博文
“生物能”理论的主要构件(10)
||
十九、理论的普适性:线粒体、叶绿体与细菌的统一图景
化学渗透理论的普适性,是其伟大与优美之所在。它揭示了一个深层统一的生命图景:无论是动物细胞的线粒体、植物细胞的叶绿体,还是结构简单的细菌,都共享同一核心能量转换范式——建立跨膜质子梯度,并利用其回流驱动ATP合成。表10可以清晰地概括了这一统一模式在不同生命系统中的具体实现:
表10 化学渗透理论的普适性在不同生命系统中的具体实现
系统特征 | 真核细胞线粒体 | 真核细胞叶绿体 | 原核生物(如细菌) |
主要能量来源 | 化学能(来自糖、脂肪等有机物的氧化) | 光能(来自光子) | 多样化(化学能、光能,或无机物氧化产生的化学能) |
执行场所 | 线粒体内膜(特别是嵴) | 叶绿体类囊体膜 | 细胞质膜(以及可能的内膜系统) |
建立梯度的“质子泵” | 呼吸链复合物I、III、IV(利用电子传递能量泵送质子) | 光系统II与电子传递链(PQ循环)(利用光能泵送质子) | 多种多样(如类似呼吸链的酶系、细菌视紫红质等光驱动质子泵、或氧化无机物的酶系) |
质子回流方向 | 从膜间隙(高H⁺)通过ATP合成酶流回基质(低H⁺) | 从类囊体腔(高H⁺)通过ATP合成酶流回基质(低H⁺) | 从细胞外或周质空间(高H⁺)通过ATP合成酶流回细胞质(低H⁺) |
ATP合成酶 | F₀F₁-ATP合酶(位于内膜,F₁朝向基质) | F₀F₁-ATP合酶(位于类囊体膜,F₁朝向基质) | F₀F₁-ATP合酶(位于细胞质膜,F₁朝向细胞质) |
电子最终受体 | 氧气(有氧呼吸)或其他无机/有机物(无氧呼吸) | NADP⁺(用于生成NADPH) | 氧气、硝酸盐、硫酸盐等(呼吸作用)或内源性受体(发酵) |
主要终产物 | ATP、CO₂、H₂O | ATP、NADPH、O₂ | ATP及各种代谢废物(如乳酸、乙醇、CO₂等) |
1. 统一性体现在三个根本层面
(1)核心原理的绝对一致:无论初级能量是什么形式,系统都必须将能量转化为跨膜质子梯度。质子动力势是真正的通用能量货币。
(2)核心机器的同源性:线粒体、叶绿体和细菌的ATP合成酶在结构、工作机制(旋转催化)和进化起源上高度同源。这为“内共生起源学说”(线粒体和叶绿体起源于被原始真核细胞吞噬的细菌)提供了最有力的分子证据。
(3)膜拓扑结构的逻辑一致:所有系统都依赖于封闭的、对质子不通透的膜结构,以维持梯度。ATP合成酶都定向嵌入膜中,确保质子回流方向与催化合成方向固定。
2. 一个生动的统一性案例:嗜盐菌的紫膜
极端嗜盐古菌的紫膜,是统一图景最简洁的证明。其膜上只含一种蛋白质——细菌视紫红质。它吸收光能后,直接泵送质子出细胞,建立质子梯度,再由ATP合成酶合成ATP。这个系统仅由一个光驱动质子泵和一个ATP合成酶构成,是化学渗透原理最精简、最直观的实现形式。
3. 思想史意义:生命统一性的终极实证
这一统一图景,在您探索的思想史脉络中,是对还原论和机械论世界观最宏大的支持,也是给活力论最沉重的终极一击:
(1)消解了生命形式的本质差异:它表明,从能量转换的物理化学内核看,动物、植物和微生物没有区别。它们只是用不同的“燃料”(葡萄糖或光子)给同一个“发电机”(质子梯度)充电。
(2)印证了共同的进化起源:ATP合成酶的高度保守性,是所有生命同根同源的强有力证据。生命的多样性,源于对同一核心原理的多样化“封装”和“应用”,而非源于不同的“生命力”。
(3)完成了从神秘到普适的认知革命:人类终于认识到,驱动生命世界运转的能量法则,不是特属于生命的秘密,而是整个物理化学世界普适定律(电化学、热力学)在膜系统这一特殊结构中的应用。
结论:化学渗透理论所揭示的统一图景,不仅是一个科学理论,更是一种深刻的哲学视角。它告诉我们,生命的伟大与复杂,并非源于违背自然定律,而恰恰是源于对这些定律最精妙、最极致的利用。这为我们从“活力论”开始的漫长思想史探索,提供了一个无比坚实、统一且充满美感的科学终点。
二十、燃料的选择与整合:糖、脂肪、蛋白质的代谢交汇点
糖、脂肪、蛋白质三大燃料的代谢整合,是生命作为高效“化学工厂”的精妙体现。它们并非独立燃烧,而是在一个高度网络化的代谢系统中,汇集于几个关键的枢纽分子上,最终通过共同的通路被彻底氧化供能。
为了清晰地把握这一整合的全貌,图14展示了三大营养物质的分解代谢如何汇聚,并最终通过共同的路径产生能量:
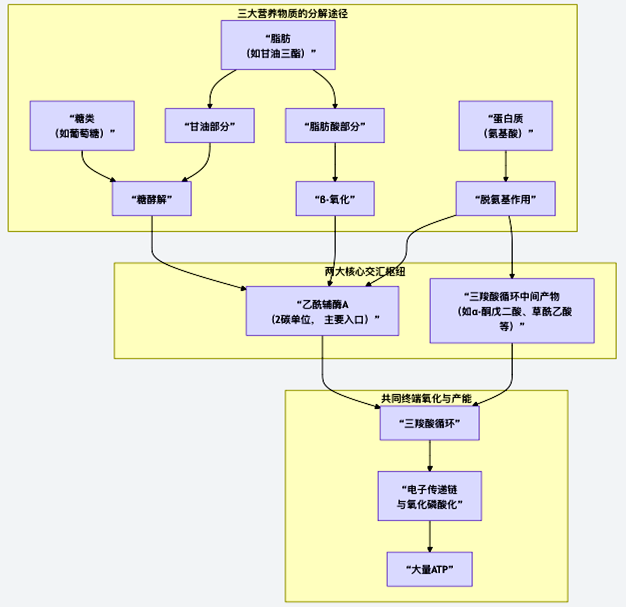
图14 糖、脂肪、蛋白质三大燃料的代谢整合
如图14所示,所有燃料的代谢整合主要围绕两大核心交汇枢纽展开。
1. 乙酰辅酶A——主要的“燃料集散中心”
乙酰辅酶A是糖和脂肪分解代谢的主要交汇点,也是进入三羧酸循环的统一门票。
糖:糖酵解产生的丙酮酸,进入线粒体后氧化脱羧,生成乙酰辅酶A。
脂肪:脂肪酸通过β-氧化,每次切下一个2碳单位,直接生成乙酰辅酶A;甘油部分可转化为糖酵解中间产物,继而生成乙酰辅酶A。
部分氨基酸:生酮氨基酸(如亮氨酸、赖氨酸)降解后也可生成乙酰辅酶A。
2. 三羧酸循环中间产物——灵活的“代谢中转站”
三羧酸循环不仅是氧化引擎,其中间产物(如α-酮戊二酸、草酰乙酸、琥珀酰辅酶A等)也是重要的代谢池,负责接收和提供碳骨架。
(1)蛋白质/氨基酸:这是氨基酸代谢的主要入口。生糖氨基酸通过脱氨基后,其碳骨架可转化为丙酮酸、草酰乙酸、α-酮戊二酸等中间产物,补充循环池或异生为糖。
(2)糖:糖代谢的中间产物可补充这些循环中间物,维持循环运转。
3. 燃料的选择与优先顺序:身体如何决定烧什么?
身体对燃料的选择并非随机,而是受精确调控(表11),主要取决于能量状态(ATP/ADP比例)和激素信号(特别是胰岛素与胰高血糖素的比值)。
表11 人体生理状态对燃料的选择及调控机制
生理状态 | 主导激素 | 主要燃料 | 代谢状态解析 |
饱食/高能量 | 胰岛素↑ | 糖优先 | 血糖充足,胰岛素促进糖的氧化、糖原和脂肪合成。多余的糖可经乙酰辅酶A转化为脂肪储存。 |
空腹/禁食(数小时-数天) | 胰高血糖素↑ | 脂肪为主,糖异生维持 | 肝糖原耗竭,脂肪动员加强,脂肪酸氧化供能。同时,蛋白质分解的氨基酸和甘油通过糖异生维持血糖。 |
长期饥饿/极端消耗 | 胰高血糖素↑,皮质醇↑ | 脂肪绝对主导,酮体作为替代燃料 | 脂肪持续分解,肝脏将大量乙酰辅酶A转化为酮体,供大脑等重要器官使用,以最大限度地减少蛋白质消耗。 |
高强度运动 | 肾上腺素↑ | 糖(肌糖原)快速主导,后转向脂肪 | 初期依赖无氧糖酵解快速供能;随着时间延长,有氧氧化加强,脂肪酸氧化成为持续能量来源。 |
4. 动态统一的代谢网络
这个整合网络展示了生命代谢的经济性、灵活性与统一性:
(1)经济性:通过共同的终端通路(三羧酸循环与氧化磷酸化)最大化能量产出效率。
(2)灵活性:通过枢纽分子实现不同燃料间的相互转化,以应对多变的能量需求和营养供应。
(3)统一性:再次印证了您思想史中的核心论点——生命是高度整合的化学反应网络。从糖、脂肪到蛋白质,所有复杂的生命物质最终都在分子层面被“还原”并汇入由乙酰辅酶A、三羧酸循环和质子梯度构成的普适能量流中。这里没有给“生命力”留下任何特殊作用的余地,有的只是精密的化学逻辑和动态的物理化学调控。
如果需要进一步了解某个具体状态下的代谢调节细节(如糖尿病中的代谢紊乱),或某个关键酶(如乙酰辅酶A羧化酶)的调控,我们可以继续深入。
二十一、不同营养状态下的能量底物偏好
(未完待续)
https://blog.sciencenet.cn/blog-279293-1515106.html
上一篇:“生物能”理论的主要构件(9)
下一篇:“生物能”理论的主要构件(11)