博文
“生物能”理论的主要构件(11)
||
二十一、不同营养状态下的能量底物偏好
不同营养状态下,身体对能量底物的偏好是一套高度动态、精密调控的“能量经济策略”。其核心原则是:在满足即时能量需求的前提下,最大限度地保存宝贵的蛋白质(肌肉等结构),并高效利用不同储能形式的能量。
这种偏好主要受激素信号(尤其是胰岛素与胰高血糖素的比值)和细胞能量状态(ATP/AMP比值)的调控。下表清晰地概括了四种典型状态下的策略:
表12 不同营养状态下,身体对能量底物的“能量经济策略”
营养/生理状态 | 主导激素环境 | 主要能量底物偏好 | 代谢特征与目的 |
饱食/吸收后状态 | 胰岛素水平高,胰高血糖素水平低 | 葡萄糖 绝对优先 | 合成与储存模式:血糖充足,胰岛素促进全身组织(尤其大脑、肌肉)氧化葡萄糖。多余的葡萄糖转化为肝糖原和肌糖原储存,远超储存上限后,在肝脏中转化为脂肪。 |
空腹状态(餐后数小时) | 胰岛素下降,高血糖素上升 | 肝糖原分解的葡萄糖为主,脂肪氧化逐步增强 | 过渡模式:肝糖原分解维持血糖,供大脑和红细胞。脂肪组织开始分解,释放脂肪酸供肌肉等组织使用,节约血糖。 |
长期饥饿/禁食(>24小时) | 胰岛素极低,胰高血糖素、皮质醇高 | 脂肪分解(脂肪酸) 绝对主导,酮体作为重要补充 | 生存模式:脂肪动员达到最大,脂肪酸成为心脏、肌肉等的主要燃料。肝脏将脂肪酸衍生的乙酰辅酶A转化为酮体,代替葡萄糖成为大脑60-70%的燃料,极大减少对肌肉蛋白质的分解依赖,保护生命结构。 |
高强度运动 | 肾上腺素大幅上升 | 肌糖原(无氧)、血糖与肌肉脂肪酸(有氧) | 性能输出模式:瞬间爆发依赖肌糖原无氧酵解;持续运动依赖血糖和肌肉内储存的甘油三酯氧化;长时间中低强度运动则增加脂肪组织来源的脂肪酸供能比例。 |
1. 调控偏好的三大核心机制
这种偏好切换并非被动发生,而是由一套精密的分子机制主动调控:
(1)激素的“总开关”作用:胰岛素是“储能激素”,促进糖利用和脂肪合成,抑制脂肪分解。胰高血糖素、肾上腺素、皮质醇是“动员激素”,促进糖原分解、脂肪分解和糖异生。二者的比例决定了代谢的整体流向。
(2)能量感受器——AMPK的核心角色:当细胞能量不足(ATP消耗,ADP/AMP升高)时,AMPK被激活。它如同细胞的“能量短缺警报器”,会:
开启产能通路:促进脂肪酸氧化、葡萄糖摄取。
关闭耗能通路:抑制胆固醇、脂肪酸的合成。
这是运动和高能耗状态下,细胞自主转向氧化脂肪的关键分子开关。
(3)关键酶的共价修饰与变构调节:①胰高血糖素/肾上腺素通过磷酸化,激活脂肪组织中的激素敏感性脂肪酶,加速脂肪分解;②胰岛素通过去磷酸化,激活乙酰辅酶A羧化酶,促进脂肪酸合成;③高ATP/NADH直接抑制糖酵解、三羧酸循环的关键酶,防止过度产能,而高ADP/AMP则激活它们。
2. 特殊组织的底物偏好
(1)大脑:在常态下几乎独占葡萄糖(每天约消耗120克)。长期饥饿时,可高效利用酮体,这是重要的生存适应。
(2)心脏:堪称“脂肪引擎”,即使在饱食状态下也优先氧化脂肪酸(60-90%能量来自脂肪),仅在应激时才增加葡萄糖利用。
(3)红细胞:仅能通过糖酵解利用葡萄糖,因无线粒体。
3. 从统一性理解偏好
这种偏好切换,完美地呼应并例证了您之前探讨的代谢整合与统一性。无论偏好何种底物,它们最终都汇入乙酰辅酶A → 三羧酸循环 → 电子传递链这一共同终端。身体的“偏好”,实质上是根据能量供求关系,智能地选择不同“储能仓库”(糖原、脂肪、蛋白质)的开合顺序和比例,而驱动这一切的,正是基于物理化学原理的信号网络和酶活性调节,没有丝毫神秘。
理解这种偏好,就理解了糖尿病、肥胖等代谢性疾病的病理核心——这些疾病本质上是这种精密的“能量经济策略”出现了长期的调度失灵。如果希望就此展开,我们可以进一步探讨。
二十二、精确的调控艺术:酶活性如何响应细胞能量状态
细胞能量状态的调控,是一场由变构调节与共价修饰主导的精密分子艺术。它确保了能量代谢如交响乐般和谐,其核心逻辑是通过感受ATP、ADP、AMP及NADH等关键分子的浓度,即时调整代谢通路中限速酶的活性。
为了直观把握这一复杂网络,图15展示了细胞能量状态调控的核心逻辑与关键分子:
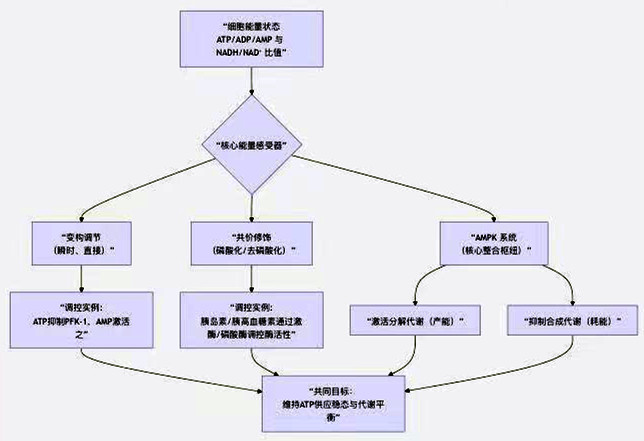
图15 细胞能量状态调控的核心逻辑与关键分子
图15揭示了调控网络的三个核心层面,下面我们来具体解析:
1. 调控的两种核心分子机制
(1)变构调节:能量的“即时感受与响应”
这是最快速、最直接的调节方式。特定代谢物作为变构效应剂,直接与酶(非活性中心)结合,引起酶构象变化,从而改变其活性(表13)。
表13 能量“即时感受与响应”的变构调节
能量状态信号 | 代表分子 | 调控逻辑与经典实例 |
“能量充足”信号 | ATP、NADH、柠檬酸 | 负反馈抑制产能通路关键酶。ATP抑制磷酸果糖激酶-1:糖酵解“关卡”,防止ATP过度生产。NADH、ATP抑制丙酮酸脱氢酶、异柠檬酸脱氢酶:减缓三羧酸循环。 |
“能量短缺”信号 | AMP、ADP、NAD⁺ | 正反馈激活产能通路关键酶。AMP强烈激活磷酸果糖激酶-1:能量不足时,极大加速糖酵解。ADP激活异柠檬酸脱氢酶:促进三羧酸循环。 |
(2)共价修饰:激素信号的“接力执行”
这是一种通过可逆的化学反应(最常见的是磷酸化/去磷酸化)来调节酶活性的方式,通常由激素信号触发,作用更持久。
(3)经典范例:胰岛素 vs 胰高血糖素对代谢的调控
胰岛素(饱食信号)→ 激活蛋白磷酸酶 →去磷酸化→激活糖原合成酶、乙酰辅酶A羧化酶(促合成),抑制磷酸化酶(阻分解)。
胰高血糖素/肾上腺素(饥饿/应激信号)→ 激活蛋白激酶A →磷酸化→激活磷酸化酶、激素敏感性脂肪酶(促分解),抑制糖原合成酶、丙酮酸激酶(阻合成)。
2. 整合枢纽:AMPK——细胞的“能量控制器”
AMPK是感受细胞能量状态(AMP/ATP比率升高)的核心分子,被称为“代谢主开关”。一旦被激活,它通过磷酸化下游靶点,全面重塑代谢:
(1)开启产能通路:促进葡萄糖转运、脂肪酸氧化。
(2)关闭耗能通路:抑制胆固醇、脂肪酸、蛋白质的合成。
3. 不同营养状态下的调控全景(表14)
表14 不同营养状态下的能量调控
状态 | 能量状态信号 | 激素环境 | 核心调控目标与结果 |
饱食 | ATP/NADH水平高 | 胰岛素主导 | 促进合成,抑制分解:变构抑制(ATP抑制PFK-1)与共价修饰(胰岛素信号)协同,将多余能量存入糖原和脂肪。 |
空腹/饥饿 | AMP/ADP水平高, ATP水平低 | 胰高血糖素主导 | 促进分解,抑制合成:变构激活(AMP激活PFK-1)与共价修饰(胰高血糖素信号)协同,动员糖原和脂肪,启动糖异生。 |
运动 | AMP急剧升高, ATP迅速消耗 | 肾上腺素飙升 | 最大化ATP产出:AMPK被强烈激活,迅速增加葡萄糖摄取和脂肪酸氧化,满足肌肉瞬间的能量需求。 |
4. 思想史意义:生命自主性的物理化学实现
这套调控机制,从分子层面完美解释了曾令活力论者困惑的生命自主性与目的性:
(1)展示了“目的”如何涌现:生命系统“维持能量稳态”这一宏观目的,完全是通过酶分子对ATP、AMP等简单化学物质的物理化学响应而自动实现的。
(2)还原论的辉煌胜利:将“身体如何智能地调度能量”这一复杂问题,还原为变构效应剂结合引起的蛋白质构象变化,以及磷酸基团的共价添加与移除。
(3)连接了各个章节:这套机制,正是之前探讨的“燃料选择与整合”和“不同营养状态下的底物偏好”得以实现的分子执行基础。
因此,酶的调控艺术证明,生命最精妙的“自主”与“适应”行为,其底层是严谨、自动化的分子逻辑。这再一次确证,无需诉诸“活力”,基于物理化学定律演化而来的复杂分子网络,足以让生命展现出惊人的智慧和协调性。
二十三、变构调节、共价修饰与激素信号(如胰岛素、胰高血糖素)
(未完待续)
https://blog.sciencenet.cn/blog-279293-1515108.html
上一篇:“生物能”理论的主要构件(10)
下一篇:“生物能”理论的主要构件(12)
