ВЉЮФ
Grassland Research | ЧрВиИпджВЮяШКТфЮЂЩњОГгыЮШЖЈЦкИпКЎВнЕщЕФаЭЌаЇгІ
||
![]() ВнЕибаОПGR -- Grassland ResearchВнвЕПЦбЇСьгђЙњМЪбЇЪѕЦкПЏ
ВнЕибаОПGR -- Grassland ResearchВнвЕПЦбЇСьгђЙњМЪбЇЪѕЦкПЏ


ЮФеТаХЯЂ
ЦкПЏУћГЦЃКGrassland ResearchЃЈВнЕибаОПЃЉ
жаЮФБъЬтЃКЧрВиИпджВЮяШКТфЮЂЩњОГгыЮШЖЈЦкИпКЎВнЕщЕФаЭЌаЇгІ
ЕквЛзїепЃКСж РіЃЈжаЙњПЦбЇдКЮїББИпдЩњЮябаОПЫљЃЉ
ЭЈбЖзїепЃКаьаЫСМЃЈжаЙњПЦбЇдКЕиРэПЦбЇгыздШЛзЪдДбаОПЫљЃЉ
ЗвыепЃКСж Рі жаЙњПЦбЇдКЮїББИпдЩњЮябаОПЫљаЃ ЖдЃКЭѕЯўЗМ РМжнДѓбЇВнЕиХЉвЕПЦММбЇдК дкЖСВЉЪПЩњ
ЩљУїЃКИУЮФНіДњБэБрвыепЖдТлЮФЕФРэНтЃЌШчашВЮПМКЭв§гУЯрЙиФкШнЃЌЧыВщдФдЮФЁЃ
еЊвЊ
баОПБГОАЃКНќМИЪЎФъРДЃЌЙ§ЖШЗХФСКЭЦјКђБфЛЏЕФЫЋжигАЯьЪЙЕУИпКЎсдВнВнЕщЗЂЩњСЫВЛЭЌГЬЖШЕФЭЫЛЏЃЌЕМжТдкЯрЭЌЕиРэКЭЦјКђЕЅдЊЯТжВЮяШКТфЮЂАпПщЕФЗжвьМАЖрЮШЬЌЕФЙВДцЬиеїЁЃ
баОПЗНЗЈЃКВЩгУЪБПезЊЛЛЕФЗНЗЈЃЌЬНЬжЧрВиИпдИпКЎсдВнВнЕщЃЈalpine Kobresia meadowsЃЉВЛЭЌбнЬцЮШЬЌЮЂАпПщжВЮяШКТфЪ§СПКЭБШР§ЬиеїБфЛЏЃЌвдУїЮњИпКЎсдВнВнЕщЮЂАпПщМАШКТфЬиеїбнЛЏЕФёюКЯЙиЯЕЁЃ
баОПНсЙћЃКЃЈ1ЃЉИпКЎсдВнВнЕщВЛЭЌЭЫЛЏбнЬцЮШЬЌжаЃЌЫцзХЗХФСЧПЖШЕФдіИпЃЌЦфЕиЩЯзмЩњЮяСПГЪНЕЕЭЧїЪЦЃЌЦфжаЃЌЗХФСЧПЖШЁм3.65бђЕЅЮЛ/ЙЋЧъЕФВнГЁЦфЯФМОЕиЩЯзмЩњЮяСПзюИпЃЈ196.2 ЁР 20.3 g m-2ЃЉЃЌЧвЯджјИпгкЦфЫћЗХФСЧПЖШВнГЁЃЛЖјбаОПЗЖЮЇФкЦфЫћЗХФСВнГЁЯФМОЕиЩЯзмЩњЮяСПВювьВЛЯджјЁЃЃЈ2ЃЉИпКЎсдВнВнЕщКЬБОПЦКЭЩЏВнПЦжВЮяЙІФмШКживЊжЕЁЂЕиЩЯЩњЮяСПКЭЩњЬЌЮЛПэЖШЫцзХЗХФСЧПЖШЕФдіИпЖјЯджјНЕЕЭЁЃЃЈ3ЃЉЁн0ЁуCЛ§ЮТЁЂзмНЕЫЎСПЁЂКЃАЮЁЂОЖШКЭЮГЖШЖдИпКЎсдВнВнЕщВЛЭЌЮШЬЌЮЂАпПщЙІФмШКЬиеїЗжвьгАЯьЕФРлЛ§ЙБЯзТЪВЛзу20%ЁЃ
баОПНсТлЃКЃЈ1ЃЉЙ§ЖШЗХФСПЩвдНЕЕЭИпКЎсдВнВнЕщЕФГѕМЖЩњВњСІЃЌЕЋЩњЬЌЯЕЭГПЩвдЭЈЙ§ИФБфжВЮяШКТфзщГЩКЭНсЙЙЕжгљЗХФСИЩШХЃЌвдЮЌГжЯЕЭГЩњВњФмСІЕФЯрЖдЮШЖЈЁЃЃЈ2ЃЉЙ§ЖШЗХФСПЩвдЭЈЙ§ИФБфЮЂАпПщЕФБШР§дьГЩжВЮяШКТфЕФзДЬЌзЊБфЃЌВЂОнДЫНЈСЂСЫжВЮяШКТфЮЂАпПщЬиеїМАЯЕЭГЮШЖЈадЕФаЭЌБфЛЏЙиЯЕЁЃ
ЙиМќДЪЃКИпКЎЩњЬЌЯЕЭГЭЫЛЏЃЌЮЂАпПщЃЌЖрЮШЬЌЃЌЙ§ЖШЗХФСЃЌбнЬц
ЧАбд
ИпКЎВнЕщеМЧрВиИпдзмУцЛ§ЕФ35%вдЩЯЃЌзїЮЊИпдаѓФСвЕЩњВњЕФжїЬхЃЈеХН№ЯМЕШ, 2003ЃЉЃЌЦфжВБЛзДПіЕФгХСгЃЌВЛНіЖдЕБЕиаѓФСвЕЩњВњЃЌЭЌЪБЖдЛЦКгКЭГЄНСїгђЫЎЮФЩњЬЌЬиеївдМАЗчЁЂКЕЁЂРдЁЂЩГГОБЉЁЂЫЎЭССїЪЇЕШздШЛджКІЕФЗЂЩњМАЮЃКІГЬЖШОљОпгажиДѓгАЯьЃЈбюШъШй, 2002ЃЉЁЃНќФъРДЃЌЪмЕНЦјКђЁЂОМУЁЂеўВпЁЂЮФЛЏЕШЖрживђЫиЕФгАЯьЃЌИпКЎсдВнВнЕщжВБЛЗЂЩњВЛЭЌГЬЖШЕФбнЛЏЃЈУЯЗВЖАЕШ, 2014ЃЉЃЌгШЦфдкМвЭЅФСГЁзджїОгЊФЃЪНЯТЃЌФСГЁМфОАЙлЦЦЫщЖШдіМгЃЌГіЯжСЫЖрбнЬцЮШЬЌЙВДцЬиеїЃЈВмЙуУёЕШ, 2007ЃЉЁЃВнЕиЩњЬЌЯЕЭГЪЧгЩШєИЩгаађЕФЁЂОпгавЛЖЈНсЙЙЁЂЙІФмКЭЖЏЬЌБфЛЏЙцТЩЕФРыЩЂЕШМЖВуДЮзщГЩЃЈJørgensen et al., 2016; Wellemeyer et al., 2019ЃЉЃЌЮЂАпПщзїЮЊЯЕЭГаЁГпЖШПеМфНсЙЙЕЅдЊЃЌЦфЪєадЬиеїдкПеМфЩЯЯрЛЅИєРыЃЌгжБЫДЫЩјЭИЃЌЦфадзДЁЂНсЙЙКЭИёОжЬиеїЖдЦРЙРВнЕиЯжзДЁЂУїЮњВнЕибнЬцЧ§ЖЏвђзгМАбнЬцЙ§ГЬОпгаживЊвтвхЃЈЮкдЦФШЕШ, 2012ЃЉЁЃздШЛЩњЬЌЯЕЭГЮЂАпПщЕФаЮГЩвдЕиаЮЁЂЕиУВЬиеїЮЊЛљДЁЖдЫЎШШзЪдДНјаадйЗжХфЃЈGonzalez et al., 2010ЃЉЃЌвджВЮяШКТфЮЊКЫаФЖдЮЂАпПщЭСШРбјЗжЃЈШчЮоЛњЕЊЕШЃЉНјааРЙНиЁЂзЊвЦЁЂзЊЛЏКЭЛ§РлЃЈTongway, 1996; РюаЁОќЕШ, 2011; ЮкдЦФШЕШ, 2014; еХЗяНмЕШ, 2018ЃЉЃЌзюжеаЮГЩОпгаЯЪУїЬиеїЕФЭСШР-жВЮяИДКЯЬхЃЌЖјИУИДКЯЬхвВПЩвдЗДгГЯЕЭГЙЙМўЬиеїЃЈWedin & Tilman, 1996; KlinerovЈЂ & Dostal 2020; Lauren, 2018ЃЉЃЌвђДЫЃЌЮЂАпПщБЛШЯЮЊЪЧЩњЬЌЯЕЭГЖдЛЗОГМАИЩШХЯьгІЕФЁАжИЪОЦїЁБЁЃЗХФСЪЧШЫРрЖдИпКЎсдВнВнЕщзїгУЪБМфзюГЄЃЌзїгУаЇЙћзюЮЊЩюдЖЕФвЛжжИЩШХЗНЪНЃЌЦфзїгУЧПЖШЕФВЛЭЌЖджВБЛШКТфНсЙЙЁЂЭСШРРэЛЏаджЪЁЂЩњЬЌЯЕЭГЕФНсЙЙКЭЙІФмЁЂОАЙлИёОжЕШОљОпгаЩюдЖгАЯьЃЈProulx et al.,1998; бюРћУёЕШ, 2001ЃЉЁЃЗХФСЖдВнЕигАЯьЕФЛљБОзїгУЕЅдЊЮЊЮЂАпПщЃЈеХЮРЙњЕШ, 2003; ЪЏУїУїЕШ, 2015; КњбогюЕШ, 2018ЃЉЃЌЖјЗХФСЖдВЛЭЌЮЂАпПщзїгУЧПЖШВЛЭЌЃЌЕМжТСЫВнЕиЩњЬЌЯЕЭГЮЂАпПщжЎМфЕФЧЈвЦКЭЩјЭИЃЌЪЙЕУЗХФСЧПЖШЭЌЮЂАпПщИёОжбнЛЏжЎМфДцдкЖржиЪБМфГпЖШЩЯЕФжиЕўКЭЗжвьЃЈNicola et al., 2010; МжЬ№ЛЊ, 2019ЃЉЃЌНјЖјгАЯьСЫЯЕЭГЕФзмЬхЬиеїЁЃСЫНтЕиаЮЁЂЕиУВЁЂЦјКђЁЂИЩШХЖдИпКЎсдВнВнЕщЮЂАпПщМАЦфИёОжЕФгАЯьЃЌЖдУїЮњВнЕиЮШЖЈадЮЌГжМАЮШЬЌзЊЛЏЛњжЦЃЌЦЪЮіВнЕиздЮШЖЈадЮЌГжЙ§ГЬОпгаживЊвтвхЁЃБОбаОПвдЧрВиИпдЕфаЭИпКЎсдВнВнЕщЮЂАпПщжВЮяШКТфЬиеїМАЕиБэЬиеїЮЊбаОПЛљДЁЃЌЬНЬжЃЈ1ЃЉЯрЭЌЕиРэКЭЦјКђЕЅдЊЯТЃЌЗХФСЧПЖШВЛЭЌЪЧЗёПЩвддьГЩЖрЮШЬЌЕФЙВДцЃЛЃЈ2ЃЉИпКЎсдВнВнЕщЮЂАпПщЬиеїЭЌВнЕибнЛЏжЎМфЕФёюКЯЙиЯЕЃЌвдЦкЮЊИпКЎсдВнВнЕщПЩГжајЗЂеЙбаОПЬсЙЉРэТлЛљДЁЁЃ
ВФСЯКЭЗНЗЈ
бљЕиЕФбЁдё
ИљОнЁАЦјКђ-ЭСШР-жВЮязлКЯЫГађЗжРрЗЈЁБЃЈthe Comprehensive and Sequential Classification System of GrasslandЃЉЃЌИпКЎсдВнВнЕщжВБЛРраЭЙщЪєгкЁАКЎРфГБЪЊЖргъЖГдЁЂИпЩНВнЕщРрЁБЃЌОнДЫЃЌбЁдёЧрКЃЪЁЦјКђЬиеїТњзуЁн0ЁуCЛ§ЮТДІгк659.1-1228.4ЁуCЃЌЩњГЄМОНЕЫЎСПДІгк257.5-693.7mmЃЌЪЊШѓЖШЃЈНЕгъСП/0.1ЁСЛ§ЮТЃЉДІгк2.4-5.3ЕФКЃББжнЁЂгёЪїжнЁЂЙћТхжнЁЂКЃЮїжнЁЂКЃФЯжнЕФЯргІЧјгђЮЊВЩбљЧјгђЃЌВЂИљОнИпКЎсдВнВнЕщжВЮяШКТфКЭЕиБэЬиеїЕФЪЕЕиПБВьЪ§ОнЃЌзюжеШЗЖЈ109ИіВЩбљЕуЃЈЭМ1ЃЉЁЃ
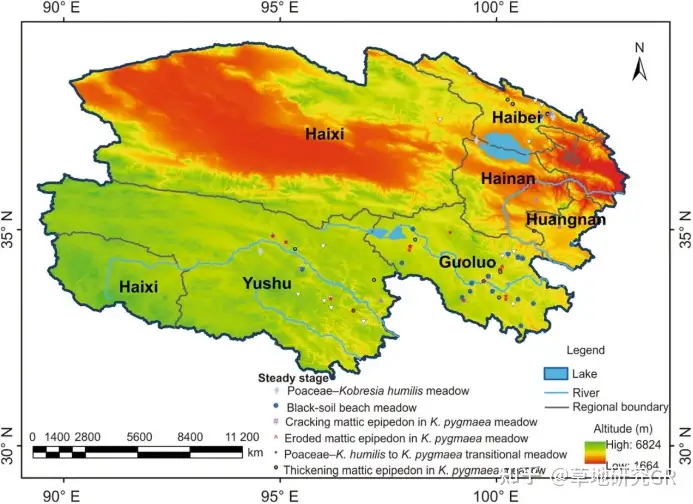
ЭМ1 ВЩбљЕуМАЦфЬцДњЮШЖЈзДЬЌ
бнЬцЮШЬЌЕФШЗЖЈ
ИљОнИпКЎсдВнВнЕщБЛЖЏ-жїЖЏЭЫЛЏМйЫЕЃЈthe passiveЉ\active degradation process hypothesisЃЉМАИпКЎсдВнВнЕщжВЮяШКТфКЭЕиБэЬиеїЃЌНЋЦфЭЫЛЏбнЬцЙ§ГЬЛЎЗжЮЊ8ИіЮШЬЌЃЈCao et al., 2007ЃЉЃЌЗжБ№ЮЊЃККЬБОПЦ-АЋЩњсдВнВнЕщЃЈPoaceae-Kobresia humilis meadowЃЉЃЌАЋЩњсдВнВнЕщЃЈK. humilis meadowЃЉЃЌКЬБОПЦ-АЋЩњсдВнКЭИпЩНсдВнВнЕщЕФЙ§ЖЩЮШЬЌЃЈPoaceae-K. humilis to K. pygmaea transitional meadowЃЉЃЌИпЩНсдВнВнЕщВнеББэВуе§ГЃЦкЃЈhealthy mattic epipedon in K. pygmaea meadowЃЉЃЌИпЩНсдВнВнЕщВнеББэВуМгКёЦкЃЈthickening mattic epipedon in K. pygmaea meadowЃЉЃЌИпЩНсдВнВнЕщВнеББэВуПЊСбЦкЃЈcracking mattic epipedon in K. pygmaea meadowЃЉЃЌИпЩНсдВнВнЕщВнеББэВуАўЪДЦкЃЈeroded mattic epipedon in K. pygmaea meadowЃЉКЭКкЭСЬВ-дгРрВнДЮЩњТуЕиЃЈblackЉ\soil beach meadowЃЉЃЈЭМ 1ЃЉЁЃгЩгкФГаЉЮШЬЌжВЮяШКТфЪ§СПЬиеїПЩвддкЗХФСЧПЖШИФБфЕФЬѕМўЯТПьЫйбнЬцЃЌвђЖјБОбаОПНЋИпКЎсдВнВнЕщЛЎЗжЮЊИќЮЊЮШЖЈЕФ5РрЮШЬЌЃКЕквЛРрЃЈIЃЉЃЌжївЊАќРЈКЬБОПЦ-АЋЩњсдВнВнЕщЃЈPoaceae-Kobresia humilis meadowЃЉЃЌАЋЩњсдВнВнЕщЃЈK. humilis meadowЃЉвдМАКЬБОПЦ-АЋЩњсдВнКЭИпЩНсдВнВнЕщЕФЙ§ЖЩЮШЬЌЃЈPoaceae-K. humilis to K. pygmaea transitional meadowЃЉЃЛЕкЖўРрЃЈIIЃЉЃЌжївЊАќРЈИпЩНсдВнВнЕщВнеББэВуе§ГЃЦкЃЈhealthy mattic epipedon in K. pygmaea meadowЃЉКЭИпЩНсдВнВнЕщВнеББэВудіКёЦкЃЈthickening mattic epipedon in K. pygmaea meadowЃЉЃЛЕкШ§РрЃЈIIIЃЉЃЌжївЊАќРЈИпЩНсдВнВнЕщВнеББэВуПЊСбЦкЃЈcracking mattic epipedon in K. pygmaea meadowЃЉЃЛЕкЫФРрЃЈIVЃЉЃЌжївЊАќРЈИпЩНсдВнВнЕщВнеББэВуАўЪДЦкЃЈeroded mattic epipedon in K. pygmaea meadowЃЉЃЛЕкЮхРрЃЈVЃЉЃЌжївЊАќРЈКкЭСЬВ-дгРрВнДЮЩњТуЕиЃЈblack-soil beach meadowЃЉЁЃбљЦЗВтЖЈМАМЦЫуЗНЗЈ
ВЩбљЪБМфЮЊ2011-2012Фъ8дТжВЮяЩњГЄЪЂЦкЁЃжВЮяШКТфЪ§СПЬиеїЕїВщВЩгУбљЯпЗНЗЈЃЌдкУПИібљЕибизХ100 mбљЯпНјааВЩбљЃЌУПИібљДјбЁШЁ8ИібљЗНЃЌбљЗНУцЛ§ЮЊ50 cm ЁС 50 cmЁЃЗжБ№ЪеМЏУПИібљЗНУПжжЙІФмШКЃЈЙІФмШКжївЊАќРЈКЬБОПЦЃЈPoaceaeЃЉЁЂЩЏВнПЦЃЈCyperaceaeЃЉЁЂЖЙПЦЃЈFabaceaeЃЉЁЂдгРрВнЃЈforbsЃЉКЭЙрДдЃЈshrubsЃЉЃЉЕиЩЯЩњЮяСПЃЌ60-80ЁуCКцИЩ48hЃЌГЦжиЃЛМЧТМУПжжЙІФмШКИЧЖШЁЃЭЌЪБМЧТМУПИібљЗНзмИЧЖШЁЂЦТЖШЁЂЦТЯђЁЂЦТЮЛЃЌвдМАУПИібљЕиОЮГЖШЃЌВЩгУЗЧНсЙЙЪНЮЪОэЕїВщШЗЖЈУПИібљЕиЕФЗХФСЧПЖШЃЈСжРіЃЌ2017ЃЉЃЌвдШЗЖЈМвЭЅФСГЁЕФУцЛ§ЁЂЗХФСЩќаѓЕФжжРрКЭЪ§СПвдМАВнГЁПЊЪМКЭНсЪјЗХФСЕФШеЦкЁЃАДеевЛЭЗъѓХЃЕФЗХФСЧПЖШЕШгк4жЛУрбђЕФЗХФСЧПЖШМЦЫуЃЈXuЃЌ1994ЃЉЃЌНЋбаОПбљЕиЗХФСЧПЖШЩшжУЮЊ4ИіЬнЖШЃЌЗжБ№ЮЊЃК<3.65ЁЂ3.65-7.50ЁЂ8.25-13.00КЭ>13.00бђЕЅЮЛha-1ЁЃЕБИпКЎсдВнВнЕщЭЫЛЏЮЊКкЭСЬВ-дгРрВнДЮЩњТуЕиЪБЃЌЭЈГЃВнЕивбОЮоЗХФСРћгУМлжЕЃЌДЫЪББъМЧЕФЗХФСЧПЖШЪЧИУЮШЬЌзюКѓЗХФСЪБЕФЗХФСЧПЖШЁЃ
Ъ§ОнЕФЭГМЦЗжЮі
ЮЂАпПщРраЭАДеежВЮядкЮЂАпПщжаНЈШКжж/ЙІФмШК+бЧНЈШКжж/ЙІФмШКУќУћЃЌНЋЫљгаЮЂАпПщЛЎЗжГЩКЬБОПЦ-АЋЩњсдВнЮЂАпПщЁЂАЋЩњсдВн-ИпЩНсдВнЮЂАпПщЁЂИпЩНсдВнЮЂАпПщЁЂИпЩНсдВн-дгРрВнЮЂАпПщМАдгРрВнЮЂАпПщЃЈВмЙуУёЕШ, 2007; СжРі, 2017ЃЉЁЃвдЮЂАпПщжВЮяЙІФмШКИЧЖШЁЂЕиЩЯЩњЮяСПКЭживЊжЕЕФЦНОљжЕБэеїИпКЎсдВнВнЕщИїЮШЬЌжВЮяЙІФмШКЬиеїЃЈLin, 2017ЃЉЁЃвдЮЂАпПщжВЮяЙІФмШКживЊжЕЮЊЛљДЁЪ§ОнЃЌгІгУShannon-WienerжИЪ§МЦЫуИпКЎсдВнВнЕщИїЮШЬЌжВЮяЙІФмШКЕФЩњЬЌЮЛПэЖШЬиеїЃЈLinЃЌ2017ЃЉЁЃВЩгУSPSSЃЈ19.0ЃЉЖджВЮяЙІФмШКЕиЩЯЩњЮяСПЁЂИЧЖШЁЂживЊжЕКЭЩњЬЌЮЛПэЖШНјааЕЅвђЫиЗНВюЗжЮіЃЌВЂМЦЫуУПжжЙІФмШКИїЪ§СПЬиеїдкИїЮШЬЌжаЕФОљжЕКЭБъзМЮѓЁЃЦфжаЕиЩЯЩњЮяСПЁЂИЧЖШЁЂживЊжЕКЭЩњЬЌЮЛПэЖШЕФЙЋЪНШчЯТЃК
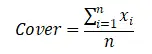
ЦфжаИЧЖШЪЧвЛИібљЕижавЛИіжВЮяЙІФмШКЕФИЧЖШЃЌxiЪЧвЛИібљЗНжавЛИіжВЮяЙІФмШКЕФИЧЖШЃЌnЪЧбљЕижабљЗНЕФЪ§СПЁЃ
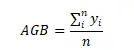
ЦфжаAGBЪЧвЛИібљЕижавЛИіжВЮяЙІФмШКЕФЕиЩЯЩњЮяСПЃЌyiЪЧвЛИібљЗНжавЛИіжВЮяЙІФмШКЕФЕиЩЯЩњЮяСПЃЌnЪЧбљЕижабљЗНЕФЪ§СПЁЃ
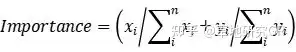
ЦфжаживЊжЕЪЧвЛИібљЕижавЛИіжВЮяЙІФмШКЕФживЊадЃЌxiЪЧвЛИібљЗНжавЛИіжВЮяЙІФмШКИЧЖШЃЌyiЪЧвЛИібљЗНжавЛИіжВЮяЙІФмШКЕФЕиЩЯЩњЮяСПЃЌnЪЧжВЮяЙІФмШКЕФЪ§СПЁЃ
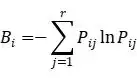
ЦфжаBiЪЧвЛИібљЕижажВЮяЙІФмШКЕФЩњЬЌЮЛПэЖШЃЌPijЪЧбљЗНжажВЮяЙІФмШКЕФживЊжЕЁЃУПвЛРрЮЂеОЕуОпгаФГжжЛЗОГЬиеїЕФИХТЪМЦЫуЮЊОпгаИУЛЗОГЬиеїЕФЮЂеОЕуЪ§СПгыЮЂеОЕузмЪ§ЕФБШжЕЁЃРћгУR3.6.1ШэМўНјааШпгрЗжЮіЃЈRDAЃЉЃЌМЦЫуЫљгаЮЂЮЛЕужВЮяЙІФмШКгыИїЛЗОГвђзгЕФёюКЯЙиЯЕЁЃЪЊШѓЖШЪ§ОнЕФШЗЖЈЃКвд1962-2012ФъШЋЧђШеЦјКђРњЪЗЪ§ОнЭјТчЃЈDaily Global Historical Climatology Network-DailyЃЌGHCN-DЃЉКЭжаЙњЦјЯѓОжЙњМвЦјЯѓаХЯЂжааФжЦзїЕФЁАжаЙњЕиУцЦјКђзЪСЯШежЕЪ§ОнМЏЁБЮЊЛљДЁЃЌбЁШЁШеНЕЫЎКЭШеОљЮТСНИівЊЫиЃЌЭЈЙ§жаЙњПЦбЇдКЕиРэПЦбЇгызЪдДбаОПЫљЩњЬЌЯЕЭГЭјТчЙлВтгыФЃФтжиЕуЪЕбщЪвПЊЗЂЕФЁЖЛљгкMatlabЕФЙњМЪжївЊЦјЯѓеОЕуЪ§ОнПеМфВхжЕдЄДІРэЯЕЭГЁЗЃЈШэМўЕЧМЧКХЃК2014SR142569ЃЌШэМўЗжРрКХЃК30200-7500ЃЉКЭАФДѓРћбЧЙњСЂДѓбЇПЊЗЂЕФЩЬвЕШэМўANUSPLINЦјЯѓЪ§ОнПеМфВхжЕЗНЗЈЃЌЩњГЩПеМфЭјИёЮЊ1kmЕФЦјЮТКЭНЕЫЎеЄИёЪ§ОнЁЃВЂвдДЫЮЊЛљДЁШЗЖЈВЛЭЌбљЕиЦјЮТКЭНЕЫЎСПЃЈЭѕОќАюЕШ, 2017ЃЉЁЃЮЂАпПщЙІФмШКЬиеїЭЌЦјКђвђзгжЎМфЕФЙиСЊадЗжЮіЃКВЩгУRЃЈVersion 4.0.5ЃЉШэМўЕФRDAШпгрЗжЮівдОЖШЁЂЮГЖШЁЂКЃАЮИпЖШЁЂЁн0ЁуCЛ§ЮТКЭФъНЕЫЎСПЮЊЛЗОГвђзгЃЌвдЮЂАпПщЙІФмШКЬиеїЮЊЯьгІБфСПЃЌЗжЮіЛЗОГвђзгЖдЯьгІБфСПБфЛЏЕФЙБЯзТЪЁЃ
НсЙћ
ЮЂАпПщжВЮяЙІФмШКЬиеї
ИпКЎсдВнВнЕщЕиЩЯЩњЮяСПЭЌЦфЫљДІбнЬцЮШЬЌЕФЙиЯЕБШЦфЫљДІЕиРэЮЛжУКЭЦјКђвђЫиЕФЙиЯЕИќЮЊУмЧаЁЃбаОПЗЂЯжЃЌЕБЗХФСЧПЖШДѓгк3.65бђЕЅЮЛ/ha-1ЪБЃЌИпКЎсдВнВнЕщжВЮяЩњГЄЦфЕиЩЯЩњЮяСПИЩжиЮЊ95.2ЁР7.8 ~ 100.6ЁР9.5 g m-2ЃЌЧвИїЮШЬЌжЎМфЕФГѕМЖЩњВњСІМфЮоЯджјВювьЃЈp > 0.05ЃЉЁЃЖјЕБЗХФСЧПЖШЕЭгк3.65бђЕЅЮЛ/ha-1ЪБЃЌЦфжВЮяЩњГЄЪЂЦкЕиЩЯЩњЮяСПИЩжиЮЊ196.3ЁР20.3 g m-2ЃЌЯджјИпгкЗХФСЧПЖШДѓгк3.65бђЕЅЮЛ/ha-1ЕФЫљгаЮШЬЌЁЃвђДЫЃЌИпКЎсдВнВнЕщЕиЩЯЩњЮяСПИЩжиЫцЗХФСЧПЖШЕФБфЛЏЖјЗЂЩњБфЛЏЁЃЦфжаЃЌЫцзХЗХФСЧПЖШЕФдіМгЃЌКЬБОПЦЁЂЩЏВнПЦКЭЖЙПЦЙІФмШКЕиЩЯЩњЮяСПЯджјНЕЕЭЃЌЕЋЦфЫћЙІФмШКЕиЩЯЩњЮяСПИЩжиНЯЧА3епЕФЙІФмШКЮШЖЈЃЌНјЖјЕМжТдкДѓЗЖЮЇЗХФСЧПЖШКЭДѓПеМфГпЖШЯТЃЌИпКЎВнЕщЕФзмЩњВњСІЕФЮШЖЈЃЈЭМ2ЃЉЁЃ
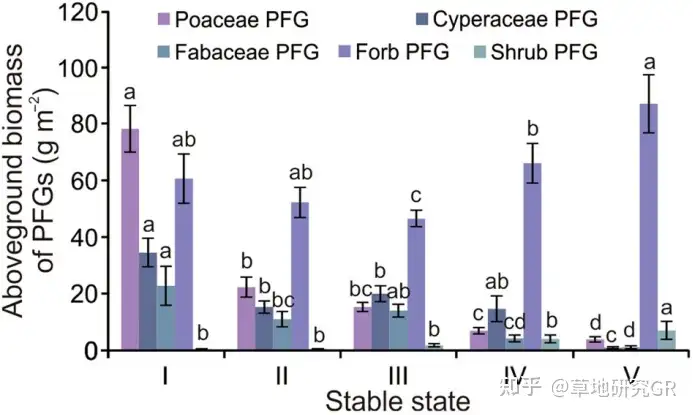
ЭМ2 ВЛЭЌбнЬцЮШЬЌЯТжВЮяЙІФмШКЃЈPFGsЃЉЕиЩЯЩњЮяСПЬиеїЁЃIЁЂIIЁЂIIIЁЂIVКЭVЗжБ№ДњБэPoaceae-Kobresia meadow and Poaceae-K. humilis to K. pygmaea transitional meadow; healthy and thickening mattic epipedon in K. pygmaea meadow; cracking mattic epipedon in K. pygmaea meadow; and eroded ma
ИпКЎсдВнВнЕщЫцзХЗХФСЧПЖШЕФдіМгКЬБОПЦЁЂЩЏВнПЦКЭЖЙПЦЕФживЊГЪНЕЕЭЧїЪЦЃЌдгВнГЪдіИпЧїЪЦЃЌЖјЙрФОБЃГжЯрЖдЮШЖЈЁЃгЩгкживЊжЕПЩвдЗДгГЦфдкжВЮяШКТфжаЕФзЪдДЛёШЁФмСІЃЌЪЧжВЮяШКТфЪ§СПЬиеїЕФвЛИізлКЯаджИБъЃЌвђДЫРћгУЮЂАпПщЙІФмШКживЊжЕБфЛЏЧїЪЦЬиеїНЋИпКЎсдВнВнЕщЛЎЗжЮЊ3ИібнЬцНзЖЮЃКЕквЛНзЖЮжївЊАќРЈКЬБОПЦ-сдВнВнЕщЃЈPoaceae-Kobresia meadowЃЉЃЌАЋЩњсдВнВнЕщЃЈK. humilis meadowЃЉКЭКЬБОПЦ-АЋЩњсдВнКЭИпЩНсдВнВнЕщЕФЙ§ЖЩЮШЬЌЃЈPoaceae-K. humilis to K. pygmaea transitional meadowЃЉЃЉЃЌетвЛРрЮШЬЌДІгкЖЬЦкНћФСЛђЧсЖШЗХФСЬѕМўЯТЃЌжВЮяШКТфЕщвдКЬБОПЦжВЮяЮЊгХЪЦжжЃЌЮЊЗХФСЭЫЛЏбнЬцЕФЦ№ЪМНзЖЮЁЃЕкЖўИіНзЖЮЃЈжївЊАќРЈИпКЎИпЩНсдВнВнЕщВнеББэВуе§ГЃЦкЃЈhealthy mattic epipedon in K. pygmaea meadowЃЉЃЌИпКЎИпЩНсдВнВнЕщВнеББэВуМгКёЦкЃЈthickening mattic epipedon in K. pygmaea meadowЃЉКЭИпЩНсдВнВнЕщВнеББэВуПЊСбЦкЃЈcracking mattic epipedon in K. pygmaea meadowЃЉЃЌетвЛРрЮШЬЌЕФжївЊЬиЕуЮЊвдЩЏВнПЦЮЊгХЪЦжжЁЃЕкШ§ИіНзЖЮжївЊАќРЈАќРЈИпЩНсдВнВнЕщАўЪДЦкЃЈeroded mattic epipedon in K. pygmaea meadowЃЉКЭКкЭСЬВ-дгРрВнДЮЩњТуЕиЃЈblack-soil beach meadowЃЉЃЌетвЛРрЮШЬЌЕФжївЊЬиЕуЮЊдгРрВнГЩЮЊжВЮяШКТфЕФгХЪЦРрШКЃЌЖјКЬБОПЦКЭЩЏВнПЦГЩЮЊАщЩњжжЁЃЖЙПЦКЭКЬБОПЦжВЮяЙІФмШКЖдЗХФСЧПЖШИФБфЕФЯьгІЯрЫЦЃЌвђДЫВЛдйзИЪіЃЈЭМ3ЃЉЁЃ
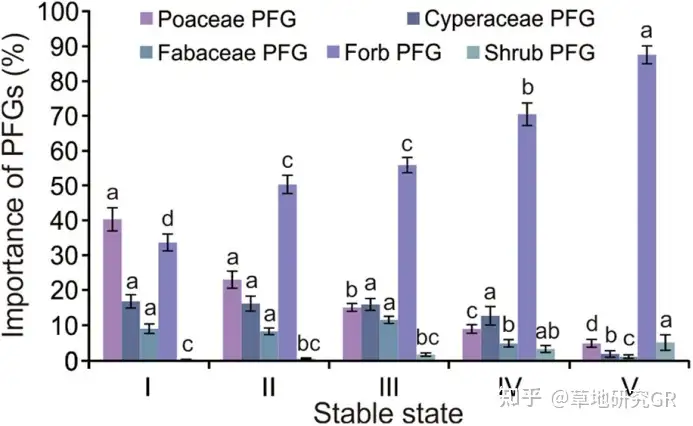
ЭМ3 ВЛЭЌбнЬцЮШЬЌЯТжВЮяЙІФмШКЃЈPFGsЃЉживЊжЕЬиеїЁЃIЁЂIIЁЂIIIЁЂIVКЭVЗжБ№ДњБэPoaceae-Kobresia meadow and Poaceae-K. humilis to K. pygmaea transitional meadow; healthy and thickening mattic epipedon in K. pygmaea meadow; cracking mattic epipedon in K. pygmaea meadow; and eroded mattic
ИпКЎсдВнВнЕщКЬБОПЦЁЂЩЏВнПЦЁЂЖЙПЦКЭдгВнЩњЬЌЮЛПэЖШОљЫцзХЗХФСЧПЖШЕФдіМгЖјМѕаЁЁЃИљОнИїЙІФмШКЩњЬЌЮЛПэЖШЫцЗХФСЧПЖШЕФБфЛЏЧїЪЦЃЌНЋетвЛЭЫЛЏбнЬцЙ§ГЬЛЎЗжГЩШ§ИіНзЖЮЁЃЕквЛНзЖЮжївЊАќРЈКЬБОПЦ-АЋЩњсдВнВнЕщЃЈPoaceae-Kobresia humilis meadowЃЉЃЌАЋЩњсдВнВнЕщЃЈK. humilis meadowЃЉЁЂКЬБОПЦ-АЋЩњсдВнКЭИпЩНсдВнВнЕщЕФЙ§ЖЩЮШЬЌЃЈPoaceae-K. humilis to K. pygmaea transitional meadowЃЉЁЂИпЩНсдВнВнЕщВнеББэВуе§ГЃЦкЃЈhealthy mattic epipedon in K. pygmaea meadowЃЉКЭИпЩНсдВнВнЕщВнеББэВудіКёЦкЃЈthickening mattic epipedon in K. pygmaea meadowЃЉЃЌЫќУЧЕФЙВЭЌЬиЕуЪЧКЬБОПЦЁЂЩЏВнПЦЁЂЖЙПЦКЭдгВнЕФЩњЬЌЮЛПэЖШОљзюИпЁЃЕкЖўНзЖЮжївЊАќРЈИпЩНсдВнВнЕщВнеББэВуПЊСбЦкЃЈcracking mattic epipedon in K. pygmaea meadowЃЉЁЂИпЩНсдВнВнЕщВнеББэВуАўЪДЦкЃЈeroded mattic epipedon in K. pygmaea meadowЃЉЃЌЫќУЧЕФЙВЭЌЬиЕуЪЧКЬБОПЦжВЮяЩњЬЌЮЛБЛЩЏВнПЦжВЮяЃЌЪЙЕУЩЏВнПЦжВЮяЬцДњКЬБОПЦжВЮяГЩЮЊЯЕЭГПЩЪГадФСВнЩњЬЌЮЛПэЖШзюДѓЕФжВЮяЙІФмШКЁЃЕкШ§ИіНзЖЮжївЊАќРЈКкЭСЬВ-дгРрВнДЮЩњТуЕиЃЈblack-soil beach meadowЃЉЃЌИУНзЖЮЕФЬиЕуЪЧКЬБОПЦЁЂЩЏВнПЦЁЂЖЙПЦКЭдгВнЕФЩњЬЌЮЛПэЖШЖМДѓЗљМѕаЁЃЌЖјЙрДдЕФЩњЬЌЮЛПэЖШдіМгЁЃШЛЖјЃЌетВЂВЛЫЕУїИпКЎсдВнВнЕщЯђИпКЎЙрДдВнЕщЗНЯђбнЬцЃЌЖјЪЧгЩгкЙрДдВнЕщвЛжББЛгУзїЯФМОФСГЁЃЌЕМжТЙрДдЯђИпКЎсдВнВнЕщЗНЯђбнЬцЃЌЫЕУїЯрБШЖЌМОВнГЁзїЮЊЯФМОФСГЁЕФЙрДдВнЕщЭЫЛЏИќЮЊбЯжиЃЈЭМ4ЃЉЁЃ
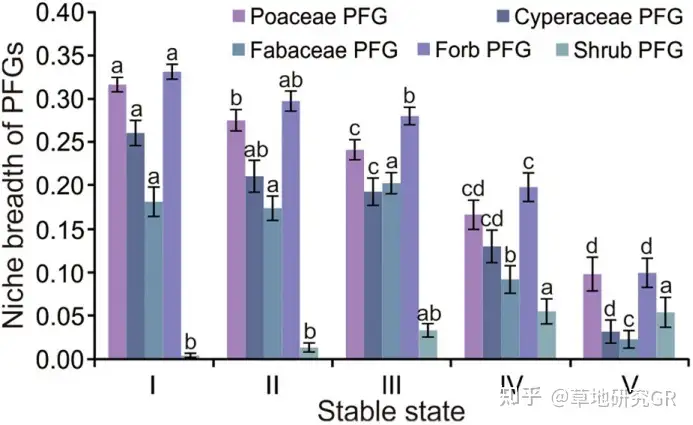
ЭМ4 ВЛЭЌбнЬцЮШЬЌЯТжВЮяЙІФмШКЃЈPFGsЃЉЩњЬЌЮЛПэЖШЬиеїЁЃIЁЂIIЁЂIIIЁЂIVКЭVЗжБ№ДњБэPoaceae-Kobresia meadow and Poaceae-K. humilis to K. pygmaea transitional meadow; healthy and thickening mattic epipedon in K. pygmaea meadow; cracking mattic epipedon in K. pygmaea meadow; and eroded ma
Л§ЮТЁЂНЕЫЎЁЂКЃБЈИпЖШКЭОЮГЖШЖдИпКЎВнЕщЮЂАпПщЗжВМгАЯьЕФЬиеї
вдКЃАЮИпЖШЁЂОЖШЁЂЮГЖШЁЂЁн0ЁуCЛ§ЮТКЭНЕЫЎСПЮЊЛЗОГБфСПЃЌвдКЬБОПЦЁЂЩЏВнПЦЁЂЖЙПЦЁЂдгРрВнКЭЙрДдЕФИЧЖШЁЂЕиЩЯЩњЮяСПКЭживЊжЕЮЊЯьгІБфСПЃЌЭЈЙ§RDAШпгрЗжЮіЗЂЯжЃКЩЯЪіЛЗОГБфСПЕФжїБфСПЖдЮЂАпПщжаКЬБОПЦЁЂЩЏВнПЦЁЂЖЙПЦЁЂдгРрВнКЭЙрДдЪ§СПЬиеїБфЛЏЕФРлЛ§НтЪЭФмСІЗжБ№ЮЊ13.29%ЁЂ8.07%ЁЂ9.05%ЁЂ6.44%КЭ2.44%ЁЃЦфжаЃЌОЖШКЭЮГЖШЖдКЬБОПЦЙІФмШКЪ§СПЬиеїБфЛЏЕФгАЯьДяЕНЯджјЫЎЦНЃЈP < 0.05ЃЉЃЛНЕЫЎСПКЭЮГЖШЖдЩЏВнПЦЙІФмШКЪ§СПЬиеїБфЛЏЕФгАЯьДяЕНЯджјЫЎЦНЃЈP < 0.05ЃЉЃЛНЕЫЎСПКЭОЖШЖдЖЙПЦЙІФмШКЪ§СПЬиеїЕФгАЯьДяЕНЯджјадЫЎЦНЃЈP < 0.05ЃЉЃЌОЖШЖддгРрВнЙІФмШКЪ§СПЬиеїБфЛЏЕФгАЯьДяЕНЯджјЫЎЦНЃЈP < 0.05ЃЉЃЛЕЋгЩгкЛЗОГБфСПЖджВЮяЙІФмШКЬиеїБфЛЏЕФНтЪЭФмСІЕЭЃЈЭМ5ЃЉЃЌвђДЫЃЌОЮГЖШЁЂКЃАЮИпЖШЁЂНЕЫЎСПКЭЁн0ЁуCЛ§ЮТЖМВЛЪЧЕМжТИпКЎсдВнВнЕщжВЮяШКТфЮЂАпПщЙІФмШКЪ§СПЬиеїЗжвьЕФжївЊдвђЁЃ
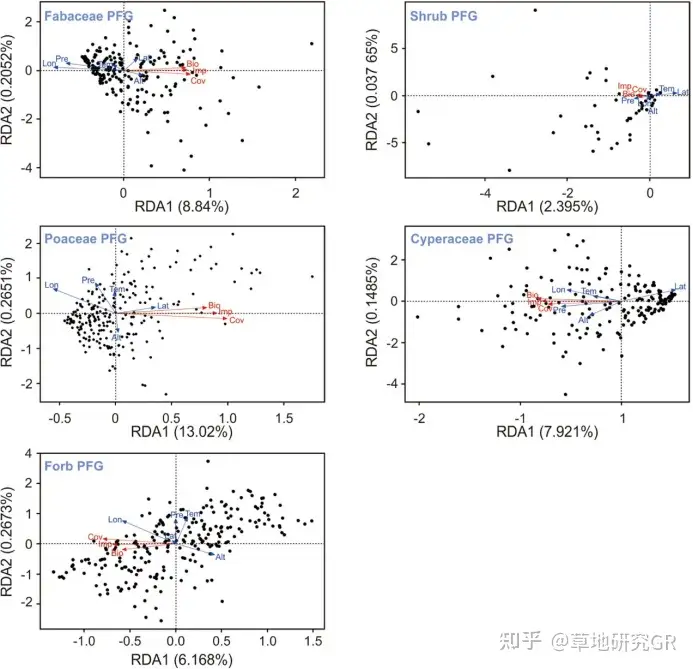
ЭМ5 ЮЂЦјКђвђзггыИпКЎсдВнВнЕщЮЂАпПщЙІФмШКЪ§СПЬиеїЙиСЊадЗжЮіЁЃAЁЂFКЭKЗжБ№ДњБэКЬБОПЦЕФЩњЮяСПЁЂИЧЖШКЭживЊжЕЃЛBЁЂGКЭLЗжБ№ДњБэЩЏВнПЦЕФЩњЮяСПЁЂИЧЖШКЭживЊжЕЃЛCЁЂHКЭMЗжБ№ДњБэЖЙПЦЕФЩњЮяСПЁЂИЧЖШКЭживЊжЕЃЛDЁЂIКЭNЗжБ№ДњБэдгВнЕФЩњЮяСПЁЂИЧЖШКЭживЊжЕЃЛEЁЂJКЭOЗжБ№ДњБэЙрДдЕФЩњЮяСПЁЂИЧЖШКЭживЊжЕЃЛP
ЮЂЕиаЮЖдИпКЎсдВнВнЕщЮЂАпПщЬиеїЕФгАЯь
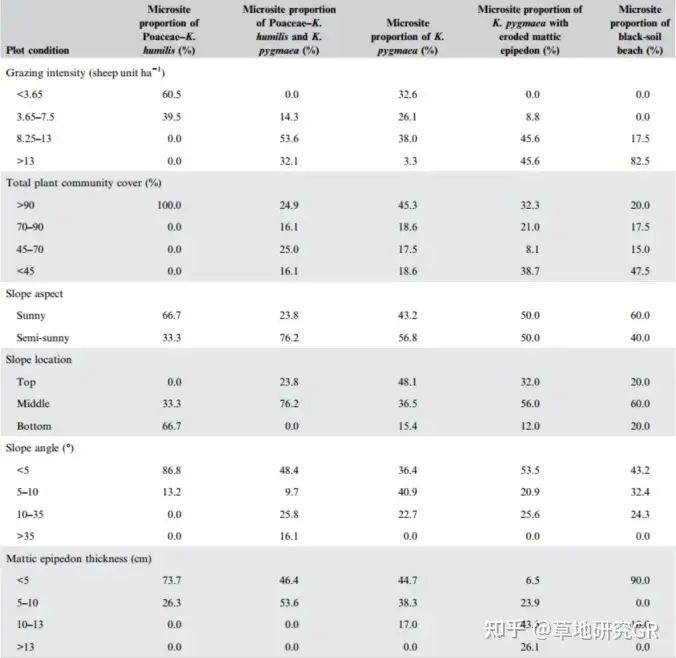
ЖдбаОПжаЫљгаЕїВщбљЗННјааЭГМЦЗжЮіЃЌЗЂЯжЃК11.1%ЪєгкКЬБОПЦ-АЋЩњсдВнЮЂАпПщЃЌ16.3%ЪєгкАЋЩњсдВн-ИпЩНсдВнЙ§ЖЩЮЂАпПщЃЌ36.2%ЪєгкИпЩНсдВнЮЂАпПщЃЌ16.6%ЪєгкИпЩНсдВн-дгРрВнЮЂАпПщЃЌ19.8%ЪєгкдгРрВнЮЂАпПщЁЃИпЩНсдВнЮЂАпПщГіЯжЕФБШР§зюИпЃЈБэ1ЃЉЁЃДѓЖрЪ§ЮЂАпПщЮЛгкЦТЖШаЁгк10ЁуЕФбєЦТЛђАыбєЦТЛђЩНМфЩНЙШжаЁЃЦфжаЭЌвЛФСГЁЮЂАпПщЕФБШР§КЭВњСПЛсЫцЗХФСЧПЖШБфЛЏЖјЗЂЩњИФБфЁЃР§ШчЃЌЭЌвЛФСГЁЮЛгкЦТжаЕФаЁсдВнЮЂАпПщЦфЩњВњСІОЭЛсЕЭгкЭЌвЛФСГЁЦфЫћЦТЮЛЮЂАпПщЕФЩњВњСІЃЌЦфжавЛВПЗждвђЪЧДІгкЦТЕзЕФФСГЁгЩгкЫљДІЧјгђЦјКђЬѕМўЯрЖдгХдНЖјЭЈГЃБЛФСУёгУгкФИаѓЗБжГЕиЛђгІЖдздШЛджКІЕФНєМББмФбЕиЃЌвђДЫБЛБЃЛЄЕФНЯКУЃЌЕМжТГЌЙ§90%ЕФКЬБОПЦ-АЋсдВнЮЂАпПщГіЯждкДЫжжЕиаЮЬѕМўЯТЃЈБэ1ЃЉЁЃДгИпКЎсдВнВнЕщзмИЧЖШКЭВнеББэВуЬиеїЭЌЮЂАпПщБШР§жЎМфЕФЙиЯЕЩЯЗжЮіЃЌЫцзХЗХФСЧПЖШЕФдіМгЃЌжВЮяШКТфзмИЧЖШГЪЯТНЕЧїЪЦЁЃИпЩНсдВнВнЕщВнеББэВуАўЪДЦкЃЈeroded mattic epipedon in K. pygmaea meadowЃЉзмжВЮяШКТфИЧЖШгыКкЭСЬВ-дгРрВнДЮЩњТуЕиЃЈblack-soil beach meadowЃЉВювьВЛЯджјЃЌЕЋЕЭгкЦфЫћЮШЬЌЁЃДЫЭтЃЌЮвУЧЕФбаОПНсЙћвВжЄЪЕСЫИпЩНсдВнВнЕщвЊЯыЮЌГжВнЕиЕФЯрЖдНЁПЕашвЊЮЌГжВнеББэВуДІгквЛИіЯрЖдКЯРэЕФКёЖШЃЈЭЈГЃЮЊ4-5РхУзЃЉЃЈБэ1ЃЉЁЃ
Бэ1 ВЛЭЌбљЕиЯТИпКЎВнЕщЮЂеОЕуБШР§
ЬжТл
ИпКЎВнЕщЖрЮШЬЌЙВДцЬиеїЪЧЖдЖёСгЛЗОГКЭЗХФСИЩШХЕФЯргІКЭЪЪгІ
ИпКЎВнЕщЫљДІЦјКђЛЗОГЖёСгЃЌЧвГЄЦкДІгкЗХФСКЭЦјКђБфЛЏЗДИДИЩШХЯТШдФмБЃГжЯрЖдЮШЖЈЕФЩњВњФмСІЫЕУїИУЯЕЭГОпгавЛЖЈЕФздЮвЮШЖЈадЮЌГжФмСІЁЃИпКЎсдВнВнЕщДѓдМдк2.4вкФъЧАбнЛЏЖјРДЃЈHerzschuh, 2006; Shen et al ., 2008ЃЉЁЃЧрВиИпдФПЧАЕФИпЩНВнЕщЭСЪЧдкШЋаТЪРЭэЦкЃЈ∼7-5 ka BPЃЉЗчЛ§КЭШЋаТЪРжаЦкЃЈ4 ka BPЃЉЫЎЛ§ЖјГЩЃЈZhang, 2019ЃЉЁЃЧрВиИпдаѓФСвЕЩњВњвбгаДѓдМ7300ФъЕФРњЪЗЃЈMiehe et al., 2009; Simons & Schön, 1998; Simons et al., 1994ЃЉЃЌДѓЙцФЃаѓФСвЕЩњВњвВгаДѓдМ3600ФъЕФРњЪЗЃЈQiu et al., 2015ЃЉЁЃвђДЫЃЌИпКЎсдВнВнЕщОпгагЦОУЕФаѓФСвЕЪЙгУРњЪЗЃЌдкДЫЦкМфЦјКђКЭШЫПкУмЖШЖМгаНЯДѓВЈЖЏЃЈР§ШчЃЌЧрВиИпддкЙЋдЊ6ЪРМЭдјГіЯжЙ§ДѓЙцФЃЕФШЫПкВЈЖЏЃЉЃЈLiao & Sun, 2003ЃЉЁЃЧЇАйФъРДЃЌИпКЎВнЕщЩњЬЌЯЕЭГвЛжБЪЧЧрВиИпдаѓФСвЕЩњВњЕФживЊЩњВњзЪСЯЖјБЛЙуЗКРћгУЃЌетЕМжТСЫИпКЎсдВнВнЕщОпгаСЫвЛЖЈЕФЪмИЩШХКѓЯЕЭГЙЙМўЕФЮШЖЈадЮЌГжФмСІКЭЛжИДФмСІЁЃЖрЮШЬЌЙВДцЬиеїПЩвдЪЙИпКЎсдВнВнЕщдкЖёСгЛЗОГжаЕжПЙИЩШХЃЌБЃГжЩњЬЌЯЕЭГЙЙМўЬиеїЕФЯрЖдЮШЖЈДѓСПбаОПБэУїЃЌИпКЎВнЕщЭЫЛЏбнЬцГЪЯжЮЊЗЧЯпадРыЩЂЮШЬЌБфЛЏЬиеїЃЈLiu et al., 1999ЃЉЁЃИФИяПЊЗХвдРДЃЌгаМвЭЅСЊВњГаАќд№ШЮжЦв§ЗЂЕФМвЭЅФСГЁЕФЧјБ№ЛЏОгЊв§Ц№СЫМвЭЅФСГЁЗХФСЧПЖШЕФБфЛЏЃЌИФБфжВЮяШКТфЕФЪ§СПЬиеїЃЈШчИЧЖШЁЂИпЖШЁЂвЖУцЛ§жИЪ§ЕШЃЉвдМАжВЮяШКТфЖдИЩШХЯьгІВпТдЃЈZhang, 1990ЃЉЁЃЧрВиИпдИпКЎсдВнВнЕщОјДѓЖрЪ§жВЮяЃЈШчK. pygmaeaЛђK. humilisЃЉЖМЪЧвдЮоадЗБжГЮЊжїЃЈZhou et al., 1986ЃЉЁЃетРржВЮяЕФЬиЕуЪЧЫцзХЗХФСЧПЖШвЛЖЈГЬЖШЕФдіМгЃЌЦфЗжоСЪ§ЁЂвЖЦЌКЭгзбПУмЖШГЪдіМгЧїЪЦЃЌетПЩФмЭЌИУРржВЮяЮоаджІЬѕЕФЭижВДгзюЭтВувЖЦЌПЊЪМЃЌЗжоСНкМфОрврЫцзХЗХФСЧПЖШЕФдіМгЖјЫѕЖЬЁЃМгжЎЃЌЫцзХЗХФСЧПЖШЕФдіИпЃЌАщЫцзХИљЭСБШКЭИљЯЕЩњЮяСПЕФбИЫйдіМгЃЌМгОчСЫЭСШРбјЗжЙЉгІгыжВЮяЩњГЄЖдбјЗжашЧѓжЎМфЕФУЌЖмЃЌЧветжжЙЉашУЌЖмдкЮоадЯЕЕФжааФЧјБэЯжгШЮЊЭЛГіЃЈCao et al., 2010ЃЉЃЌЕМжТСЫдкжиЗХФСЧПЖШЯТЃЌЮоадЯЕжааФЧјгђЗжоСжІЕФЫРЭіЃЌНјЖјдкИпКЎИпЩНсдВнВнЕщКЭИпКЎАЋЩњсдВнВнЕщГіЯжДѓСПЮожВБЛИВИЧЕФЮЂаЁГпЖШАпПщЕФдвђЃЈЭМ6ЃЉЁЃ

ЭМ6 ИпКЎИпЩНАЋсдВнВнЕщЃЈKobresia pygmaea MeadowЃЉЮоадЯЕЮожВБЛИВИЧЧјгђЃЈгУАзЩЋЭждВБэЪОЃЉ
ИпКЎсдВнВнЕщЫцзХЩќаѓЗХФСЧПЖШЕФдіМгЃЌЯЕЭГЕиЩЯЩњЮяСПгыЕиЯТЩњЮяСПжЎБШГЪНЕЕЭЧїЪЦЃЈCao et al., 2010ЃЉЃЌетвЛЗНУцгЩгкМваѓЕФВЩЪГКЭМљЬЄПЩвдДЬМЄИљЯЕЕФЩњГЄЃЌдіМгЭСШРНєЪЕЖШЃЈFan et al., 2020ЃЉЃЌМѕЩйЫЎЗжЩјЭИТЪЃЈDai et al., 2019ЃЉЃЌдіМгИљЯЕжївЊЗжВМВуДЮЫРИљЕФУмЖШЃЈLin, 2017ЃЉЃЌдіМгВнеББэВуКёЖШЃЈHou et al., 2004; Zhu, 2002ЃЉЃЌНјЖјИФБфжВЮяШКТфЕФЩњГЄКЭзщГЩНсЙЙЁЃР§ШчЃЌгЩгкКЬвЖаЭжВЮяЃЈАќРЈКЬБОПЦКЭЩЏВнПЦЃЉЩњЬЌЮЛПэЖШЕФЫѕаЁЃЌЕМжТГіЯжДѓСПЮожВБЛжВЮяИВИЧЕФТуТЖПеМфЃЌетаЉПеМфдкИпЧПЖШЗХФСЯТПЩФмБЛФЭФСЛђВЛПЩЪГжВЮяЫљеМОнЃЈLin, 2017ЃЉЃЌЩњЬЌЯЕЭГЕФетжжВпТдвЛЗНУцЪЙЕУЦфдкЩњГЄЦкБЃГжЯрЖдЮШЖЈЕФЩњВњСІЃЌЧвМѕЩйСЫЯЕЭГЖдаѓФСвЕЩњВњЕФЗўЮёФмСІЃЌПЭЙлЕМжТСЫЗХФСЧПЖШЕФНЕЕЭЛђМваѓЭЫГіЗХФСВнГЁЃЌНјЖјЪЙЕУЪмЫ№ЩњЬЌЯЕЭГЕФЕНанбјЩњЯЂЕФЛњЛсЃЌПЊЪМаТвЛТжЕФбнЬцЃЌЕЋетжжбнЬцЯрЖдЛКТ§ЃЌОпгагаЙибаОПЭЦЫуЃЌЮоЗХФСМлжЕЕФКкЭСЬВ-дгРрВнДЮЩњТуЕидкВЛЗХФСЬѕМўЯТашвЊ70-80ФъВХФмЛжИДЕНИпКЎИпЩНсдВнВнЕщЃЈChen, 2016ЃЉЁЃБОбаОПврЗЂЯжЃЌЕБЗХФСЧПЖШдіИпЃЌИпКЎсдВнВнЕщКЬБОПЦЙІФмШКБЛвжжЦЕФЬѕМўЯТЃЌЩњЬЌЯЕЭГдРДДІгкбЧгХЪЦжжЕФЙІФмШКЃЈШчЩЏВнПЦЃЉКЭЖОдгВнЕФЩњЬЌЮЛПэЖШЛсБЛЬсИпЃЌгУвдРћгУЩњЬЌЯЕЭГЪЃгрзЪдДЃЌНјЖјЮЌГжСЫЯЕЭГЩњВњФмСІЕФЯрЖдЮШЖЈЃЌЧветжжЯжЯѓдкжиЖШЗХФСЬѕМўЯТБэЯжЕФгШЮЊЭЛГіЁЃМгжЎЃЌМвЭЅФСГЁЕФЗжЛЇОгЊЃЌгЩгкФСГЁОгЊепВЩШЁЕФОгЊВпТдЕФВЛЭЌЃЌЕМжТСЫМвЭЅФСГЁМфжВЮяШКТфЬиеїЕФВювьЃЈLin et al., 2022ЃЉЃЌПЭЙлЩЯаЮГЩСЫЯрЭЌЕФЕиаЮКЭЦјКђЬѕМўЯТЖрЮШЬЌЙВДцЬиеїЃЈCao et al., 2007ЃЉЁЃвђДЫЃЌдкЙ§ЖШЗХФСЬѕМўЯТИпКЎсдВнВнЕщЯђПЩЪГадФСВнВњСПЕЭЕФЗНЯђбнЬцЪЧЩњЬЌЯЕЭГЕФЮЊЕжгљЙ§ЖШЗХФСИЩШХЕФвЛжжздЮвЕїНкЕФЙ§ГЬЁЃзмжЎЃЌИпКЎсдВнВнЕщдкЧрВиИпдЗжВМЙуЗКЧвФмЪЪгІвЛЖЈЗЖЮЇЕФЗХФСЧПЖШЃЌЭЌжВЮяШКТфЖдИЩШХЕФЪЪгІадбнЛЏЙиЯЕУмЧаЃЌЧветжжбнЛЏБфЛЏЛсв§Ц№ЕиБэЬиеїЁЂЭСШРЫЎЗжКЭбјЗжЕФаЭЌБфЛЏЁЃЖјЫцзХМвЭЅФСГЁМфвдЮЇРИЮЊЕЅЮЛЕФЮяРэЗжИюЃЌЕМжТСЫжВЮя-ЭСШРЯЕЭГРлЛ§БфЛЏЬиеїЕФЙЬЛЏЃЌЕМжТдкЯрЭЌЕиРэКЭЦјКђЕЅдЊЯТЖрЮШЬЌЙВДцЯжЯѓЕФаЮГЩЁЃ
ИпКЎВнЕщЕФЮЂАпПщЬиеїПЩФмЭЌЯЕЭГЮШЖЈЮЌГжУмЧаЯрЙи
ИпКЎсдВнВнЕщЮЂАпПщЯЕЭГдкПеМфЩЯЯрЛЅЖРСЂЃЌдкЙІФмБЫДЫЯрЙиЃЌдквЛЖЈИЩШХЬѕМўЯТЃЌЩњЬЌЯЕЭГПЩвдЭЈЙ§аЁГпЖШЮЂАпПщЕФздзщжЏЕїНкЃЌЪЕЯжЯЕЭГећЬхЬиеїЕФЮШЖЈЁЃИпКЎсдВнВнЕщжВЮяШКТфЭЈГЃГЪОлМЏадЗжВМЬиеїЃЈZhang, Huang, et al., 2003ЃЉЃЌВЂаЮГЩВЛЭЌИёОжЙцФЃКЭЧПЖШЕФАпПщЯтЧЖЬхЃЌИљОнжВЮя-ЭСШРЯЕЭГЕФЙиСЊадЬиеїЃЌУПИіЮЂАпПщЕФжВЮяШКТфЬиеїОљПЩвдЗДгГЦфЫљДІЕФЮЂЕиаЮКЭЮЂЦјКђЬиеїЁЃИљОнЕКгьЩњЮяЕиРэбЇРэТлЃЈisland biogeography theoryЃЉЃЌЮЂАпПщПЩвдБЛШЯЮЊЪЧЁАЕКгьЁБЃЌжмЮЇЕиЧјПЩвдБЛШЯЮЊЪЧЁАКЃбѓЁБЃЈMacArthur & Wilson, 1967ЃЉЁЃетжжАпПщЯтЬхЕФЮШЖЈадШЁОігкЮяжжжмзЊТЪЁЂАпПщУцЛ§КЭАпПщжЎМфЕФОрРыЃЈBueno & Peres, 2019ЃЉЁЃЫфШЛЗХФСИЩШХзїгУгкећИіИпКЎсдВнВнЕщЩњЬЌЯЕЭГЃЌЕЋЦфВЛЭЌЮЂАпПщЕФгАЯьЪЧВЛОљдШЕФЃЌетжжЗЧдШжЪИЩШХЬиеїМгОчСЫЩњЬЌЯЕЭГОАЙлЕФЦЦЫщЛЏЃЈJessica et al., 2017ЃЉЃЌЕМжТЃЌЫцзХЭЫЛЏбнЬцЕФНјааЃЌЮЂАпПщЕФЯрЛЅЩјЭИКЭзЊвЦЃЌНјЖјЪЕЯжЯЕЭГдквЛЖЈИЩШХЧПЖШЯТаЁГпЖШЕФБфЛЏвдЮЌГжДѓГпЖШЮШЖЈЃЛдкЙ§ЖШИЩШХЧПЖШЯТЗЂЩњзДЬЌзЊЛЏЃЈZhang, Huang, et al., 2003ЃЉЁЃВЛЭЌРраЭЕФЮЂАпПщдкИпКЎсдВнВнЕщВЛЭЌбнЬцЮШЬЌЙуЗКДцдкЁЃЫЎЗжЁЂЮТЖШЁЂЕиаЮКЭКЃАЮЖдИпКЎсдВнВнЕщЮЂАпПщжВЮяЙІФмШКЬиеїЗжвьЕФгАЯьНЯШѕЃЌЖјЗХФСЧПЖШЭЌЮЂАпПщжВЮяЙІФмШКЬиеїКЭБШР§жЎМфЗЂЩњСЫаЭЌБфЛЏЙиЯЕЁЃИпКЎсдВнВнЕщЗХФСЧПЖШЕФдіМгЃЌЦфжаПЩЪГФСВнБШР§ИпЕФЮЂАпПщГЪНЕЕЭЧїЪЦЃЌВЛПЩЪГФСВнБШР§ИпЕФЮЂАпПщГЪдіМгЧїЪЦЁЃетжжгаМваѓЗХФСааЮЊв§Ц№ЕФЮЂАпПщБШР§ЕФБфЛЏдкИФБфжВЮяШКТфЕФзщГЩКЭНсЙЙЕФЭЌЪБЮЌГжСЫжВЮяШКТфЕиЩЯзмЩњЮяСПЮШЖЈЃЌЕЋЕМжТЮШЬЌЕФзЊЛЏЁЃИпКЎсдВнВнЕщЪЧвЛИіПЊЗХЯЕЭГЃЌЯЕЭГЙЙМўЖдЭтВПИЩШХОпгаЗЧЯпадЕїНкФмСІЃЌетжжЯЕЭГЙЙМўвђзгЕФЗЧЯпадЕўМгаЇгІЃЈМДЩњЬЌЯЕЭГЖдИЩШХЕФе§ЗДРЁКЭИКЗДРЁаЇгІЃЉЃЌШнвзЕМжТЩњЬЌЯЕЭГЖдИЩШХЧПЖШБфЛЏЕФжЭКѓЯьгІЃЌНјЖјЕМжТЩњЬЌЯЕЭГзмЬхЬиеїГЪЯжНзЖЮадБфЛЏЬиеїЃЈLiu et al., 1999ЃЉЁЃЕЋИЩШХЧПЖШДяЕНвЛЖЈСйНчжЕЪБЃЌЦфЮЂАпПщЭЌЛљжЪжЎМфМДПЩЗЂЩњЯрЛЅЩјЭИКЭзЊЛЏЃЌЕМжТжВЮяШКТфЮШЬЌЕФзЊЛЏЃЈOborny & Hubai, 2014ЃЉЁЃвђДЫЃЌЗХФСИЩШХЧПЖШЖдВнЕиЩњЬЌЯЕЭГЕФгАЯьЪЧвЛИіНЅНјЕФЁЂСЌајЕФБфЛЏЙ§ГЬЃЌЕЋЕМжТВЛЭЌЮШЬЌзЊЛЏЕФуажЕОпгаВЛСЌајЬиеїЃЌЕМжТИпКЎВнЕщВЛЭЌЮШЬЌМфЕФзЊЛЏБэЯжЮЊдОБфадЖјЗЧНЅБфадЁЃдкЧрВиИпдКЎРфИЩдяЕФЦјКђЬѕМўЯТЃЌИпКЎсдВнВнЕщОпгаНЯИпЕФжВЮяШКТфЮЂАпПщЖрбљадЃЌОіЖЈИпКЎсдВнВнЕщЭЫЛЏГЬЖШЕФЪЧЮЂАпПщЕФБШР§КЭИёОжЃЌЖјВЛНіНіЪЧЯЕЭГжВЮяШКТфЮяжжЗсИЛЖШЁЃНќ70ФъРДЃЌШЫЮЊЛюЖЏдкжаЙњИпКЎсдВнВнЕщЩњЬЌЯЕЭГЬиеїдйЫмдьКЭзЪдДбнЛЏЙ§ГЬжаЗЂЛгСЫживЊзїгУЃЈYang et al., 2016ЃЉЁЃдкаЁЪБПеГпЖШЩЯЃЌИпКЎВнЕщЭЫЛЏЕФИљБОдвђЪЧШЫРрЖдЩњЬЌЯЕЭГЕФВЛКЯРэРћгУЃЌЙ§ЖШЗХФСВЛПЩБмУтЕиИФБфСЫПЩЪГадФСВнЮЂАпПщЕФЪ§СПЃЌНјЖјНЕЕЭСЫИпКЎсдВнВнЕщЩњЬЌЯЕЭГЖдЧјгђОМУКЭЩчЛсЗўЮёФмСІЃЈWang, 2004ЃЉЁЃвђДЫЃЌОЁЙмЛљгкЮЂАпПщЯЕЭГЕФИпЩНсдВнВнЕщЗўЮёЙІФмЕФЦРЙРПЩвдУїЮњВнЕиЕФбнЛЏЙ§ГЬЃЌжЦЖЈЭЫЛЏЮШЬЌаоИДФЃЪНЃЌЕЋНтОіИпЩНсдВнВнЕщЭЫЛЏЮЪЬтЕФИљдДдкгкВнЕиЙмРэЁЃ
НсТл
ЃЈ1ЃЉЙ§ЖШЗХФСНЕЕЭСЫИпКЎВнЕщЕФГѕМЖЩњВњСІЃЌЕЋЩњЬЌЯЕЭГПЩвдЭЈЙ§ЖджВЮяШКТфЕФзщГЩКЭНсЙЙЕФЕїНкЕжЯћВПЗжЗХФСИЩШХЕФгАЯьЃЌЮЌГжЯЕЭГЕФЩњВњФмСІЕФЯрЖдЮШЖЈЁЃЃЈ2ЃЉЙ§ЖШЗХФСПЩвдЭЈЙ§ИФБфЮЂАпПщжаПЩЪГадКЭВЛПЩЪГадФСВнЕФБШР§РДИФБфжВЮяШКТфЕФзщГЩКЭНсЙЙЃЌв§Ц№ЯЕЭГбнЬцЮШЬЌЕФзЊБфЃЌЖјжВЮяШКТфЮЂАпПщбнЛЏгыЯЕЭГЮШЬЌжЎМфЕФаЭЌБфЛЏКмДѓГЬЖШЩЯЮЌГжСЫЧрВиИпдИпКЎсдВнВнЕщЕиЩЯЩњЮяСПЕФЯрЖдЮШЖЈЁЃ/doi/10.1002/glr2.120527
ЭГГяЃКЭѕаТгю ЩђНѕЛл ЧидѓЦН
ЩљУїЃКИУЮФНіДњБэБрвыепЖдТлЮФЕФРэНтЃЌШчашВЮПМКЭв§гУЯрЙиФкШнЃЌЧыВщдФдЮФЁЃ

ЦкПЏНщЩм
Grassland ResearchЪЧЮвЙњВнвЕПЦбЇСьгђЕквЛБОЙњМЪбЇЪѕЦкПЏЃЌМОПЏЃЌгЩжаЙњВнбЇЛсКЭРМжнДѓбЇЙВЭЌжїАьЁЃИУПЏЪмжаЙњПЦММЦкПЏзПдНМЦЛЎИпЦ№ЕуаТПЏЯюФПжЇГжЃЌгЩЙњМЪГіАцМЏЭХJohn Wiley & Sons Australia, Ltd.ЬсЙЉГіАцМАаћДЋЗўЮёЃЌгк2022Фъе§ЪНГіАцЁЃ
Grassland ResearchТлЮФПЏЗЂЗЖЮЇЙуЃЌзлКЯадЧПЁЃДгЗжзгЕНШЋЧђБфЛЏВуУцЃЌШЋЮЌЖШОлНЙВнвЕПЦбЇМАЦфдкШЫРрПЩГжајЗЂеЙжаЕФзїгУЁЃЦкПЏНЋПЏЕЧЬьШЛВндЃЌддХрВнЕиЁЂВнЦККЭЩњЮяФмдДзїЮяЃЌвдМАВнЕиЩњЬЌЯЕЭГШ§ДѓАхПщЕФЛљДЁадКЭгІгУадбаОПГЩЙћЁЂзлЪіЁЂТлЕуЕШРраЭЕФЮФеТЁЃгХЯШПМТЧЗЂБэЧрФъбЇепгХаубаОПГЩЙћЃЌЦкД§ГЩЮЊЧрФъПЦбЇМвЯВАЎЕФЙњМЪбЇЪѕНЛСїжїеѓЕиЁЃ
дкДДПЏЧАШ§ФъЃЌGrassland ResearchНЋУтЪеАцУцЗбЃЌвдOAаЮЪНЭЈЙ§ШЋЧђЛЏГіАцЦНЬЈWiley Online LibraryГіАцЁЃ
https://blog.sciencenet.cn/blog-3568995-1422768.html
ЩЯвЛЦЊЃКGrassland Research | вьжЪВнЕиАпПщжаЕФЭСШРгаЛњЬМДЂСПКЭЕиЯТЩњЮяСП
ЯТвЛЦЊЃКШ№бЉезЗсФъЃЁGrassland ResearchдИФњбЇвЕНјВНЃЌЪТвЕгаГЩ!
ШЋВПзїепЕФЦфЫћзюаТВЉЮФ
- • GRжїБр | ФЯжОБъдКЪПГіЯЏЛФФЎЛЏзлКЯЗРжЮКЭЁАШ§ББЁБЙЄГЬНЈЩшбЇЪѕбаЬжЛс
- • ПЦЦе | жЊВнЁЂАЎВнЁЂЛЄВн 6ЁЄ18ВндБЃЛЄШе
- • GRЛяАщ | Ек26НьЙњМЪВнЕиДѓЛсЁЊ100 Years of Grassland Research
- • Grassland ResearchмйоЃбаОПЮФеТКЯМ
- • Grassland Research | ЖЋЗЧИФСМФСВнжжзгЪаГЁЕФЗЂеЙЁЂЦПОБКЭЛњгіЃКПЯФсбЧАИР§
- • Grassland Research | УРЙњЖЋФЯВПЗХФСЕиЭСШРЬМЙЬДцЕФдіМгЛњЛс
ШЋВПОЋбЁВЉЮФЕМЖС
- • ПЦбЇЭј2024Фъ6дТЪЎМбВЉЮФАёЕЅЙЋВМЃЁ
- • РюРМОъдКЪПЭХЖгЃКамШЅбѕЕЈЫсКЭМфГфжЪИЩЯИАћдкжЮСЦЮЃжижЂCOVID-19ЛМепжаЕФМлжЕ | Microorganisms
- • ЫежнДѓбЇаьдѓЮФ/ЛЦГЬЕШЃКЛљгкбЙЕчадКкСзЯЉ/Ti₃C₂TₓвьжЪНсдіВФжЦдьЕФЗЧЖдГЦжЏЙЙФЄЕчМЋгУгкдіЧПЕчЛЏбЇДЂяЎЮШЖЈ ...
- • 2024ФъЯФМОЧрВиИпдПМВьЃКМЊТЁЙЕРЅГцЖрбљадМрВт-0716
- • ГрзгжЎаФЁЂОЋжвБЈЙњ--УхЛГз№ОДЕФЕМЪІзЏг§жЧЯШЩњ
- • бЇЯАМЧвфЕФБэЙлвХДЋЛњжЦ