博文
人等宿主如何控制体内微生物组的机制  精选
精选
|
背景
许多多细胞生物,包括人类,都携带共生微生物群落。这些微生物群具有防止感染、提供营养、促进免疫发育甚至影响认知的能力,对健康至关重要。
微生物组的许多潜在好处 - 微生物群加上相关的宿主因素 - 可以培养这样一种观点,即人类在肠道中拥有一个内在有用的生态系统,主要由饮食调节。实际上,微生物群的成员在每天大量细胞被排出的环境中面临着对资源和物理定位的长期竞争。新菌株的移民,以及快速和短视的微生物进化,对贪婪生长但伤害宿主的共生体构成了持续的威胁,包括病原体和“作弊者”基因型,它们投资于自己的生长而不是使宿主受益。为了应对这种压力,宿主已经进化出广泛的控制机制,不断监测和操纵它们的共生体,以限制伤害并促进所获得的好处。因此,人类微生物组等例子可以被概念化为“束缚的生态系统”,其中共生体在不同的生态群落中相互作用,这些群落受到宿主控制的强烈影响。
进展
在这篇对不同文献的综述中,我们展示了宿主控制机制如何影响微生物组生物学的几乎每个方面。我们的重点是哺乳动物微生物组,但我们也研究了来自无脊椎动物和植物微生物组的充分特征的例子。首先,我们探讨了宿主控制的生物学,包括免疫、屏障功能、生理稳态、转运和宿主行为。许多宿主控制机制的预期结果是促进有益的共生体而不是有害的共生体(伴侣选择),但控制也可以通过改变共生体的行为和新陈代谢来使它们更有益(伴侣操纵)。如果微生物的快速进化能够产生对微生物组内有益性状的自然选择,那么它就可以成为宿主控制的机会。然而,如果共生体进化使共生体能够逃避宿主的控制,那么共生体的进化也可能是一个问题,一些微生物已经找到了利用和运用控制机制来对付我们的方法,对健康产生灾难性的影响。
展望
人们对微生物组为其宿主提供的好处非常感兴趣。在这里,我们认为这些好处中的许多可以追溯到宿主控制机制。数亿年来,多细胞宿主一直处于自然选择之下,以增加其共生体的益处,其结果是一系列惊人的适应能力,能够控制多样化和高度可进化的微生物群落。认识到微生物组是宿主控制和共生体进化之间永久紧张关系的产物,有助于理解微生物组生物学和疾病的大部分内容。事实上,宿主控制机制对复杂微生物群的力量表明,其中许多微生物群将提供治疗靶点来重塑微生物组,从而改善健康。
Jacob Wilde et al. ,Host control of the microbiome: Mechanisms, evolution, and disease.Science385,eadi3338(2024).
许多物种,包括人类,都是共生微生物群落的宿主。关于这些微生物组如何影响宿主的方式有大量文献,但在这里,我们主张更多地关注宿主如何影响其微生物组。宿主通过多种机制控制其共生体,包括免疫、屏障功能、生理稳态和运输。这些机制使宿主能够塑造微生物组的生态学和进化,并产生有益于宿主的微生物性状的自然选择。我们的微生物组是宿主控制和共生体进化之间永久紧张的结果,我们可以利用宿主的进化能力来调节微生物群来预防和治疗疾病。宿主控制的研究对于我们理解和操纵微生物群以改善健康的能力至关重要。
 图示 夏威夷短尾鱿鱼光器官选择生物发光细菌。许多动物避免腐烂的食物以减少病原体定植。一些豆科植物在根瘤中容纳固氮细菌,并通过切断固氮太少的根瘤的营养来控制它们。大熊猫是一种最近进化的食草动物,其肠道微生物群缺乏有效分解植物材料的酶,是宿主控制不佳的一个例子。
图示 夏威夷短尾鱿鱼光器官选择生物发光细菌。许多动物避免腐烂的食物以减少病原体定植。一些豆科植物在根瘤中容纳固氮细菌,并通过切断固氮太少的根瘤的营养来控制它们。大熊猫是一种最近进化的食草动物,其肠道微生物群缺乏有效分解植物材料的酶,是宿主控制不佳的一个例子。
许多多细胞生物,包括人类,都携带微生物群落,或在其上皮表面有与微生物群落相关联。一个生物的微生物群(microbiota),凭借其抵御感染、提供营养、促进免疫发展乃至影响认知能力的能力,对于健康功能至关重要。微生物组(microbiome)定义为微生物群加上相关的宿主因素,其众多潜在益处可以促成一种观点,即人类在他们的肠道内寄居着一个本质上有益的生态系统,主要由饮食调节。实际上,微生物群的成员面临着资源的长期竞争和物理定位的竞争,在一个每天都有大量细胞被排出的环境中。新菌株的迁移,以及快速而短视的微生物进化,构成了共生体贪婪生长却伤害宿主的持续威胁。作为回应,宿主已经进化出了一系列机制,不断地监视和操纵它们的共生体,以限制伤害并促进所接受的益处。因此,例如人类的微生物组可以被概念化为“一个受约束的生态系统”,其中共生体在由宿主控制强烈塑造的多样化生态社区中相互作用。
许多与宿主相关的微生物组似乎为宿主提供益处,以换取营养和一个相对稳定的环境。人们可能会假设这些互惠互利本身就足以解释宿主与其微生物群之间的合作,但进化文献强烈警告不要做出这种假设。例如,考虑一个投资于生产供养宿主的产品的共生体。尽管宿主受益,但预计不生产该产品而是使用资源进行生长的微生物将会繁荣(有时在进化生物学中被称为“骗子”)。共生体的快速世代时间可以通过允许短视的共生体进化在宿主一代内发生来极大地加剧这个问题。
方框1 宿主控制机制可以通过产生对合作的自然选择来逆转这一进化轨迹。这一结果要求宿主找到方法来促进有益基因型胜过骗子和病原体。一种方法是在特殊的器官或组织中隔离有益共生体的克隆群体,在那里可以监控和调节行为——固氮细菌在根瘤中的隔离和发光细菌在乌贼光器官中的隔离就是两个这样的例子。另一种方法是通过位置识别有害基因型:免疫系统通常对突破上皮屏障的微生物和那些没有这样做的微生物的反应非常不同。然而,在一些情况下,这样的空间信息是缺乏的。例如,大肠的腔主要通过创造营养生态位来运作。在这里,厌氧环境和复杂碳水化合物的提供有利于将碳水化合物发酵成宿主可以利用的产物的微生物,以至于缺乏这种能力的微生物可能处于竞争劣势。当与其他机制结合时,如适应性免疫系统能够用IgA针对特定基因型,这种环境控制似乎足以调节腔微生物组。然而,宿主如何有效地控制如此多样化的微生物组并不总是清楚的,这是未来探索的一个重要方向。
在本综述中,我们展示了宿主如何控制影响微生物组生物学机制的几乎每一个方面。我们的重点是哺乳动物的微生物组,但也考察了来自无脊椎动物和植物微生物组的明确例子。首先,我们探索了宿主控制的生物学,包括免疫反应和其他方面的宿主生理学。这些机制通过改变共生体组成、改变共生体表型或两者兼而有之来影响微生物组。微生物组的快速进化对宿主来说既是一个机会,如果他们可以为有益特征生成自然选择,也是一个问题,如果它使共生体能够逃避宿主控制。宿主控制机制对复杂微生物组的力量表明,许多宿主因素将提供重新塑造微生物组以改善健康的治疗目标。
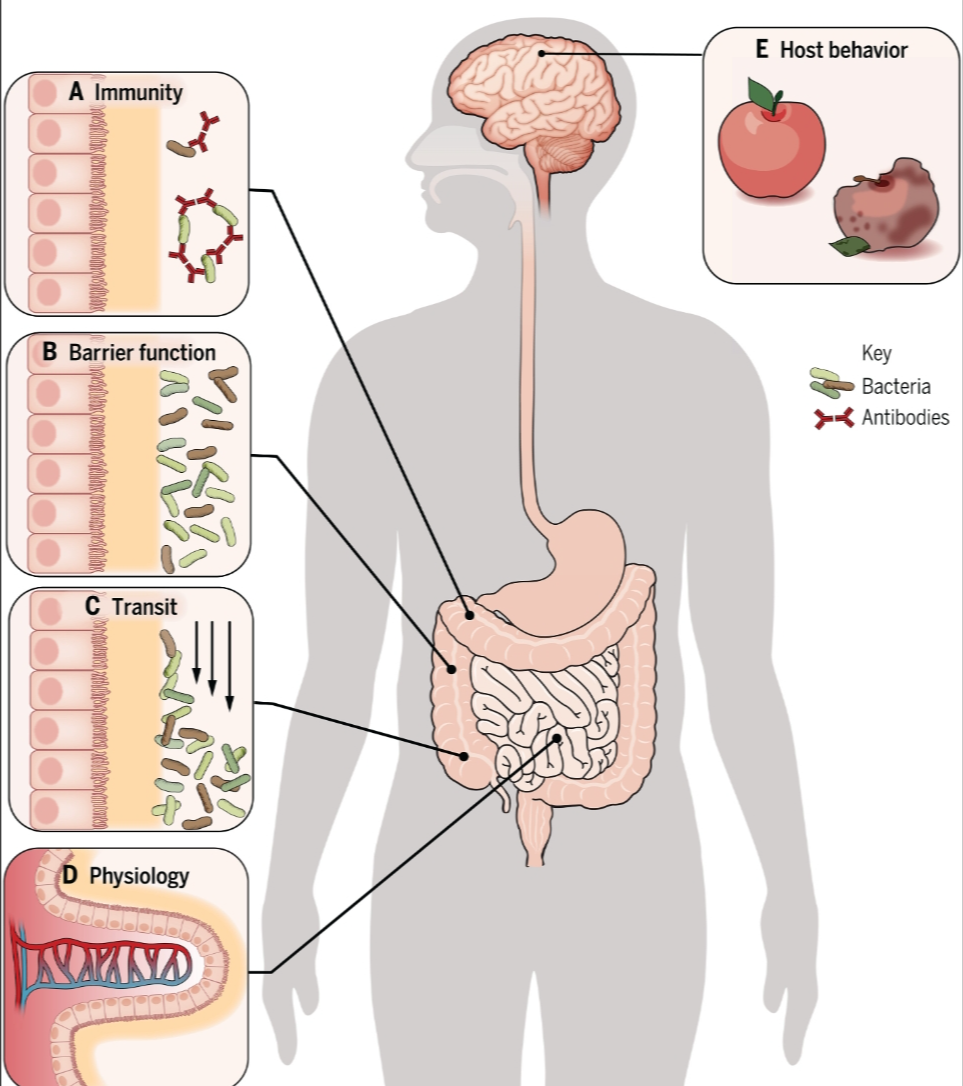
图1. 宿主对微生物组的控制机制。
(A) 免疫性:免疫系统能够迅速学习微生物表型与基因型之间的关联,是已知最复杂的宿主控制系统。在哺乳动物的肠道中,一个关键的行动机制是通过IgA,它可以帮助细菌附着在黏液上,也可以使细菌相互粘附形成可以被清除的细菌团块。(B) 屏障功能:宿主使用屏障来限制共生体。肠道黏膜是一个重要的屏障例子,它限制了微生物与宿主细胞的接触,但允许代谢物双向通过。(C) 过渡:宿主有能力通过如蠕动等机制在其微生物组内移动共生体,这可以迅速清除引起疾病的共生体。(D) 生理学:宿主以强烈影响其共生体的方式控制其微生物组的生理学,例如通过逆流血流限制氧气,促进微生物发酵。(E) 宿主行为:动物避免食用变质食物的能力可以帮助保持微生物组免受病原体侵害。
宿主控制特征
宿主与其微生物群之间的关系通常表现为合作或互惠共生,双方都能从中受益(方框1)。在这种背景下,宿主控制机制是进化生物学中所谓的“执行”的一个例子(10)。执行机制是至少部分为了减少合作联盟中的自私行为而进化的特征。证明执行重要性的例子比比皆是,包括基因组中转座元件的抑制、社会昆虫群体中的警察行为、拒绝无花蜜植物的传粉者以及人类的互惠利他主义(10)。因此,可以将宿主控制机制定义为那些至少部分为了抑制微生物组中有害或自私行为并促进宿主利益而进化的机制(3)(图1)。
多样的宿主特征可能会影响微生物群的组成和宿主获得的利益(方框2),但如果这些特征因其他原因而进化,它们就不构成有意义的宿主控制。例如,似乎不太可能因为宿主控制的需求而进化出肠道蠕动,而更可能是因为处理食物并最终排除废物的需要(11)。相比之下,感染时肠道过渡速率的快速变化确实是一种宿主控制机制,用于帮助清除问题微生物(12)。在某些情况下,不清楚某个特征是为了宿主控制目的还是其他原因而进化的,考虑到我们讨论的例子时应该记住这一点。我们的重点是“开放”的微生物组,其中每个宿主代内有大量移民和共生体的水平转移潜力(9)。一些共生体,如线粒体、叶绿体和昆虫的细胞内共生体,被功能性地整合到宿主细胞中并通过垂直传递,这可以减少对宿主控制的需求(4,6,9)。
方框2关于全基因组关联研究、宿主遗传学和宿主控制
有趣问题是,宿主遗传学是否可以解释个体间微生物群组成的变化,这可以通过全基因组关联研究来探究。有时这类研究没有发现效应(143),看起来这是一种评估宿主控制机制重要性的方法。但这个结论有问题,因为遗传关联依赖于具有相当大持续遗传变异和功能性后果的性状,对于最重要的宿主控制性状,如厌氧肠道,这种变异可能不存在,因为强烈的净化选择维持了性状的功能。
第二个问题是,微生物群组成是一个复杂且多变的表型,与宿主遗传学相关联(与疾病风险等相比)(144)。即使宿主基因型存在变异影响组成,例如筛选能够发酵的微生物,也可能无法检测到宿主遗传学的影响,因为不同的微生物可以为不同的宿主提供相同的功能。与此一致的是,由于细菌种内替换事件,人类微生物组中的微生物基因型随时间变化(121,145),但对宿主的功能效应可能不会。同样地,改变饮食可以改变肠道微生物群中分类单元的丰度,但这种转变并不意味着缺乏宿主控制,宿主控制只预期限制对宿主有负面影响的变化。然而,饮食引入的变异性再次可能混淆宿主遗传学与微生物组变异之间的任何联系。
最终的问题是,即使检测到宿主遗传学的影响,它们可能不代表宿主控制。在一项大型双胞胎研究中发现的关键关联是宿主乳糖酶表达与消耗乳糖的双歧杆菌丰度之间的负链接(146),这更好地解释为宿主对牛奶的饮食偏好而不是宿主控制。尽管关联研究并不直接评估宿主控制的重要性,但它们可能是理解某些控制机制的途径,因为一些确实显示出遗传变异。例如,人类中有一个常见的TLR5等位基因(它检测细菌鞭毛蛋白)似乎使其失去功能,这与增加细菌感染的风险(147)但降低自身免疫疾病的风险(148)有关。
一些宿主控制机制,如适应性免疫,可能对微生物群的某些成员高度特异,而其他机制,如缺氧,可能影响数百种物种(13)。我们以哺乳动物肠道微生物组作为关键例子,但许多机制和原则广泛适用,包括其他身体部位、其他脊椎动物(12,14,15)、无脊椎动物(16,17)、植物(5,18)甚至一些单细胞宿主(19)(图2)。

图2. 在多种物种中观察到宿主控制机制。
从一系列动物、植物和单细胞物种的实验研究中展示了宿主控制的证据。(A) 硅藻 Asterionellopsis glacialis 释放不寻常的代谢物,增强有益共生体的附着和生长,同时抑制机会性病原体(19)。(B) 豆类 Glycine max 切断对不固定氮的根瘤的营养供应(5)。(C) 十字花科植物 A. thaliana 编码的模式触发免疫反应对于维持正常微生物组至关重要(18)。(D) 各种水螅物种分泌抗菌肽,维持独特的微生物组(16)。(E) 琥珀甲虫 Xylosandrus germanus 偏爱树木内富含乙醇的环境,促进其真菌园共生体而非其他微生物(17)。(F) 斑马鱼 Danio rerio 的肠道过渡清除了宿主微生物组中竞争力较弱的菌株(14)。蠕动可能最初是为了消化功能而进化的,而不是宿主控制,但过渡速率的变化与宿主控制一致(12)(方框1)。(G) 二聚体IgA使微生物凝集并链接以从小鼠肠道清除(15)。
先天免疫和适应性免疫
脊椎动物免疫系统是已知最复杂的宿主控制机制。它包括先天免疫,这是进化上古老的,检测微生物和感染的保守特征,以及适应性免疫,仅存在于有颌脊椎动物中,可以在宿主的生命周期内生成新的受体以识别特定的微生物菌株。免疫系统的研究历史上集中在感染和疾病(20)。然而,越来越清楚的是,免疫机制更广泛地作用于形成和控制微生物组,特别是在关键的上皮表面,这些表面标志着微生物组与宿主本身之间的分界线(21)。
植物(21)和动物(22)的先天免疫与多细胞性一起进化,免疫组分可以追溯到古代原核生物防御病毒感染的机制(23)。动物和植物免疫之间的相似之处令人瞩目:两者都使用多样化的模式识别受体(PRRs),这些受体结合许多相同的常见微生物特征(20,21),或识别病原体毒力对宿主细胞功能的影响(22)。宿主PRRs与常见微生物特征的结合,称为病原相关分子模式(PAMPs)或微生物相关分子模式(MAMPs)(20,21),驱动一系列反应,这些反应可以重塑微生物组并帮助维持正常的宿主-微生物组关系(21,24)。
在动物中,一个关键的宿主受体类别是Toll样受体(TLRs),它们检测一系列细菌和病毒的PAMPs。理解TLRs如何区分具有相同表位的无害和有害微生物仍然是该领域的一个关键挑战。一种区分形式似乎基于位置:TLRs可以放置在通常不发现微生物的位置,如上皮层的面向宿主一侧(25),因此只有侵入性病原体才会激活它们。这种空间信息有助于免疫系统优先响应那些构成更大威胁的物种(方框1)。TLR信号传导在建立宿主与微生物组之间的关系中发挥关键作用,关键的TLR信号组件的遗传缺陷导致针对微生物组的异常适应性免疫(24)和由常见的微生物组成分驱动的侵袭性疾病(26)。除了监测微生物细胞的成分外,植物(27)和动物(28,29)的免疫系统还监测微生物组的代谢输出。先天免疫激活在健康的微生物组中具有重要功能,并与抗微生物肽表达相关(30),其中敲除关键抗微生物RegIIIγ,一种分泌的抗菌凝集素,导致微生物组侵犯小鼠的上皮层(31)。
抗微生物肽的重要性也在无脊椎动物中看到。在海洋无脊椎动物水螅中敲除生产,使黏膜微生物组组成远离自然状态(16),并且有证据表明,苍蝇物种进化出特定的抗微生物肽以匹配其环境中的细菌威胁(32)。吞噬细胞也是先天免疫的一个重要方面,它们在包括哺乳动物呼吸道和海绵中的地方吞噬并杀死微生物,并在其中大量漫游微生物组并似乎控制其组成(33)。尽管哺乳动物健康肠道腔内基本没有吞噬细胞,但在炎症肠道区域它们可以被大量释放,在那里它们通过分泌的DNA网捕获细菌,通过吞噬作用消除它们,并帮助加强肠道上皮屏障(34,35)。感染还可以驱动宿主细胞的程序性死亡和将受感染的宿主细胞物理排出到肠道腔中(36)。植物缺乏移动的免疫细胞和吞噬细胞,但包括程序性细胞死亡和抗微生物产品在内的机制同样用于限制和停止侵入性感染(21)。
脊椎动物的进化见证了动物免疫系统的精细化,包括了适应性免疫以及先天免疫机制。关键是,适应性免疫使系统能够学习和改变激活其受体的化学配体,这取决于相关威胁。系统通过不断从脊椎动物体表采样配体到黏膜相关淋巴组织中,通过微褶细胞和树突状细胞实现这一点(37)。尽管已经确定了调节性T细胞和白细胞介素-10信号的作用(37),但最终决定哪些抗原在被采样进入黏膜时产生何种反应仍然难以预测。明确的是,颗粒性抗原(即整个细菌、病毒)和细菌毒素在激活黏膜抗体反应方面非常有效。相反,一些可溶性抗原在被幼儿摄入时会诱导免疫耐受,这被认为是帮助成熟的免疫系统忽略许多无害的配体(38)。类似于先天免疫,抗体反应可能偏向于那些突破上皮屏障的物种,因为免疫监视和调控的关键部位在宿主组织内,即在引流淋巴组织内(37)(方框1)。
在健康的哺乳动物黏膜中,大量的浆细胞不断产生免疫球蛋白A(IgA)。这种二聚体抗体亚型通过多聚Ig受体积极分泌到上皮表面。分泌后,IgA缺乏其他免疫球蛋白的典型效应功能,如补体固定(39)。相反,IgA的许多功能可以通过其交叉连接抗原的能力来解释。特别是,通过高亲和力结合增加细菌聚集,提高粪便流中细菌清除的速率(15,39,40)。一些证据表明,IgA可以既作为胡萝卜又作为棍棒,例如,通过帮助细菌定殖于肠道上皮的黏液层(41)或保护它们免受胆汁酸和噬菌体的伤害(39,42)。IgA也在乳腺中产生,并随IgM一起通过牛奶传递给新生儿,人们认为它在成人中执行类似的功能。先天免疫成分,如哺乳动物尿道中的尿调蛋白(43)或中华绒螯蟹(Eriocheir sinensis)肠道中分泌的C型凝集素(44),也似乎主要通过聚集细菌来发挥作用,这表明这可能是黏膜中的一般保护机制。
许多研究调查了免疫对微生物组稳定性和组成的重要性。例如,在植物中,影响免疫信号和抗微生物反应的突变破坏了内生真菌Colletotrichum tofieldiae与拟南芥根部之间的有益共生关系(45),并且敲除与模式触发免疫相关的基因会导致叶片微生物组有害的组成变化(18)(图2)。在果蝇中,通过RNA干扰失调先天免疫会改变肠道中的共生体群落,导致一种肠道微生物占主导地位并增加宿主死亡率(46)。在小鼠中,数据可能受到宿主基因型、笼子效应和卫生隔离品系间微生物组漂移的影响,这在文献中产生了争议。例如,许多在小鼠中剔除特定免疫组分的研究未发现对微生物组组成有可检测的影响(31,47,48)。尽管如此,一些使用同窝仔或骨髓嵌合体以最小化潜在混杂因素的研究确实显示了免疫系统对微生物组组成的明确影响(49,50)。总之,在各项研究和不同宿主物种中,免疫对于塑造和调控微生物组很重要(16,18,31,44-46)。
屏障功能
限制微生物可以在哪里定殖和生长的屏障对于控制微生物组至关重要。一些屏障,如哺乳动物的皮肤,阻止微生物和其他几乎所有物质的通过。其他屏障,包括动物的黏膜上皮和植物的根上皮,充当选择性屏障,限制微生物的转运但允许化学交换。免疫有助于维持所有这些屏障。例如,豆蝽的肠道上皮由200多种抗菌肽加强(51)。屏障的渗透性也可以根据需要改变:紧密连接在哺乳动物上皮细胞之间形成密封,但在腹泻期间发生的大量水和盐分流失时可以变得可渗透(12)。然而,这种屏障的失败可能是致命的,允许细菌定殖于血液和重要器官。
与屏障功能同等重要的是黏液,它在动物中既古老又广泛存在(52,53)。黏液含有称为黏蛋白的长链糖基化蛋白,它们自组装成动态网状结构和水化的凝胶,其孔径大小和流变性质可通过包括pH、黏蛋白浓度和糖基化在内的因素选择性修改(54,55)。黏液覆盖人体内部的几个关键上皮表面,包括胃肠道(54,55)。其他动物,包括两栖动物(56)和许多海洋无脊椎动物(16,53),也有外部覆盖的黏液或类似黏液的凝胶,其中定殖着微生物组并使气体交换成为可能。昆虫产生围食膜凝胶,其中包含黏蛋白,以包覆肠道中的食物颗粒(57),而植物已趋同进化出从根部释放黏胶,这是一种再次主要由糖蛋白组成的类似黏液的凝胶(58)。这些凝胶不仅充当选择性屏障和润滑剂,还作为微生物的食物来源和附着位点,宿主可以利用这些来塑造微生物组的组成(55,59-64)。
生理学和稳态
宿主对其微生物组影响的关键之一是定义共生体可以栖息的生态位(方框1)。这些生态位在时间和空间上可能有所不同,并在宿主的不同部分选择不同的微生物组(64)。宿主塑造生态位的方式也是多样的,包括提供营养物如黏蛋白(正如刚才讨论的),以及其他产品。例如,人类的阴道微生物组在哺乳动物中是不寻常的,因为它由维持低保护性pH值的乳酸杆菌主导,这可能是通过宿主提供糖原来实现的(65)。同样,哺乳动物肠道中产生的胆汁酸可以强烈影响微生物的代谢,以阻止感染(66)。
在某些微生物组中,氧气控制是至关重要的,它促进复杂碳水化合物和其他底物的发酵(28)。在包括昆虫和脊椎动物在内的不同动物群体中都发现了厌氧肠道条件(67, 68)。通常人们认为,肠道缺氧是由兼性厌氧微生物消耗氧气驱动的。然而,有证据表明,通常是宿主而不是微生物从胃肠道中移除了大部分氧气(13)。最值得注意的是,与拥有微生物组的动物相比,无菌小鼠和无菌蟑螂在整个肠道中的氧气分压同样很低(67, 68),无菌蜜蜂肠道中的氧气张力也非常低(69)。还有证据表明,沿着胃肠道降低氧气水平的活跃机制存在(70)。哺乳动物小肠使用绒毛中的逆流机制帮助从血液中清除氧气,确保到达大肠的内容物已经氧气含量很低。在健康的大肠中,适应低氧环境的上皮细胞快速消耗氧气,进一步降低了氧气水平。这一过程创造了促进微生物发酵的条件(13, 71),并限制了有害的兼性厌氧细菌和其他负面健康结果(28, 71)。蟑螂肠道上皮细胞似乎也采用了类似的氧气控制机制(67)。
肠道形态的总体进化在生理控制中也扮演着重要角色,对于依赖微生物发酵获取能量的宿主物种尤为重要(图3)。尽管人类可能只从微生物发酵中获得总能量的一小部分,但一些食草动物,如马和白蚁,依赖其共生体满足大部分能量需求(72, 73)。与此一致的是,许多反刍动物,包括牛,有一个四室胃,用于培养厌氧菌并吸收大量的微生物发酵产物,然后将相同的微生物传递到后肠作为蛋白质来源进行消化(72)。一个值得注意的例外是大熊猫,其近期进化为食草动物,但仍然拥有与其食肉祖先相似的肠道。因此,大熊猫似乎缺乏促进植物物质有效微生物发酵的能力(74)(图3);结果是极其低效的消化(75)。
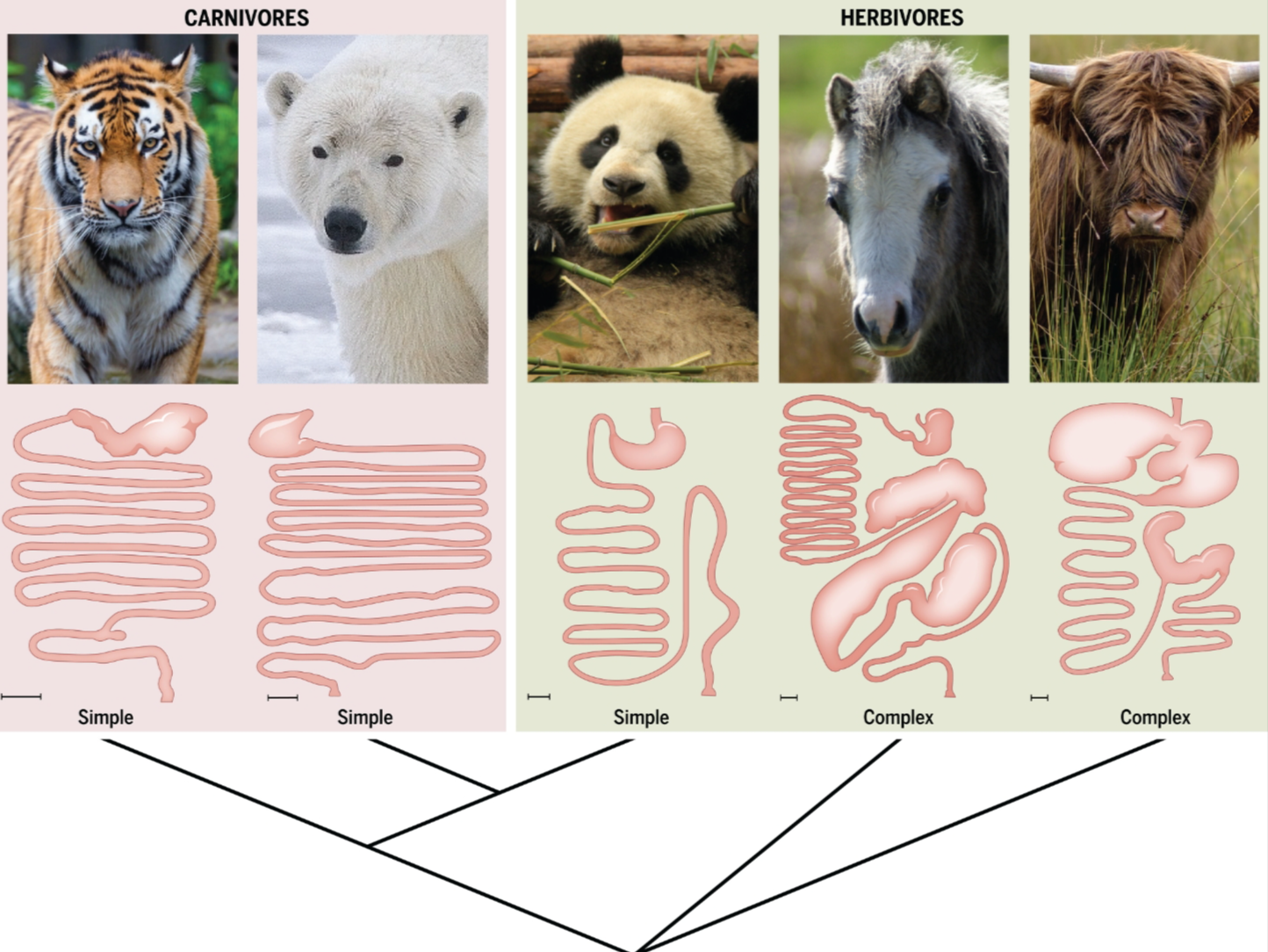
图3. 大熊猫作为宿主控制不佳的一个例子。
多样的哺乳动物食草者进化出了复杂且厌氧的肠道,这使得植物材料的发酵成为可能(例如马和牛)(149)。相比之下,如北极熊这样的食肉动物拥有更简单的肠道形态,较不利于促进微生物发酵(150, 151)。作为一个最近才演化为严格食草者的大熊猫,似乎也缺乏促进纤维素有效微生物发酵的肠道形态和生理功能,其消化效率极低。
转运
许多动物宿主也能够影响共生体的位置。黏液的产生创造了一种从上皮表面远离的持续流动,这在免疫激活下可以增加以清除有问题的微生物(63)。纤毛的动作将微生物和颗粒沿着珊瑚表面、哺乳动物肺部以及肺鱼肠道等不同环境中的上皮表面移动(53)。尽管纤毛在层流下有效地推动一层黏液,但对于大量流体的整体运输来说,它们是不够的,正如大多数脊椎动物肠道中发生的那样。相反,平滑肌使得蠕动的强大和有调节的收缩成为可能。肠道收缩极其复杂,从几乎静止的混合收缩到强烈的蠕动波,这些波推动物质沿肠道移动,最终排出体外(76)。人类肠道中的大部分液体来自分泌物(与食物和饮料相比,分泌了6到7升,而食物和饮料只有2到3升)(77),这意味着分泌和再吸收的相对速率对转运也是至关重要的。
肠道转运的全部潜力在动物腹泻反应中最为明显,其中改变的水重吸收和增加的肠道运动性可以排出大部分肠道微生物(78)。这种反应有助于清除病原体(12),但它也会大幅减少微生物群及其代谢产出(2)。即使在正常情况下,肠道转运也可以降低微生物密度,这在小肠中很明显,那里的转运速度快,共生体密度低(64)。在大肠中,密度要高得多,那里有一系列机制限制共生体的损失,包括湍流混合(2)和规律的反蠕动收缩(76),可以将微生物向相反方向送回胃部。损失进一步通过盲端结构如阑尾和盲肠以及一些物种的结肠生理学来限制,这些结构作用是将富含微生物的颗粒与食物废物分离并保留(79)。尽管有这些措施,蠕动仍可为共生体创造不利环境,共生体必须附着、游动或分裂以补偿其影响(2, 14)。由蠕动引起的高周转意味着组成可以迅速变化,但这种动态环境也为宿主塑造其微生物组创造了机会。
宿主行为
行为选择使宿主能够偏好有益共生体的移入而非有害共生体。例如,避免摄入变质食物可能减少摄入病原体的可能性(80)。包括哺乳动物在内的一些物种还可以迅速学习味道与随后的疾病之间的新关联,以避免此后的危险食物(81),最近的研究表明免疫系统在这种学习中起作用(82)。除了避免有害共生体之外,宿主还表现出与摄入有益共生体相关的行为。例如,对酸味的偏好可能帮助灵长类、大鼠和野猪摄入乳酸菌(83)。此外,许多小型哺乳动物,包括啮齿动物和兔形目动物,产生并食用富含微生物的粪便,这有助于它们满足营养需求并稳定其微生物组(84)。行为还使得特定共生体在亲属间垂直传播成为可能,如蜜蜂狼(85)、白蚁(86)和考拉,考拉母亲会给她的小宝宝喂食牛奶和一种称为pap的特殊形式的粪便(87)。社会昆虫展现出许多额外的行为,有助于调节它们的集体微生物组,包括木蚁使用树脂限制病原体(88)和切叶蚁巡逻真菌园以移除寄生真菌(89)。
宿主控制如何影响微生物组?
因此,宿主已经进化出一套多样化且复杂的机制,允许它们控制和塑造其微生物组。从进化的角度来看,宿主控制的目的是为了增加宿主从其微生物组中获得的适应性益处(图4和框1)。但是,宿主控制机制是如何实现这一目标的呢?宿主控制可以通过改变存在的共生体表型(“伙伴操纵”),或两者兼而有之(10)(图4),来增加宿主的收益。在这里,我们讨论宿主控制可以对共生体产生的四个关键效应(图4),这些效应大致可以分为伙伴选择(对共生体移入或组成的效应)或伙伴操纵(对共生体行为或相互作用的效应)。
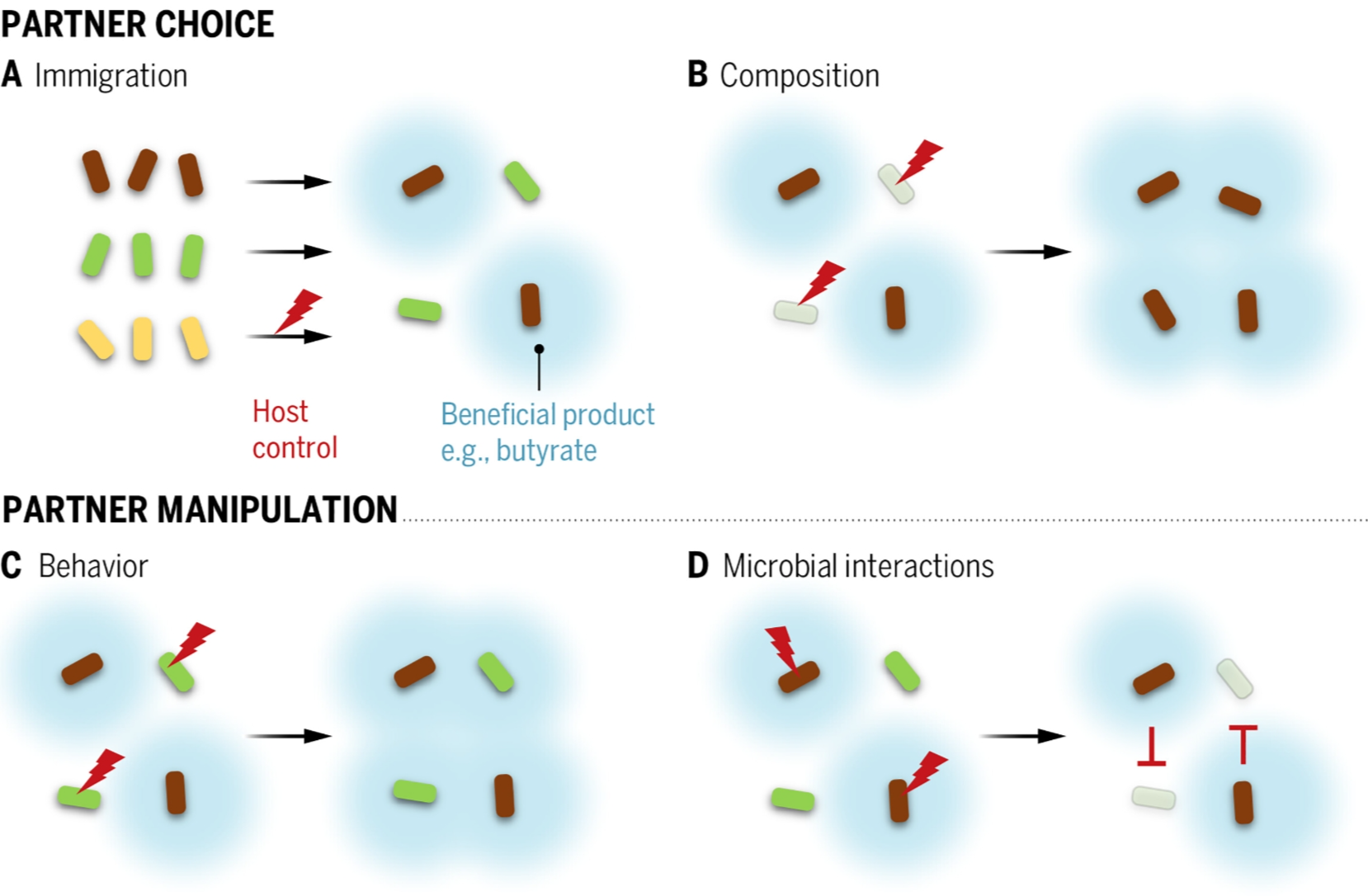
图4. 宿主控制如何影响微生物组?
宿主控制(红色闪电箭头)可以通过四种方式增加从共生体获得的利益。在每种情况下,棕色细菌株是产生有益产物的受青睐株,如细菌周围的蓝色光晕所示,例如短链脂肪酸如丁酸。(A)移民控制:宿主限制了一些不产生有益产物的菌株(黄色)的定植。(B)组成控制:宿主选择不利于已建立的不产生该产物的菌株(绿色)。(C)行为控制:宿主通过提供前体物质改变绿色菌株的代谢,使其现在产生有益产物(蓝色光晕)。(D)控制微生物相互作用:宿主促进共生体之间的竞争(红色抑制符号),有益的菌株(棕色)消除了绿色菌株。前两个案例是伙伴选择的例子,因为它们通过促进有益共生体的丰度超过非有益共生体而起作用。底部的案例是伙伴操纵的例子,它们通过改变共生体的行为起作用,这些行为的变化也可能导致随后的丰度变化(10)。
共生体移民
对宿主来说,一个重要目标是影响哪些物种从其他宿主和更广泛环境中存在的可传播微生物中到达并在其微生物组中建立(9, 90)(图4A)。这种伙伴选择的形式包括在宿主发育期间调节微生物组组装的过程(91)。当一些共生体相互依赖以获取营养时,组装可能会遇到障碍,因为如果没有其他物种的存在,某些物种可能无法定植(91)。宿主的一个解决方案是通过亲属间的传递确保所需的共生体组合一起到达(86, 87),尽管理论预测,为了避免完美的垂直传递,也存在进化激励,以获得适合当前环境的有益共生体组合(92)。另一种塑造微生物组组装的方式是为共生体提供特定营养素:人类乳汁含有婴儿无法代谢但某些定植细菌可以代谢的多样化寡糖和尿素(93, 94)。此外,乳汁中含有免疫球蛋白,它们通过为特定菌株提供附着位点来塑造哪些微生物定植于婴儿黏膜上(95)。
不同脊椎动物中发现的酸性胃限制了进入肠道的总移民率,一些物种,包括秃鹫和人类,具有极低的肠道pH值,这可能与高病原体暴露有关(96)。宿主还使用生理控制从移民的菌株和物种中选择高质量的共生体。例如,许多动物的无氧肠道有助于筛选能够进行发酵的微生物。在短尾鱿鱼光器官中,筛选通过生成具有挑战性的生长条件进行,包括活性氧和氮物种(97, 98),而生长真菌的蜜蜂则在宿主树木内偏好乙醇丰富的条件,以选择它们的真菌园共生体而不是其他微生物(17)。另一种控制移民的方式是对待常驻共生体与新来者不同。埃及伊蚊从其肠上皮释放抗菌肽。然而,它还释放C型凝集素,这些凝集素包裹常驻共生体,保护它们免受抗菌剂的影响(99)。
宿主与已建立的共生体之间的相互作用
除了移民控制,宿主还可以通过影响已建立微生物的数量来获益(图4B)。可以限制问题共生体的资源配置或为有益共生体提供资源。在哺乳动物的微生物组中,炎症期间释放的脂质运载蛋白-2与某些细菌的铁螯合剂结合,限制它们的铁摄取(100)。豆科植物将固氮细菌容纳在特化的根瘤中,但会切断供应过少硝酸钾的根瘤的资源(5)(图2)。此外,从植物根部释放的大量碳水化合物和粘液可以喂养根际微生物组(58)。在某些情况下,这种粘液围绕着气生根,并似乎选择抑制环境细菌而非固氮细菌生长的真菌伙伴(101)。粘液的产生具有类似潜力,通过优先喂养能够分解粘液中复杂碳水化合物的微生物来影响动物微生物组,例如拟杆菌科和阿克曼氏菌属的成员(62)。类似地,蜜蜂分泌有机酸以支持核心共生体如Snodgrassella alvi的定殖,这些共生体无法代谢饮食中的糖分(102)。理论工作表明,营养供给可能是一种特别强大的宿主控制模式,因为被喂养的菌株有潜力繁荣并排除其他微生物从上皮表面(60)。
宿主还可以通过调节与宿主组织的粘附以及微生物间的粘附来改变微生物组成(59)。例如,适应性免疫对肠道微生物组的主要影响是通过粘附介导的。IgA通过促进细菌-细菌聚集来促进细菌的排出,这形成了可以通过蠕动移除的团块(15, 39);但是,正如所讨论的,IgA可能反而通过促进外层粘液中的细菌-粘液相互作用来协助定殖(39, 41)。当细菌以粘液为食时,也会附着到粘液上,使用糖结合蛋白(62)。一个有趣的可能是,结合蠕动对未附着微生物的负面影响,这种粘附将富集具有粘液代谢生化机制的共生体。我们对哺乳动物微生物组内部精细空间结构的理解仍然有限(64)。然而,远端肠道的分层粘液层,包括糖萼和内外粘液层,有可能生成多个生态位,微生物可以附着和生长(64)。此外,哺乳动物中粘膜和肠道腔内的微生物组成可能不同(103),并且有证据表明,一些细菌物种依靠粘液觅食(61, 62)或IgA附着(41)才能在肠道中竞争。潜在的病原体如大肠杆菌也可以在粘膜中生存,其移动性、耐氧性和利用能力以及在宿主脂质上觅食的能力使其能够存活(103),这可能反映了宿主控制的一些限制。尽管如此,如果宿主能够选择一个以粘液相关共生体为主的群落,而不是那些不与粘液相互作用的共生体,由于粘液粘附似乎使细菌不太可能穿透上皮层,因此致病风险可能会降低(104)。此外,尽管粘附到密集粘液可能增加靠近上皮层的微生物密度,但理论工作预测,宿主可以通过增加粘液产生速率来保留从粘膜上剥离这些粘性共生体的能力(59)。
共生体的代谢和行为
除了调节微生物组的组成,宿主可以直接影响常驻共生体的行为,以增加它从中获得的好处(图4C)。在实践中,影响微生物行为的机制也可能改变丰度和组成(图4B)。例如,在细菌感染后,小鼠增加了胆汁酸牛磺酸的产生,这不仅将微生物组的组成向能够进行牛磺酸代谢的微生物物种转变(伙伴选择),还促进了那些能够进行该代谢的微生物的牛磺酸代谢(伙伴操纵)(图4)。结果是硫化物产量增加,这抑制了呼吸并限制了依赖呼吸的病原体未来感染的潜力(66)。然而,在某些情况下,宿主能够影响单个共生体的代谢,而对整体微生物组成的影响有限。例如,豆科植物合成豆血红蛋白(105),这是动物血红蛋白的类似物,能在纳摩尔范围内结合和缓冲游离氧。这种缓冲使得植物的共生细菌能够进行固氮作用,这些细菌依赖于对氧敏感的固氮酶,同时仍保持足够的氧气流量以支持细菌呼吸所需的能量需求。
氧气控制对于许多动物物种也很重要。呼吸允许微生物通过电子转移到氧气或替代接受器如NO3−来最大化从营养物质中提取的能量(三磷酸腺苷)。然而,结果,呼吸可以大大减少宿主可用的能量,因为共生体可以完全氧化富含能量的化合物为CO2和H2O。因此,通过限制氧气供应,宿主可以迫使微生物使用较弱的电子接受器,如丙酮酸,并产生发酵产品,如丁酸和其他短链脂肪酸。这些产品保留了代谢潜能,可以被自由接触氧气的宿主组织进一步代谢(28)。在人类结肠中,肠上皮细胞消耗这些产品,从而也消耗氧气并生成正反馈循环以进一步促进微生物发酵。这个循环不断受到肠道上皮细胞的监测,因为如丁酸等发酵产品的减少可能会破坏低氧诱导因子,这是一种协调屏障保护的转录因子(13, 29)。
共生体的行为也受到限制微生物侵入宿主组织的屏障的控制。小鼠的粘液产生既保护上皮表面,又有助于粪便材料的流变特性,这帮助将微生物群与宿主分离(106)。宿主可以通过影响共生体的运动和附着来进一步加强屏障。游泳细菌构成威胁,因为它们可以抵抗蠕动或纤毛介导的转运(107),并突破上皮屏障(4, 108)。作为回应,动物和植物中都进化出了多样的抗鞭毛机制(4, 109)。哺乳动物可以从肠道上皮分泌一种名为含Ly6/PLAUR结构域8(LYPD8)的蛋白质,该蛋白与细菌鞭毛结合并抑制其游动(110)。另一种宿主策略是产生诱饵分子,提供替代的结合位点,从而限制对上皮的附着(111)。一些抗微生物肽还能改变共生体的行为:哺乳动物抗微生物肽Y在小鼠肠道中抑制真菌念珠菌的菌丝(和侵入性)形态。在存在这种肽的情况下,真菌保持其酵母形态,不附着和侵入宿主组织(112)。
共生体的相互作用
微生物组内的共生体可以强烈地相互影响生长和存活(3, 9)。宿主可以从塑造这些相互作用中获益——这是一种生态工程的形式(图4D)。微生物间的生态互动可以是负面的或正面的,对于微生物组成很重要,因为竞争排斥可以消除物种,而交叉喂养可以创造新的生态位。生态互动还影响微生物组的生产力(91, 113)和对病原体入侵的抵抗力(114),以及微生物群落如何响应如抗生素治疗这样的扰动(生态稳定性)。例如,共生体之间的强合作互动在它们造成相互依赖时可能会破坏稳定,即一个物种的数量下降导致另一个物种的数量下降。强烈的竞争互动也可能破坏微生物组的稳定性,并在降低共生体生长速率时导致灭绝(113)。因此,限制共生体间互动的强度有可能通过促进稳定性和共存而使宿主受益。
宿主调节共生体互动的一种策略是将有益的共生体安置在专用器官或组织中,这可以限制与较少益处但生长速度更快的微生物的竞争。这种结构广泛存在于昆虫和海洋无脊椎动物中以容纳内共生细菌(85, 97, 98)以及植物中的固氮细菌(5)。在哺乳动物中,通过隐窝(115)或盲端结构如盲肠引入的空间结构也可能有助于限制共生体之间的竞争(113)。从上皮表面喂食共生体(113),例如通过提供粘液,也可能减少竞争排斥并生成新的生态位(62, 116)。另一个有趣的可能是宿主可以操纵微生物组中细菌的细胞间信号传递以促进合作表型。与此可能性一致的是,哺乳动物细胞制造了一种广泛使用的群体感应自诱导剂AI-2的模拟物(117),刺胞动物水螅修改了其关键共生体的群体感应信号,这促进了定殖(118)。
在某些情况下,宿主似乎促进物种间的相互作用。粘液提供了噬菌体附着等待细菌的位置,这被认为通过促进细菌和噬菌体之间的寄生互动改善了屏障功能(119)。宿主产生的蠕动也可以促进互动强度,因为增加流速和混合收缩将倾向于使不同菌株和物种的种群均质化,从而增加不同共生体相遇和互动的可能性(2, 64)。当物种通过代谢相互作用为宿主提供营养时,这种混合可能带来好处,正如我们的大肠中发生的一样,其中乙酸生产者养活丁酸生产者,丁酸生产者养活我们,以及在反刍动物中发生的情况(120)(图3)。而且,当这种代谢互动基于剥削时,一个物种以另一个物种的损失为代价获益,这产生了一个可以为微生物组稳定的负反馈循环(113)。然而,肠道传输也有潜力限制多样性和稳定性:发现斑马鱼中的两种群落由于蠕动运动而被破坏稳定,因为它促进了一种共生体通过竞争排斥另一种共生体(14)。
如果竞争能选择出为宿主提供益处的旺盛生长的细菌,比如能够超越并使入侵的病原体中毒的能力,那么促进竞争可能在进化时间尺度上带来好处(66, 114)。然而,共生体的进化可能是一把双刃剑,如果它使得共生体能够从宿主控制中逃脱,正如我们接下来讨论的。
共生体的进化与反适应
相对于宿主,共生体的代时通常非常短。在生态学上,这导致了微生物群落物种组成的快速变化,这种变化是由共生体的死亡和生长速率差异驱动的。短的代时也意味着共生体物种可以在宿主的生命周期内迅速进化并在其种群内经历遗传变化(9, 40, 121, 122)。当微生物首次被引入宿主并经历由新环境驱动的自然选择时,进化适应的可能性最为明显(123)。共生体经历的进化压力也可能在宿主的一生中发生变化,正如在人类发展厌氧肠道期间可能发生的那样(124)。对于在宿主中建立的共生体,自然选择可能只是偶尔成为改变的力量。通常,自然选择会反对变化,剔除偏离宿主偏好表型的变异(即净化选择)(9)。尽管如此,成熟的宿主中也持续看到自然选择的迹象。例如,对已建立的小鼠和人类微生物组的研究发现,与碳水化合物利用(121, 125)和IgA亲和力(121)相关的细菌基因存在正向选择的证据。宿主控制机制还可能通过促进共生体专业化和与宿主共同多样化,影响更长时间尺度上的进化过程(126)。
如果有效利用,共生体的进化可能对宿主有益。新定植于人类微生物组的物种可能会经历自然选择,变得非运动性(4)、与粘液相互作用(121)以及产生丁酸(13)。这意味着曾经是病原性的菌株可能进化成中性(127),而曾经是中性的菌株可能进化出实质性的好处(4)。与这些压力一致——以及宿主用来对抗游动细菌的多样机制——最近的一项系统发育分析表明,作为动物共生体的细菌比自由生活的物种具有更高的鞭毛丢失进化率(4)。
尽管对宿主有潜在的好处,共生体的进化也带来了逃离宿主控制的潜在可能性。因此,进化理论预测,只有当控制机制能够限制共生体的反向进化时,才会被保持(4)。与这一预测一致的是,控制机制通常针对微生物的表型(即基于特征的歧视),而不是特定的菌株或物种(即基于基因型的歧视)(3)。基于特征的歧视对宿主很重要,因为如果某个共生体进化并改变了其表型,它们将受到不同的对待。在我们讨论的大多数例子中都看到了这种基于特征的控制。例如,在短尾鱿鱼中,共生体Aliivibrio fischeri失去发光能力后,在鱿鱼的光器官中变得没有竞争力(97, 98)。基于特征的歧视的一个显著例外是适应性免疫(39),它通过独特的化学签名识别微生物的基因型。然而,由于能够学习新的关联,适应性免疫也能够修改响应,以防原本有益的共生体进化成有害状态。
为了进一步限制反向进化,宿主控制可以针对微生物的进化受限特征。例如,如果控制机制需要微生物获得多个代谢位点,如黏蛋白代谢所需的那些,那么喂养共生体的控制机制可能很难被利用(62)。惩罚性宿主控制机制也似乎针对了微生物的受限特征,以防止进化逃逸。通过TLR5识别鞭毛素促进炎症和产生限制游泳的抗鞭毛素IgA(128, 129)。因此,避免这种反应可能为一些细菌物种带来相当多的好处。然而,TLR5绑定到细菌鞭毛素的一个高度保守区域,氨基酸的变化通常会导致游泳缺陷,这限制了对抗适应的范围(4, 130)。
尽管有这些宿主适应,一些共生体仍进化出规避宿主控制机制的方法。这些反策略很重要,因为它们为宿主控制对共生体进化的影响提供了令人信服的证据。预计这些反策略也会广泛存在,但它们的识别通常需要详细了解宿主和微生物生物学。因此,许多潜在的实例来自于研究充分的病原体。例如,Shigella flexneri抑制了原本会将受感染上皮细胞排入肠道腔内的炎症小体反应(131),而Clostridium difficile(现为Clostridioides difficile)既抑制肠道蠕动(132),又通过拥有储存铁的铁蛋白器官来抵抗宿主的铁螯合(133)。还有证据表明非病原性共生体对宿主控制的进化响应。一个例子是最近发现一些肠道细菌携带一种形式的鞭毛素,它仍然能与TLR5结合但沉默其信号传导(134)。多样的共生体似乎也进化出了对宿主抗微生物肽(AMPs)的抗性。这种抗性并不是对宿主的问题,而是增强了微生物组抵御仍易受AMPs影响的入侵病原体的能力(30)。因此,共生体对控制的反适应可能并不总是问题。然而,阻止有害物种采取相同的反适应措施的原因并不总是清楚的。
一些微生物不仅逃避宿主控制,还利用本应控制它们的系统。植物病原体Agrobacterium tumefaciens利用关键的宿主免疫蛋白将其自身的DNA输送到植物细胞核中,并控制宿主生理以自利(135)。另一个引人入胜的例子是肠道病原体Salmonella enterica对宿主炎症反应的颠覆(28)。这种细菌进入肠道上皮细胞并释放引起大规模炎症反应的免疫原性分子。尽管这种反应将S. enterica与常驻肠道微生物群一起击倒,但该病原体能够迅速恢复并由于其能够使用由免疫反应产生的副产品作为电子受体进行呼吸,从而超过常驻物种生长。这样,S. enterica设法劫持最复杂的宿主控制机制来增殖和传播。
展望
为了更好的健康操纵微生物组是具有挑战性的。抗生素可以帮助处理一些疾病,但这是一种粗糙的操纵方式,它会摧毁微生物组并导致药物抗性的进化。因此,人们非常感兴趣于潜在的替代方法,包括噬菌体治疗、粪便微生物移植和特定微生物联合体的递送(114, 136, 137)。然而,像抗生素一样,这些策略都集中在微生物本身。一个替代方案是关注我们在这里讨论过的宿主控制机制。这些机制经过自然选择塑造,以应对微生物组内在的多样性和可变性,甚至限制抗药性的进化。
应用包括诊断作用——其中宿主生理和控制的标志物在条件和个体之间可能比微生物组成更相似(28)(方框2)——但也用于预防和治疗疾病。在这些策略中,核心是疫苗接种,近期工作展示了疫苗针对肠道微生物组中特定细菌的潜力(138, 139)。另一种策略是通过恢复肠道的厌氧环境(13, 28)或调节关键营养素如铁(100)、硝酸钾(140)或黏蛋白衍生的聚糖(116, 141)来增强宿主对共生体代谢的控制。更一般地说,任何促进健康黏膜层和宿主上皮屏障完整性的策略都具有明显的健康益处。随着我们年龄的增长,微生物组变得更加多变和易患疾病(64, 142),这可能是宿主控制机制衰退的一个症状,这样的治疗可能会变得尤为重要。
人们对微生物组(包括人类)影响宿主的多种方式非常感兴趣。在这里,我们认为这些效应中的许多都可以追溯到宿主控制的机制。数亿年来,多细胞宿主一直在自然选择下增加来自其共生体的益处,结果是一系列惊人的适应性,能够调节和控制多样的微生物群落。研究这些机制对于理解微生物组和为了更好的健康操纵它们是至关重要的。
https://blog.sciencenet.cn/blog-41174-1443359.html
上一篇:GLP-1的益处远不是减肥
下一篇:发现令科学家困惑海底“暗氧”