kaaronen, 2017; Gibson, 1979)。
4.1 案例1:三维空间定向的声学机制
植物的声学感知能力为三维空间导航提供了新解释:研究发现植物能主动发出咔哒声并检测回声(Gagliano, unpublished),这一机制可能类似蝙蝠的回声定位——通过声波反射判断环境结构。对攀援植物而言,回声定位可能是其寻找宿主、规避障碍的关键手段,通过分析不同支架材料的回声差异,植物可评估支撑物的可用性(Bradbury & Vehrencamp, 2011)。
支架材料的声学特性(如吸收系数、反射强度)决定了回声的质量,进而影响植物的可供性评估。例如,木质支架与金属支架的回声差异可能引导植物选择更稳定的支撑结构,这种环境信号与行为决策的耦合,体现了植物认知的适应性逻辑。
4.2 案例2:邻体识别的生物声学信号
不同植物物种的特异性声学发射(Gagliano, unpublished)可能成为邻体识别的"声学指纹",类似动物通过鸣叫区分个体(Voigt-Heucke et al., 2010)。这与植物对光信号的感知机制(如通过反射光判断邻居身份,Aphalo et al., 1999)共同构成多模态识别系统,为亲缘选择与种间竞争提供决策依据。
植物生物声学研究尚处于探索阶段,虽缺乏直接实验证据,但其潜在价值显著:未来可通过精准声学测量与基因编辑技术,揭示声波信号的产生机制与感知通路,这将为理解植物认知的分子基础开辟新方向。
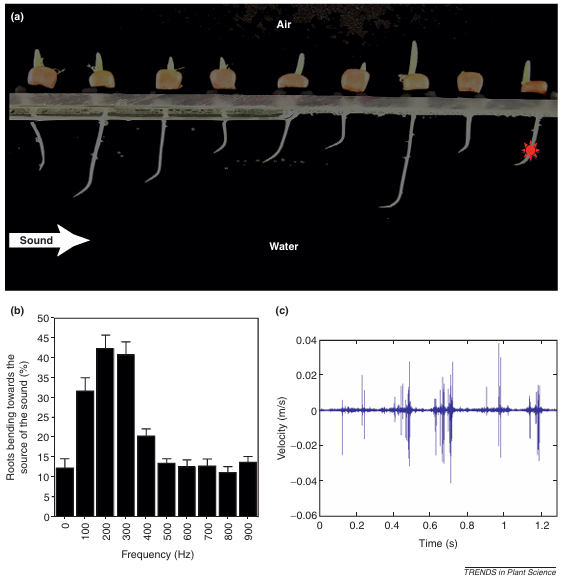
图7 玉米根系的生物声学响应机制
玉米根系的生物声学特性:(a)趋声性——根尖向220Hz声源弯曲;(b)频率选择性——对200-300Hz振动响应率最高;(c)主动声发射——根尖伸长区产生结构化声波。这些发现直接证实植物通过声学信号实现环境感知与通讯(Gagliano et al., 2012)。
5 结论与未来展望
近20年研究彻底颠覆了亚里士多德以来"植物缺乏行为能力"的传统认知,通过行为生态学与认知生物学的交叉视角,证实植物具备学习记忆、亲缘识别、多模态感知等复杂认知能力(Silvertown & Gordon, 1989; Silvertown, 1998)。这一突破推动行为概念从动物中心主义向生命系统普适性框架重构。
传统行为定义将植物响应简单归因为环境诱导的表型可塑性(Karban, 2008; Trewavas, 2009),虽勉强将植物纳入研究范畴,却忽视了行为的两大核心要素——主动行动与认知能动性,延续了"植物仅能做出刻板反应"的机械论认知。
动物行为研究早已整合运动能力与认知决策,但植物研究长期受限于观测技术。高速摄像技术的突破(如捕捉根尖生长动态、卷须搜索行为)直接证实了植物的主动行为能力,揭示其通过细胞信号网络实现类似动物的环境响应灵活性(Vincent et al., 2011)。
机械论还原论是植物认知研究的核心障碍。本文基于马图拉纳的认知生物学(结构耦合理论)与吉布森的生态心理学(可供性理论),提出开放认知框架:认知是生命系统与环境动态互动的涌现属性,而非神经组织的专属功能。
这一框架的核心论点可概括为:(1)环境通过"可供性"提供行为机会,而非直接决定行为(如光照梯度为植物提供生长方向选择,而非强制向光生长);(2)感知是主动的信息获取行为,通过持续监测环境信号(如化学梯度、声波振动)构建动态认知地图;(3)所有生命体均为具备自主性的行为主体,植物通过细胞网络实现分布式认知决策(Withagen et al., 2012)。
本文系统梳理了植物认知的实验证据(学习记忆、亲缘识别、多模态感知等),呼吁学界突破"神经中心主义"范式,以行为生态学与认知生物学的交叉视角:1)探索植物信号网络的认知机制;2)建立跨物种认知比较框架;3)挖掘植物智能对人工智能与生态管理的启示,推动认知科学向生命系统普适性理论发展。
延伸阅读
Gagliano, M. (2014). Learning by association in plants. Scientific Reports, 4, 6863.
Gagliano, M. (2018). Penser comme une plante: perspectives sur l'écologie comportementale et la nature cognitive des plantes. Cahiers Philosophiques, 153, 42-54.
https://blog.sciencenet.cn/blog-38998-1525356.html
上一篇:“拉夫领”:解析文艺复兴科学家“领饰”之密
下一篇:植物塑造国家象征:解码欧洲国家的植物文化基因
