博文
土壤高亲和力氢氧化细菌的生态特征
||
高亲和力氢氧化土壤细菌的生态特征
1. 土壤中的大气氢循环涉及氢氧化细菌。
2. 高亲和力氢氧化基因在系统发育上可能不具有保守性。
3. 本文提出了一种鉴定含高亲和力氢氧化(hhy)基因细菌生态特征的方法。
4. 生态特征可解释氢氧化细菌对土壤因子的响应机制。
摘要
每年,土壤微生物介导的氢气(H₂)氧化会清除全球约80%的大气氢气。土壤中的高亲和力氢氧化细菌可将这种微量气体作为能量来源,在其他底物有限时维持生存,或在休眠期间满足自身维持能量需求。然而,目前对这类细菌的分布、组成、多样性及功能,尤其是其生态特征,即影响其与环境及其他生物相互作用的特性的了解仍较为有限。这是因为高亲和力氢氧化细菌在系统发育上不具有保守性,其原因可能是其功能基因的水平转移。这使得以下生态问题难以解答:氢氧化细菌在土壤氢循环中的分布、功能作用及生态贡献,以及它们对环境因子的响应机制。而这些信息对于估算氢氧化细菌对全球氢循环的贡献至关重要。尽管许多氢氧化细菌无法培养,但它们在应对环境变化(如pH值、含水量、氢气浓度)时,可能具有相似的生态特征。因此,基于群落或功能群水平的特征分析方法,或可用于整合复杂的基因组和系统发育信息。本综述探讨了土壤环境因子对土壤氢气吸收(通过氧化作用)的影响,并确定了受控条件下的生态响应特征。我们的研究方法将氢氧化细菌的生物活性与其生存环境联系起来,旨在进一步扩大研究尺度,并估算土壤微生物群落缓解与大气氢气增加相关的全球变暖的能力。
Ecological traits of high-affinity hydrogen-oxidizing soil bacteria involved in the hydrogen cycle
1. 全球氢气平衡
1.1 全球氢气平衡的当前估算
分子氢(H₂)是大气中含量仅次于甲烷(CH₄)的可氧化气体。全球氢气储量估计为155太克(Tg)(Novelli等,1999;Ehhalt和Rohrer,2009),其中约60太克/年(Tg H₂ yr⁻1)的氢气会从大气循环至岩石圈。这使得氢气的大气停留时间约为2年,大气浓度约为0.53体积百万分比(ppmv)。在过去150年间,大气氢气浓度已从估计的0.3 ppmv上升至约0.53 ppmv(Patterson等,2021)。这一变化归因于人类活动(如化肥生产和氢能产业)——如今这些活动的氢气排放量可能高于以往。
尽管大气氢气混合比在长期内会波动(主要由人为排放及其他来源驱动),但土壤仍是大气氢气的主要汇。大气氢气向土壤转移并在土壤中被氧化为水,这一过程有助于维持氢气产生与消耗之间的动态平衡(图1)。尽管存在这种平衡,氢气作为间接温室气体的问题仍令人担忧:它会与羟基自由基(OH)发生反应,从而延长强效温室气体甲烷(CH₄)的大气停留时间。在100年时间尺度上,氢气本身的全球变暖潜能值为5±1;若未来不对氢能来源的泄漏进行严格控制,其对全球变暖的影响预计将十分显著(Derwent等,2020)。

图1 全球氢气循环**(改编自Constant等,2009;Ehhalt和F. Rohrer,2009)
大气氢气主要通过非生物和生物途径中的光化学氧化产生。约90%的大气氢气来源于非生物反应,如甲烷(CH₄)及其他非甲烷简单烃的光化学氧化。此外,烃类燃烧(即化石燃料和生物质燃烧)也会向大气释放氢气。工业用氢的逸散排放是大气氢气的另一来源。约10%的大气氢气通过生物过程释放,包括生物固氮(N₂)以及光发酵/暗发酵(Klein等,2020)。
大气氢气浓度由生物汇和非生物汇共同调控。氢气的生物汇涉及微生物氧化作用,由土壤原核生物完成——这些生物每年可消耗约80%的大气氢气(Greening等,2015)。氢气的非生物汇涉及羟基自由基(OH),其每年可从大气中清除约20%的氢气(Constant等,2009)。在全球范围内,北半球的大气氢气浓度低于南半球,这是因为北半球陆地面积更大,对氢气的吸收能力更强(Schmidt和Wetter,2003)。
1.2 大气氢气产生与消耗的关键过程
1.2.1 大气氢气的非生物产生
甲烷(CH₄)及其他简单烃(如异戊二烯、萜烯)的光化学氧化,通过式(1)中的连续反应,产生约50%的大气氢气(Ravishankara,1988;Schmidt和Wetter,2003;Constant等,2009)。
(1)OH + CH₄ → CH₃ + O₂ → CH₃OOH + OH → CH₂OOH + H₂O → CHOH → H₂ + CO
通过对实验性森林火灾周边气团的分析发现,全球生物质燃烧贡献了约20%的大气氢气(15 Tg H₂ yr⁻1)(Constant等,2009)。分子氢(H₂)通过干废物残渣的降解及复杂固体烃的加热分解产生。例如,由于化石燃料燃烧,在早晚交通高峰期,大气氢气浓度会升高(Vollmer等,2007)。
化学和石油化工行业对分子氢(H₂)的工业利用,是氢气逸散排放的来源之一。这些行业依赖天然气制得的高浓度氢气生产氨肥和石油精炼副产品,并应用于冶金、电子和制药领域的工艺中(Ramachandran和Menon,1998;Olah等,2005)。根据氢气产量与一氧化碳(CO)排放清单的相关性及H₂/CO排放比估算,约15%~20%的大气氢气来源于这些人为源(Hauglustaine和Ehhalt,2002)。此外,辐射分解、机械自由基反应、火山脱气和热液蚀变等地质过程,对大气氢气库的年贡献量不足总氢气产量的0.01%(Klein等,2020)。
1.2.2 大气氢气的生物产生
生物固氮(N₂)是目前已知最主要的氢气生物产生途径,约占全球氢气生成量的4%。该反应由细菌催化,这些细菌可能是植物共生体(如在豆科植物中形成根瘤),也可能是定殖于根、茎或叶组织中的植物内生固氮菌。在豆科植物根瘤中,转移至固氮酶(负责生物固氮的酶)的电子中,约30%~40%用于合成氢气(McLearn和Dong,2002;Constant等,2009)。一些编码氢吸收氢化酶(Hup)的共生根瘤菌,可在根瘤内循环利用产生的氢气。然而,自然界中大多数根瘤菌-豆科植物共生体(尤其是农业生产中针对性接种豆科植物所形成的共生体)缺乏Hup。例如,美国种植的商业大豆中,约75%接种的根瘤菌不具备Hup活性;而许多高效固氮组合(如苜蓿根瘤)中则含有含Hup的根瘤菌(Uratsu等,1982)。如式(2)所示,每固定1分子N₂会生成1分子H₂,生成的H₂要么通过Hup循环利用,要么从根瘤中释放:
(2)1N₂ + 8H⁺ + 16MgATP + 8e⁻ → 2NH₃ + 1H₂ + 16MgADP + 16Pi
因此,生物固氮每年向全球大气释放的氢气估计为3.5 Tg。该估算值基于氢气年通量,同时考虑了各生物群系中实证性生物固氮的平均速率,以及每个生物群系中可能形成固氮共生体的豆科植物种植面积(Conrad和Seiler,1980;Constant等,2009;Vitousek等,2013)。除豆科植物共生体及与植物相关的固氮菌外,蓝细菌(Cyanobacteria)、假单胞菌(Pseudomonas)、固氮螺菌(Azospirillum)、固氮菌(Azotobacter)等自由生活的固氮菌,也会参与氢气的生物产生。
发酵作用是分子氢(H₂)的另一生物来源(图2),主要发生在缺氧的黑暗环境中,如渍水土壤、蚯蚓和白蚁等土壤动物的消化道。光发酵产氢是另一种微生物过程,发生在厌氧、氮限制条件下:该反应以光为能量来源,通过有机残渣的不完全氧化产生电子和质子,并可能生成分子氢(Androga等,2012)。在无光无氧条件下,微生物介导的暗发酵也可通过有机物的不完全降解生成氢气(Ma和Tao,2023)。然而,这些过程对全球氢气产生的贡献几乎可忽略不计,因为土壤中该氢气产生途径与氢气氧化反应紧密耦合——土壤氢氧化菌通常能在生成的氢气释放到土壤外之前将其循环利用。例如,在豆科植物根际,氢气会保留在植物-土壤系统中,并在其产生地(根瘤)周围1~4厘米范围内被氧化(Annan等,2012;Porte等,2020)。
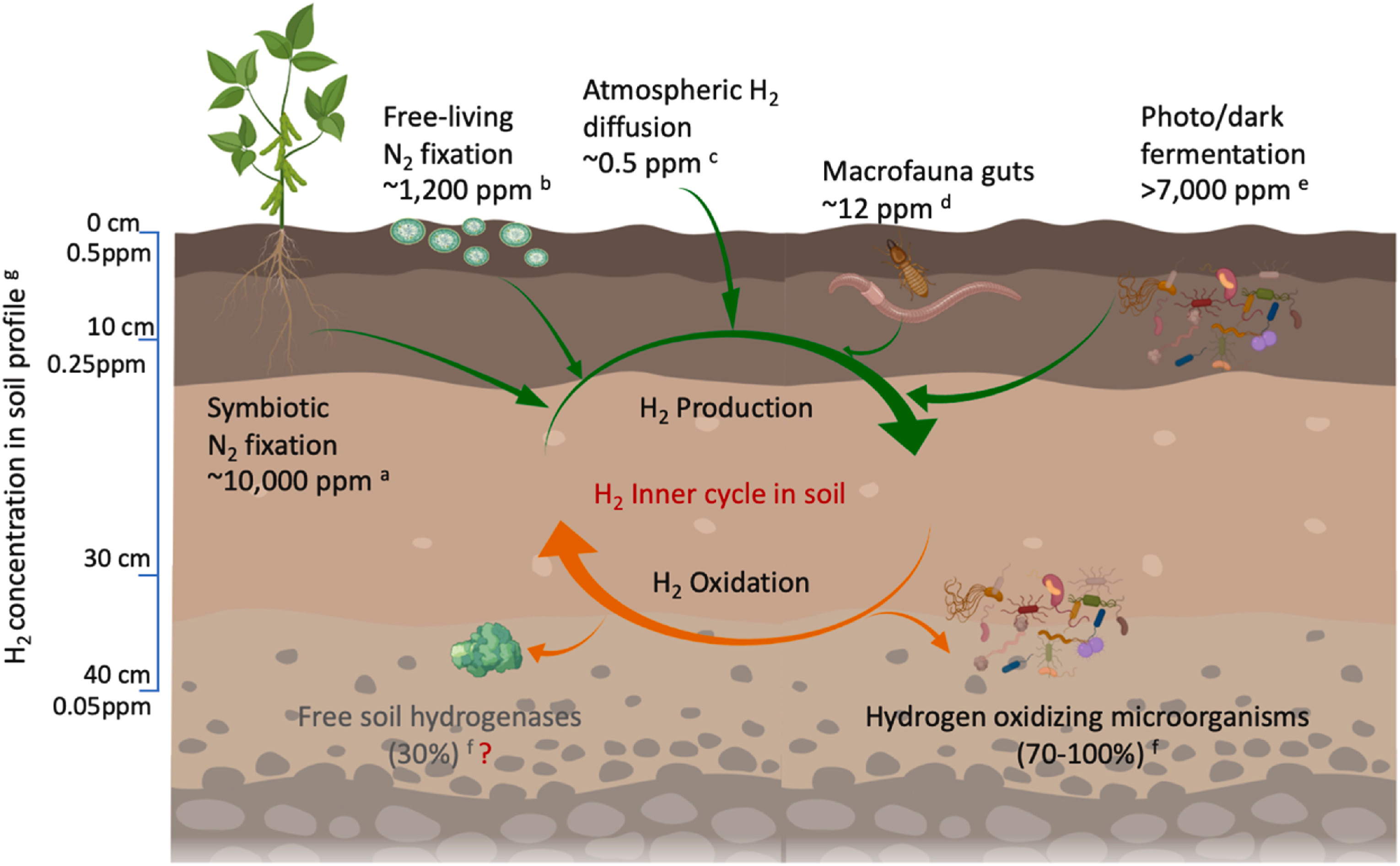
图2 土壤环境中的氢气来源及氢气浓度**
a Porte等(2020);b Nyberg等(2015);c Constant等(2009);d Wust等(2009);e Kayahan等(2017);e Akhlaghi和Najafpour-Darzi(2020);f King(2003);g Smith-Downey等(2008)
在土壤剖面中,根据土壤性质和气体扩散速率的不同,氢气浓度会随土壤深度呈指数下降(Yonemura等,2000a)。Smith-Downey等(2008)开展的田间调查分析了土壤剖面(深度达40厘米)的氢气浓度:在10厘米深度处,氢气浓度仅为大气水平的50%;而在40厘米深度处,浓度急剧下降至大气水平的10%。土壤剖面中的氢气浓度,反映了每个空间位置上氢气来源(大气扩散+生物产生)与氢气氧化之间的平衡关系(图2)。
1.2.3 羟基自由基介导的非生物氧化
大气中的氢气(H₂)通过与羟基自由基(OH)反应发生氧化,主要产生两种结果:一是生成水蒸气;二是在大气中一氧化氮(NO)浓度较低的特定条件下,导致臭氧(O₃)消耗。另一方面,在污染区域,高浓度的一氧化氮(NO)会引发光化学臭氧的形成(Constant等,2009)。一系列涉及氢气(H₂)与羟基自由基(OH)的反应会加剧间接温室效应。尽管氢气(H₂)本身并非强效温室气体,但它参与的关键反应会通过多种途径影响地球气候。以下四种机制可解释氢氧化作用与全球气候变化的关联(图3):
(1)生成的水蒸气作为温室气体,会将热量截留于高层大气中,导致地球升温;
(2)平流层中的臭氧消耗会使太阳释放的有害紫外线辐射穿透大气层到达地球表面,进而加剧地表升温;
(3)在一氧化氮(NO)浓度较高的区域,光化学作用形成的臭氧(O₃)属于温室气体,会将热量截留于对流层上层;此外,它还会促进对流层下层雾霾的形成;
(4)氢气(H₂)与羟基自由基(OH)的反应会降低羟基自由基的可利用性,而羟基自由基对氧化另一种强效温室气体甲烷(CH₄)至关重要。由于大气中约90%的甲烷(CH₄)通过羟基自由基(OH)氧化清除,因此羟基自由基可利用性的降低会延长甲烷在大气中的停留时间,从而加剧温室效应(Stavert等,2022)。
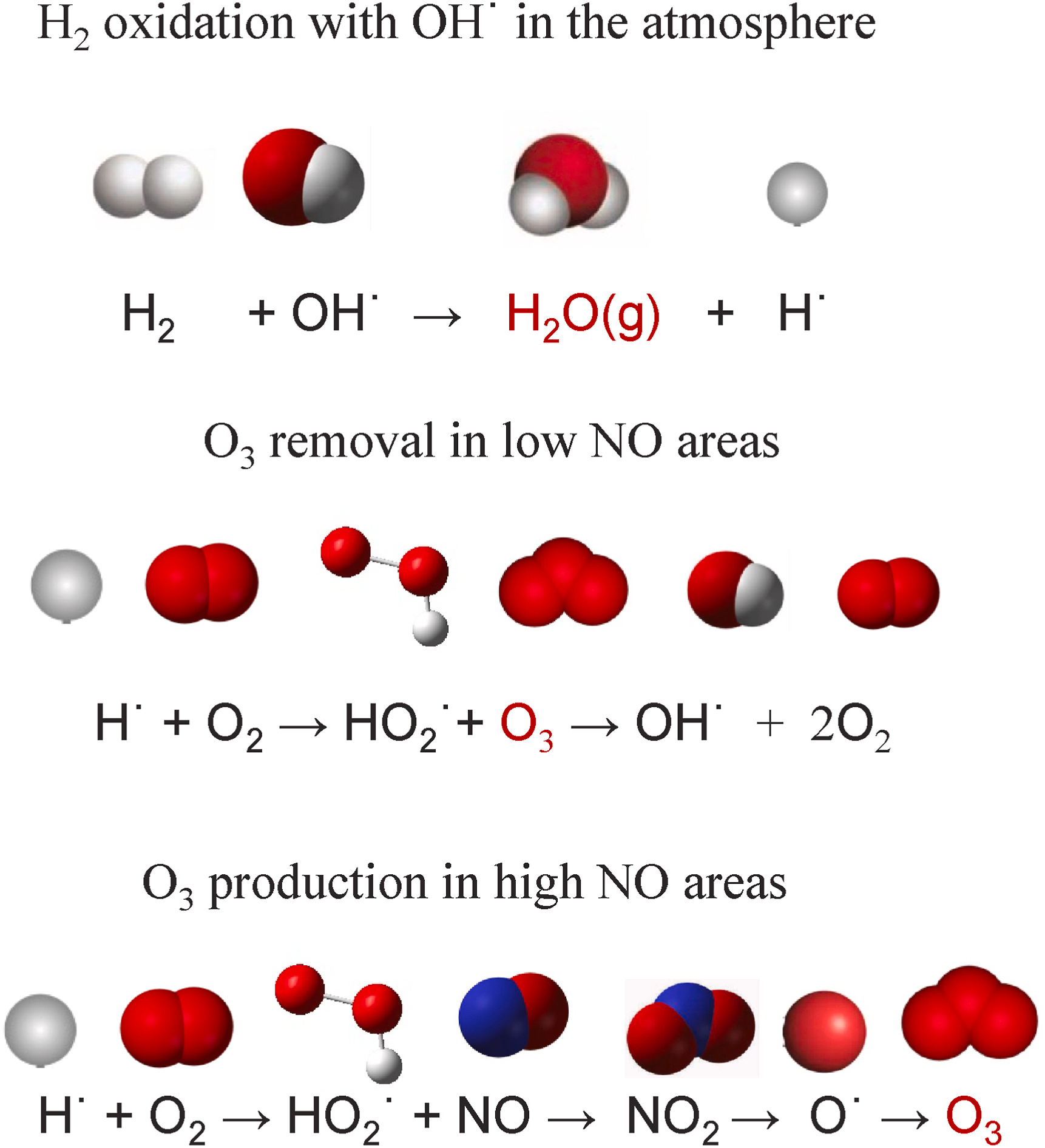
图3 氢气(H₂)介导的大气化学反应示意图,展示氢气如何间接促进温室效应**
综上,尽管氢气(H₂)不会直接导致显著的全球变暖,但其与羟基自由基(OH)的反应能力可通过生成水蒸气和消耗臭氧影响地球气候。此外,氢气(H₂)参与的反应会降低原本用于氧化甲烷(CH₄)的羟基自由基(OH)的可利用性,进而延长甲烷在大气中的停留时间。
1.2.4 土壤对氢气的生物吸收
大气中的氢气(H₂)主要通过微生物介导的土壤吸收过程被消耗,这一过程约占大气氢气清除量的80%(约60-80太克/年,Tg H₂ yr⁻1)。土壤对氢气的吸收是由以氢气为能量来源的微生物控制的生物过程。微生物在生化反应中偏好利用氢气,得益于其优异的理化特性:低活化能(21千焦/摩尔,kJ mol⁻1)、低氧化还原电位(E°’ = -420毫伏,mV)、高扩散系数(4×10⁻⁹平方米/秒,m2 s⁻1)以及高能量密度(反应式:H₂→2H⁺+2e⁻,标准吉布斯自由能变ΔGo’ = -237千焦/摩尔,kJ mol⁻1;Koch等,2014)。
氯仿熏蒸处理的土壤会丧失约80%的氢气吸收能力,这一实验结果证实了微生物在土壤氢气吸收中的核心作用(King,2003)。由于经氯仿熏蒸的土壤在密闭环境中仍能吸收约20%的供给氢气,King(2003)提出“游离氢气吸收酶(来自死亡细胞的裂解产物)”可通过氧化反应参与土壤氢气吸收。然而,后续研究对这一观点提出了质疑:从土壤中提取的无细菌提取物仅表现出不到2%的氢气吸收活性(Guo和Conrad,2008),甚至在部分研究中几乎检测不到氢气吸收活性(Constant等,2011)。因此,我们可得出结论:土壤对氢气的吸收主要由土壤微生物介导。
1.3 填补全球氢气平衡研究中的知识空白
过去150年间,大气氢气(H₂)浓度几乎翻倍,这表明全球氢气的产生量已超过消耗量。这一现象主要归因于更多非生物来源的氢气进入大气,其中氢能产业是主要贡献者之一。然而,目前我们对全球“汇”(即氢气清除途径)降低大气氢气浓度的能力仍缺乏了解。由于大气氢气浓度主要由生物汇调控,因此明确土壤氢氧化微生物如何适应氢气浓度变化、及其活性如何依赖土壤理化性质(如土壤pH值、土壤碳含量、土壤含水量、温度等)至关重要。因此,研究氢氧化微生物的分布、生态学特征及氢气消耗能力,将有助于更准确地估算全球大气氢气平衡。
2 土壤中氢氧化细菌的生理生态学特征
2.1 不同生态系统土壤的氢气吸收活性
陆地生态系统是全球氢气(H₂)的主要汇,每年通过土壤微生物介导的生物氧化(吸收)过程清除约80%的大气氢气。然而,地球陆地生态系统结构复杂,植被、地形、地质和水文条件具有高度多样性。例如,陆地生态系统包含多种土地利用类型,如森林、农田、草原、灌丛、冰川和沙漠,这些类型分布在从缓坡到高海拔的不同地貌中,且在热带、亚热带和温带生物群系的湿润至干旱区域,具有不同的地质起源和水文过程。
在特定土地利用类型的生态系统内,土壤理化性质(如pH值、温度、含水量、有机质含量、质地、孔隙度、氧化还原状态等)存在异质性,而这些性质均会影响氢气氧化的生物过程。尽管氢氧化细菌在各类景观中广泛存在,但土地利用类型仍会影响土壤的氢气吸收能力。例如,耕地土壤的氢气吸收活性比牧场土壤高10-20%(Popelier等,1985);森林土壤的氢气沉积速率比耕地土壤高约30%(Yonemura等,2000b)。Smith-Downey等(2008)的研究发现,森林土壤的氢气吸收能力比沙漠土壤高30-40%。综合来看,根据不同土地利用类型土壤的氢气吸收能力(落叶林>草原>耕地),森林被认为是清除大气氢气能力最强的生态系统(Khdhiri等,2015)。
尽管有研究报道沙漠土壤的氢气吸收活性低于其他类型土壤,但基因组分析显示,沙漠土壤中存在丰富的氢氧化细菌群落(Jordaan等,2020)。沙漠土壤中的氢氧化细菌似乎已通过适应性进化,利用氢氧化功能在干旱环境中生存(Ji等,2017)。一项针对森林土壤、海水和南极沙漠土壤中微量气体氧化菌丰度的调查显示,南极沙漠中高亲和力氢氧化菌的相对丰度是森林土壤的3倍(Greening和Grinter,2022)。
然而,在不同土壤系统中,氢气吸收活性与hhyL或Huc基因(均为氢氧化相关功能基因)的丰度之间并无一致关联,这是因为土壤理化环境是决定细菌基因转录、酶生物合成及氢气代谢过程的关键因素。因此,为理解陆地生态系统对全球氢气清除的贡献,我们必须研究不同土壤环境和土地利用类型中调控氢气氧化的关键土壤因子。
2.2 氢氧化微生物
氢氧化微生物以氢气(H₂)为能量来源进行代谢。从微生物能量学角度来看,这些原核生物利用氢气产生的能量维持基础代谢过程,部分微生物还可利用该能量固定大气中的二氧化碳(CO₂)(Piche-Choquette和Constant,2019;Meredith等,2017;McLearn和Dong,2002)。因此,氢氧化原核生物会根据终端电子受体的可利用性及热力学限制,以氢气(H₂)为电子供体,将终端电子受体还原。这些终端电子受体包括二氧化碳(CO₂)、硫酸根(SO₄2⁻)、三价铁离子(Fe3⁺)、硝酸根(NO₃⁻)、碳酸氢根(HCO₃⁻)和氧气(O₂)。
然而,氢氧化反应在微生物系统发育中并不具有保守性。目前已在51个细菌和古菌门的基因组中检测到吸收型氢化酶(即氢氧化酶),这些类群包括放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、蓝细菌门(Cyanobacteria)、广古菌门(Euryarchaeota)、厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)等(Peters等,2015;Greening等,2016;Saavedra-Lavoie等,2020)。在36个细菌门中发现了参与氢气吸收的最主要吸收型[NiFe]-氢化酶同工酶,这表明氢氧化细菌是土壤中氢氧化微生物的优势类群,占比超过70%(Vignais和Billoud,2007;Greening等,2015)。根据酶对氢气的亲和力,氢氧化细菌可分为高亲和力和低亲和力两个类群。
2.2.1 高亲和力氢氧化细菌
高亲和力氢氧化细菌能在低氢气浓度(米氏常数Km<100体积百万分比,ppmv)下以氢气为能量来源,尤其能适应大气氢气浓度(0.53 ppmv)。这类细菌每年可氧化全球约80%的大气氢气,且平均约39%的微生物群落中含有编码高亲和力氢化酶的基因(Bay等,2021)。
首个被分离的高亲和力氢氧化微生物是链霉菌属菌株PCB7(Streptomyces sp. PCB7),该菌株可 scavenge(清除)大气中的氢气(Constant等,2008a)。随后,研究人员在阿维链霉菌(Streptomyces avermitilis)、链霉菌属菌株HP3(Streptomyces sp. HP3)等多种链霉菌菌株中也证实了其氧化大气氢气的能力(Constant等,2010)。这一发现推动了高亲和力氢氧化酶(第5组/1h组吸收型[NiFe]-氢化酶)编码基因的鉴定,其中关键基因为大小亚基基因hhySL。研究确定,第5组氢化酶对氢气的米氏常数(Km)高得超出预期,达3.6±0.5微摩尔(μm)(Schäfer等,2013)。
hhySL基因先转录生成小亚基(约30千道尔顿,kDa)和大亚基(约60 kDa),随后组装成异源二聚体蛋白。小亚基中含有3个铁硫簇,大亚基则包含活性中心(镍-铁中心),该活性中心通过分子通道催化氢气氧化(Benoit等,2020)。对高亲和力吸收型[NiFe]-氢化酶大亚基编码基因hhyL的基因组数据库挖掘显示,放线菌门(Actinobacteria)中该酶的转录遗传潜力最高。此外,在酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycete)、疣微菌门(Verrucomicrobia)、α-变形菌纲(Alphaproteobacteria)、硝化螺旋菌门(Nitrospirae)、β-变形菌纲(Betaproteobacteria)、δ-变形菌纲(Deltaproteobacteria)和拟杆菌门(Bacteroidetes)中,也检测到了高亲和力吸收型[NiFe]-氢化酶的遗传潜力(Liot和Constant,2016;Giguere等,2021)(图4)。
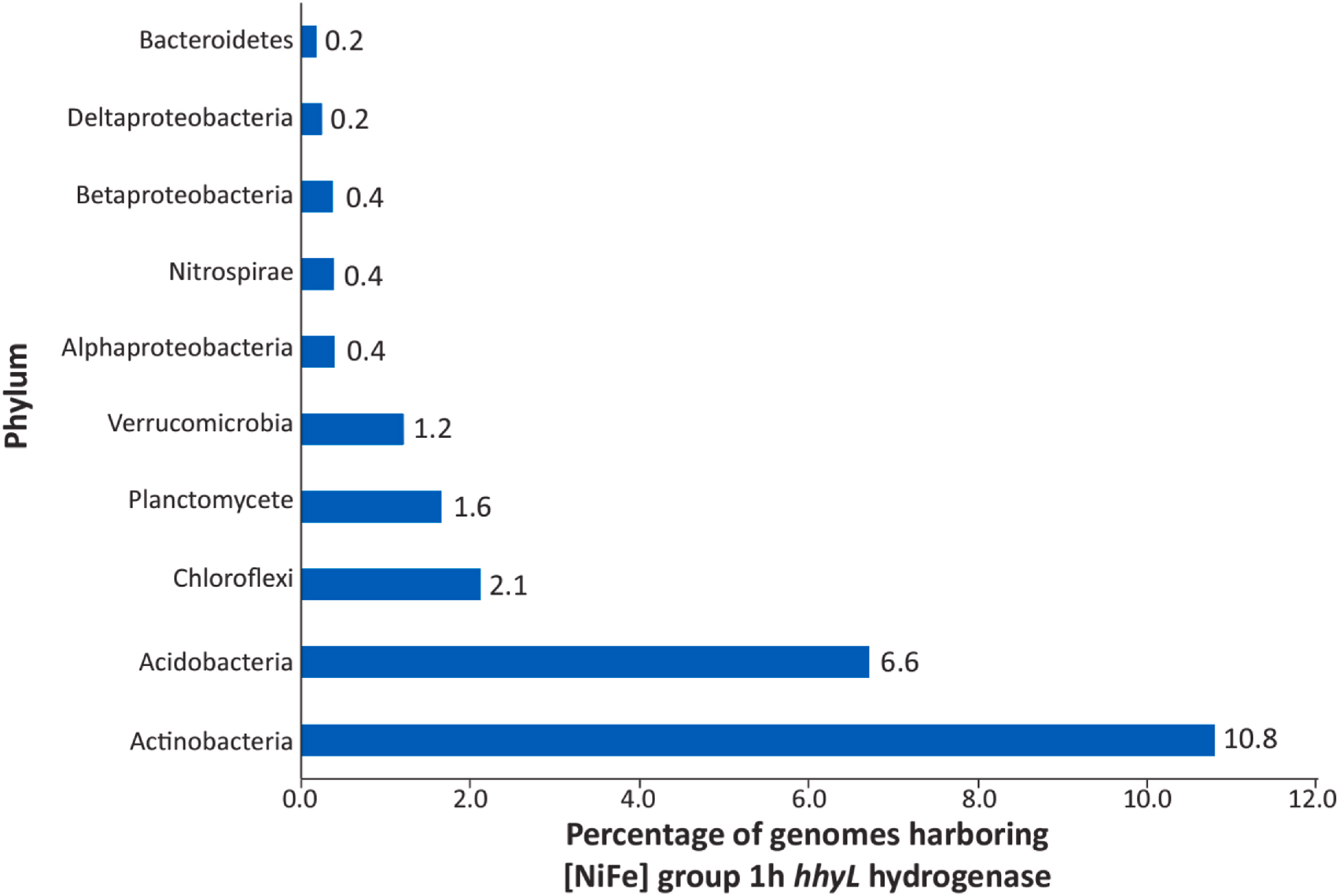
图4 不同门水平类群中含有高亲和力氢化酶基因(hhyL)的基因组及宏基因组组装基因组(MAGs)占比**(Giguere等,2021)
在含有hhySL基因的链霉菌属菌株PCB7(Streptomyces sp. PCB7)中,高亲和力氢氧化活性(Km<100 ppmv)仅存在于成熟孢子中(Constant等,2008a)。考虑到氧化大气氢气产生的能量(反应式:H₂→2H⁺+2e⁻,ΔGo’=-237 kJ mol⁻1;Koch等,2014)不足以支持化能无机营养生长,研究人员提出:放线菌门(Actinobacteria)及其他大气氢气清除菌(如耻垢分枝杆菌Mycobacterium smegmatis)在混合营养生长过程中,会将大气氢气的能量潜力与有机碳、甲烷(CH₄)、一氧化碳(CO)等物质结合利用,以满足自身维持能量需求(Jordaan等,2020;Bay等,2021)。
已鉴定的高亲和力氢氧化细菌——耻垢分枝杆菌(Mycobacterium smegmatis)通过另一种机制实现氢氧化:其编码的Huc基因可合成第2a组(Hyd 1型或蓝细菌型)吸收型氢化酶(Greening等,2014)。SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE)分析显示,Huc蛋白由3个亚基组成,分别为大亚基HucL(约58 kDa)、小亚基HucS(约39 kDa)和中间亚基HucM(作为膜连接“钩子”的四聚体,约19 kDa)(Grinter等,2023)。对纯化Huc酶的动力学分析表明,该酶对大气氢气具有高亲和力(Km=129纳摩尔,nM)和低氢气阈值(<31皮摩尔,pM),证实其在低氢气浓度下具有高效氧化能力。
Huc基因分布于13个门、171个属中,这一现象表明土壤微生物群落中编码HucL大亚基的遗传潜力显著。含有Huc基因的类群按丰度排序依次为:蓝细菌门(Cyanobacteria)、变形菌门(Proteobacteria)、拟杆菌门(Bacterioidota)、放线菌门(Actinobacteriota)、厚壁菌门(Firmicutes)、绿弯菌门(Chloroflexota)、产水菌门(Aquificota)、异常球菌-栖热菌门(Deinococcota)、硝化螺旋菌门(Nitrospirota)、浮霉菌门(Planctomycetota)、芽单胞菌门(Gemmatimonadota)、酸杆菌门(Acidobacteriota)和粘球菌门(Myxococcota)(Islam等,2020)。
尽管Hhy(第5组/1h组)和Huc(第2a组)均属于高亲和力吸收型氢化酶,但二者活性存在差异:Huc在指数生长期后期和稳定期早期表现出活性,为混合营养生长及从活跃生长向休眠期过渡过程中的能量储存提供支持;相反,当指数生长期显著延长时,Hhy活性会升高,这表明当碳储备减少时,细菌会依赖氢氧化作用为细胞维持提供能量。由于hhy是大多数土壤中主要的吸收型氢化酶基因,因此Hhy(第5组/1h组)被认为是全球氢气循环中土壤汇的主要贡献者(Islam等,2020)。除上述两种代表性高亲和力吸收型氢化酶外,第1f组(基因WP_026441619.1,Myers和King,2016)和第1l组(HylL,Ortiz等,2020)氢化酶也能氧化大气氢气。
2.2.2 低亲和力氢氧化细菌
另一类重要的氢氧化细菌是具有低亲和力氧化酶的类群。低亲和力氢氧化细菌能在高氢气浓度(Km>1000 ppmv)下以氢气为能量来源,这类细菌常见于氢气浓度较高的环境中,典型生境包括:固氮豆科植物根瘤表面(如黄杆菌属Flavobacterium)、自由生活的固氮菌内部(如蓝细菌门Cyanobacteria)、土壤大型动物肠道以及厌氧发酵器中(如脱硫脱硫弧菌Desulfovibrio desulfuricans)(图2)。
低亲和力氢氧化细菌含有编码吸收型氢化酶的基因,该酶具有功能多样性。因此,低亲和力吸收型氢化酶可根据其活性中心性质、功能及在细胞内的定位进行分类(Greening等,2016)。与高亲和力氢氧化细菌不同,低亲和力氢氧化细菌可利用氢气产生的能量合成碳水化合物(CH₂O)以支持繁殖,而非依赖氢气获取维持能量和生存。例如,莫斯科硝化螺旋菌(Nitrospira moscoviensis)能以顶空浓度为70%(体积比,v/v)的氢气(相当于在液体培养基中溶解浓度为0.37-0.52毫摩尔,mM)作为唯一能量来源,通过固定二氧化碳(CO₂)实现繁殖(Koch等,2014)。此外,莫斯科硝化螺旋菌(N. moscoviensis)还具有高亲和力氧化能力,可组成型氧化大气氢气(Leung等,2022),体现出功能多样性。
研究人员还发现了中等亲和力氢氧化细菌,其氢氧化活性介于高、低亲和力类群之间(Klüber和Conrad,1993)。尽管可根据氢化酶对氢气的亲和力(Km值)对氢氧化细菌进行分类,但无法将实验室纯培养菌株的Km值与宏基因组鸟枪法测序检测到的单一及混合种群Km值进行直接比较。如何解析不同土壤微生物群落中氢化酶的酶动力学特征,仍是未来研究面临的挑战。
2.3 影响土壤中氢氧化细菌的因素
2.3.1 氢气扩散
氢气(H₂)的扩散速率可能是土壤中氢氧化作用的限制因子。氢气的扩散系数因物理环境而异,例如,在空气中的扩散系数约为1.2×10⁻⁴平方米/秒(m2/s),在纯水中则为5×10⁻⁹平方米/秒(Strauch等,2023)。尽管氢气可通过土壤中的充气孔隙和充水孔隙扩散,但当含氢气的空气扩散到土壤深层时,氢气很可能被氧化。Myagkiy等(2020)建立了一个模型,该模型结合土壤物理性质(如曲折度、孔隙度、气体饱和度)和细菌消耗速率,用于估算土柱中氢气的扩散-反应过程(如图5所示)。

图5 土壤氢气氧化与土壤理化因子相关的概念模型
2.3.2 土壤pH值
土壤pH值通过影响底物(如营养物质)对微生物的溶解度和可利用性,显著调控化学反应与生物功能。一项针对砂质粘壤土森林的季节性大气氢气吸收监测研究显示:在酸性森林土壤(pH 4.6-5.4)、中性堆肥土壤(pH 7-7.2)和中性园土(pH 7.3-7.4)中,氢气吸收活性(包括高亲和力与低亲和力活性)均与土壤pH值变化呈负相关(Schuler和Conrad,1991)。
低亲和力与高亲和力土壤氢气吸收活性的最适pH值存在差异:对于高亲和力氢气吸收活性,其最适pH值与土壤原始pH值接近(园土最适pH为8,酸性森林土壤最适pH为5);相反,酸性森林土壤中低亲和力氢气吸收活性在两个pH值(pH 4和pH 6.7)下达到峰值,这表明低亲和力氢氧化菌群的最适pH值可能与高亲和力氢氧化菌不同。由于低亲和力与高亲和力氢氧化菌在不同系统发育类群中的分布不均匀,因此它们具有多个最适pH值具有合理性。此外,土壤pH值是微生物群落结构的重要驱动因子,这意味着氢氧化细菌群落的活性可能随pH值变化而改变。因此,需通过研究土壤pH梯度下的氢氧化细菌群落,进一步解答这一问题。
尽管土壤pH值被认为会影响土壤氢气吸收,但这种影响可能取决于测定对象是可培养分离菌株还是田间采集土壤。部分高亲和力氢氧化细菌纯培养物在中性pH值附近表现出最优活性(Schuler和Conrad,1991),但一项涵盖38个采样点的田间调查显示,土壤氢气氧化活性与pH值无相关性(Popelier等,1985)。由于Schuler和Conrad(1991)直接使用强酸(HCl)和强碱(NaOH)调控土壤pH值,这可能对土壤微生物的生长和活性产生了干扰。因此,需要开发替代方法来研究土壤氢气吸收活性与pH值的关系。
2.3.3 土壤温度
土壤温度通过调控大气与土壤间的气体交换(如氢气扩散)影响氢氧化过程。在室温和大气压下,氢气在空气中的扩散速度约为0.7米/秒(m·s⁻1);但在土壤中,实际扩散速率会因土壤性质(包括土壤质地、含水量、温度和紧实度)而变化(假设氢气不与土壤发生反应)。温度升高会增加氢分子的动能,从而加快扩散速率,因此温暖土壤中的氢气扩散速率通常高于寒冷土壤。
已有田间与实验室研究探讨了土壤温度对氢气吸收活性的影响。在一项田间研究中,Schuler和Conrad(1991)将土壤置于5-60°C的温度范围内,并暴露于低、高两种氢气浓度下,结果发现氢气吸收活性存在两个明显的最适温度:高亲和力氢气吸收活性(暴露于低氢气浓度)的最适温度为35-40°C,而低亲和力氢气吸收活性(暴露于高氢气浓度)的最适温度为55°C。这一现象可能是由于温度对土壤中同步发生的化学和生物反应产生影响,进而调控了受试土壤中的氢化酶活性。
在一项受控实验室研究中,Smith-Downey等(2006)发现高亲和力土壤氢气吸收活性的最适温度为10-30°C:在-15°C时未检测到高亲和力氢气吸收活性;在-4-0°C范围内,活性呈指数增长;在0-10°C范围内,活性略有上升;而在10-30°C范围内,活性相对稳定。由此可见,氢气吸收的最适温度与自然环境中的土壤温度相关。我们认为,土壤温度是调控土壤氢气吸收活性的重要参数,在全球变暖背景下需重点关注。
2.3.4 土壤含水量
土壤含水量除为生物提供水分外,还调控微生物细胞外部环境的离子浓度及离子向细胞表面的迁移。所有土壤微生物(包括氢氧化细菌)均需水分维持内环境稳定,这一点对沙漠土壤中的氢氧化细菌尤为关键——氢氧化作用可能为它们生成水分,以满足自身 hydration(水合)需求。
以下观测结果支持这一观点:沙漠土壤中携带高亲和力氢气吸收氢化酶基因的微生物数量多于森林土壤;且水分补充可刺激沙漠土壤中的氢气吸收活性(Jordaan等,2020;Greening和Grinter,2022)。一个具有启发性的推测是:沙漠微生物群落可合成高亲和力氢氧化酶,通过氧化氢气生成水,从而提高在干旱环境中的存活能力。若该推测成立,那么在沙漠和干旱环境中,获取大气氢气作为能量来源,将是土壤微生物群落应对长期缺水的重要适应策略。
土壤含水量还会影响氢气从大气向土壤剖面的扩散过程。这是因为土壤孔隙中空气和水的比例不同,而氢气在水中的扩散速率与在空气中存在差异(Strauch等,2023)。此外,氢气需先溶解于水中,才能到达氢氧化细菌细胞——这类细菌通常生活在土壤表面的生物膜中(周围环绕着毛管水),或作为附着/运动细胞存在于土壤孔隙中循环的水中。
此外,需明确含水量是否会使土壤微生境成为氢气的净汇或净源:在渍水土壤中,发酵作用可能产生氢气。实验室研究表明,氢气吸收活性的最适土壤含水量为6%-20%(Smith-Downey等,2006)。在田间研究中,研究人员采用微气象学方法测定了魁北克森林生态区的氢气通量,发现土壤氢气吸收活性与水分蒸发呈正相关(Constant等,2008b)。尽管受试土壤仍含有足够的水分通量(50克·平方米⁻2·小时⁻1)以支持微生物活性,但水分蒸发增加了土壤中的充气孔隙度,从而提高了氢气的可利用性(Constant等,2008b)。
2.3.5 土壤碳含量
土壤总碳含量会导致土壤氢气氧化活性产生差异(Khdhiri等,2015)。对38个土壤样本的分析显示,氢气氧化活性随土壤有机质含量的增加而显著升高(Popelier等,1985;图6)。牧场土壤的碳含量高于耕地土壤,粘土的碳含量高于壤土和砂质土壤,且碳含量较高的土壤往往具有更高的氢气吸收速率。
这一现象可通过以下机制解释:大多数高亲和力氢氧化细菌为混合营养型微生物,即它们同时利用氢气与其他底物(如有机碳、甲烷、一氧化碳等)作为能量来源。在这种情况下,土壤碳含量升高会增加氢氧化细菌的丰度。然而,向土壤中添加碳可能会抑制细菌的氢氧化活性(Baril和Constant,2023),推测原因是充足的碳源使细菌不再需要将氢气作为替代能量来源。上述观测结果仍需进一步研究验证。
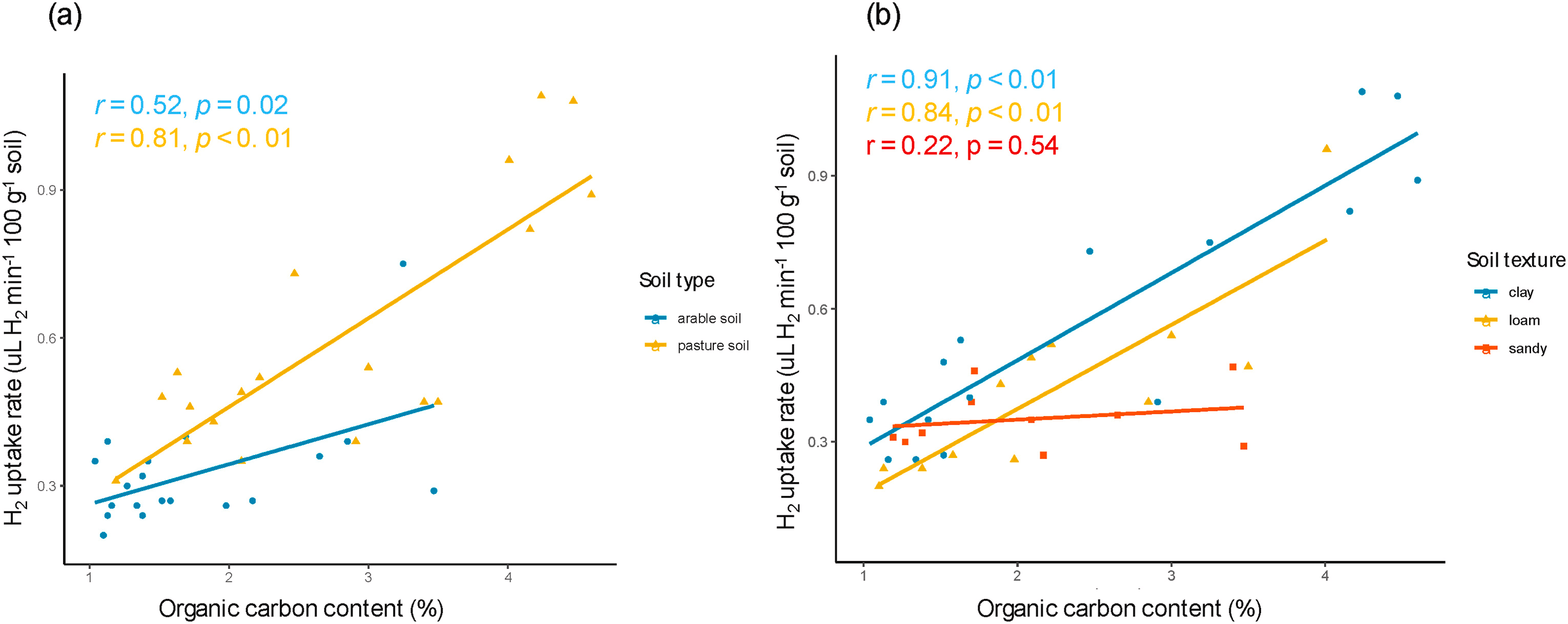
图6 土壤有机碳含量与土壤氢气吸收速率的相关性
(a)耕地和牧场土壤;(b)砂质、壤质和粘质土壤(数据来源于Popelier等,1985)
2.3.6 土壤质地
土壤质地直接影响土壤理化性质。例如,砂粒、粘粒和粉粒的含量会直接调控土壤孔隙度和团聚体结构;而土壤孔隙度的变化会进一步影响土壤通气性、含水量和氢气扩散速率,这些参数均会直接或间接影响土壤氢气氧化活性。
一项田间调查显示,土壤氢气氧化活性与粘粒含量呈正相关(Popelier等,1985)。这可能与介质的孔隙特性有关:粘粒对分子氢的吸附作用,提高了定殖于粘粒表面并形成生物膜的氢氧化细菌对氢气的可利用性(即细菌通过钙配体与粘粒结合)(Wolff-Boenisch等,2023;Zhu等,2023)。
2.3.7 土壤中氢气氧化反应的抑制剂
吸收型氢化酶的底物是氢分子(H₂),该酶通过以下反应将分子氢转化为氢离子(H⁺)和电子(e⁻):
H₂ → 2H⁺ + 2e⁻
吸收型氢化酶的抑制剂可部分或完全干扰酶活性。例如,氰离子(CN⁻)通过直接结合酶的活性中心(尤其是[NiFe]-吸收型氢化酶活性中心中的铁(Fe)和镍(Ni)),抑制氢化酶活性。氰离子与这些金属中心结合形成稳定复合物,会阻断活性中心,从而干扰酶结合和氧化氢气的能力(Vincent等,2005)。
由于吸收型氢化酶通过催化作用将电子转移至膜结合呼吸链,因此呼吸链的组成成分(如膜可溶性甲基萘醌(MQ)和甲基萘醌醇(MQHs))也可抑制氢化酶活性(Greening等,2015)。此外,细胞色素bcc-aa3超复合物、细胞色素bd和ATP合成酶会间接抑制吸收型氢化酶的活性,相关抑制剂包括:
- 叠氮化锌(细胞色素bcc-aa3氧化酶的选择性抑制剂);
- 2-庚基-4-羟基喹啉-N-氧化物(醌结合的竞争性抑制剂);
- 缬氨霉素(一种离子载体,可与钾离子(K⁺)结合形成带正电复合物,沿电势梯度特异性地将钾离子转运穿过细胞膜,破坏呼吸细菌质子动力势中的电势成分);
- 尼日利亚菌素(一种离子载体,作为钾离子和氢离子(H⁺)的反向转运体,在结合离子后呈电中性)。
此外,细胞质酸化会通过破坏酶的结构完整性、改变活性中心的电化学性质、影响催化反应中氢气的可利用性,从而抑制吸收型氢化酶活性(Cordero等,2019)。
2.4 农业管理措施对土壤氢气吸收活性的影响
耕作、有机改良和施肥等农业措施会显著改变土壤微生物群落(包括氢氧化细菌)。例如,有机农业实践或添加有机质(如堆肥)可提高微生物活性,进而可能增加氢气吸收速率;氮肥(尤其是铵态氮肥)会影响土壤微生物群落及其代谢活性(包括氢气氧化)。过量施肥或不当的养分管理可能破坏微生物群落,导致氢气吸收活性降低。
此外,能够提高土壤微生物多样性和活性的多样化种植制度与覆盖作物(尤其是作物轮作中包含豆科作物时),有望促进氢气吸收。
3 基于生态特征评估土壤氢气吸收能力的方法
3.1 与土壤氢气吸收能力相关的生态特征定义
尽管已在51个门中检测到氢氧化酶基因,但从土壤中分离培养并用于详细研究的氢氧化细菌数量相对较少。目前关于氢氧化细菌的表型和基因组特征、土壤中的分布规律及其生物学、生态学和环境功能的信息仍较为有限。我们可推测,氢氧化细菌具有适应低能量、低土壤含水量及其他胁迫环境的生态和生物学策略。
为揭示高亲和力氢氧化细菌之间的潜在共性,研究人员提出采用基于特征的方法,评估其微生物多样性和生态系统功能(如元素生物地球化学循环)(Krause等,2014)。通过对高亲和力氢氧化细菌进行生理生态学特征分析,可明确调控其在土壤中分布和活性的环境因子。尽管已有研究报道了氢氧化细菌的多样性(如分类组成)(Greening等,2016),但尚未明确其生态特征。
特征是指个体可测量的生理或形态属性,这些属性对其生存表现和最终适合度具有重要影响(Garcia等,2022)。通过特征可推断生物在不同环境条件下的生存表现(Violle等,2007)。理想情况下,研究人员应选择可在群落水平上测量或扩展的特征,并最终将其与群落和生态系统功能相关联。
微生物生态学家借鉴了植物科学中的生态特征概念(Martiny等,2015),将其定义为微生物的生理特征。这些微生物特征受生物和非生物因子共同作用形成,包括细胞化学组成、生活史、生物物理特性以及对环境和生物胁迫的适应能力(Sokol等,2022)。
# 生态特征可分为以下两类:
(1)**响应特征**:指微生物在非生物因子(如干扰、资源可利用性)发生变化时,维持生存、生长与繁殖的特性(例如,氢氧化细菌对碳类型及浓度的响应(Baril和Constant,2023)、氢氧化细菌对土壤pH梯度的响应(Hou等,2024));
(2)**效应特征**:指微生物影响环境条件、进而改变生态系统结构和/或时间动态功能的功能属性(Martiny等,2015;Loureiro等,2023)。
目前,反映氢氧化细菌在不同环境中分布、丰度、多样性及群落组成特征的生态响应特征仍不明确。已有部分研究对效应特征(如影响氢气吸收活性的pH值、温度、含水量、底物、抑制剂等因子)进行了评估,但仍需进一步验证。
在生态学中,**功能群(guild)** 指利用同一类资源的生物群体(Simberloff和Dayan,1991)。在微生物生态学中,功能群被定义为一组具有一致共丰度行为、且可能协同作用以实现同一生态功能的微生物群体,其分类不依赖于物种的系统发育地位(Wu等,2021)。由于氢气代谢并非土壤细菌的保守功能,因此可根据生态特征将氢氧化细菌划分为不同功能群。
3.2 生态特征的检测方法
微生物生态学中的生态特征研究,聚焦于微生物在自然环境中的功能特性与行为。通过检测生态特征,科学家能够理解微生物与周围环境及其他生物间的相互作用,进而明确生态系统功能。目前用于检测微生物生态特征的方法包括:
(1)**宏基因组鸟枪法测序(Shotgun Metagenomics)**:对微生物群落的全基因组进行测序,鉴定群落中存在的基因及功能通路,从而揭示微生物潜在的生态特征(Qin等,2010)。
(2)**功能基因芯片(Functional Gene Microarrays)**:检测参与关键生态过程(如氮循环、碳降解、抗生素抗性)的特定功能基因,通过基因的有无推断微生物群落的潜在特征(He等,2012)。
(3)**稳定同位素探针技术(Stable Isotope Probing, SIP)**:将群落中特定微生物物种与特定生态特征关联。该技术通过稳定同位素标记特定底物(如碳源),追踪其在微生物生物量中的整合过程,从而识别执行特定生态功能的微生物(Dumont和Murrell,2005)。
(4)**宏转录组学(Metatranscriptomics)**:对微生物群落中的RNA转录本进行测序,揭示特定时间点微生物活跃表达的基因,明确当前发挥作用的功能特征(Poretsky等,2009)。
(5)**微生物功能谱分析(Microbial Functional Profiling)**:利用GeoChip或PICRUSt等工具,基于微生物群落的分类组成预测其功能潜力,进而解读群落基因组中编码的生态特征(Langille等,2013)。
(6)**功能测定(Functional Assays)**:例如,通过实验室测定直接检测纯培养微生物的特定生态特征,如酶活性、代谢速率、对极端条件的耐受性等(Martiny等,2015)。
为全面理解特定生态系统中微生物的生态特征,通常需结合多种方法(包括分子技术与功能测定)。这种多学科方法能帮助研究人员建立微生物群落组成与生态系统功能及恢复力之间的关联。
3.3 生态特征整合到全球氢气平衡中的进展与挑战
整合生态特征可通过提高氢氧化菌活性的时空分辨率,提升全球氢气平衡估算的准确性。尽管微生物生态学家已采用改良的基于特征的研究方法(Martiny等,2015),但关于氢氧化细菌对其他环境变量响应的生态特征仍知之甚少。
其中一个局限性是缺乏用于分类高亲和力氢氧化细菌的标准体系。除氧化大气氢气的能力外,高亲和力氢氧化细菌还具有其他效应特征,例如:降解难降解有机质以维持生存、共氧化一氧化碳(CO)与甲烷(CH₄)以获取足够能量支持混合营养生长、参与硝化与反硝化过程等(Greening和Grinter,2022;Constant等,2009)。
已有研究表明,高亲和力氢氧化作用通常发生在细菌面临能量匮乏的环境中(Greening等,2022),这提示高亲和力氢氧化细菌可能为寡营养型微生物。然而,一项针对碳响应特征的研究发现,高亲和力氢氧化细菌对碳源的偏好存在差异,表明这类细菌在分类学上具有不均匀性(Baril和Constant,2023)。高亲和力氢氧化细菌的纯培养菌株具有革兰氏阳性、非运动性等表型特征,而低亲和力氢氧化细菌则多为革兰氏阴性、运动性微生物。但目前这些分类特征对氢氧化细菌而言仍定义模糊。
因此,我们建议采用基于特征的研究方法,将高亲和力氢氧化细菌群落的环境功能与其基因组信息关联。这一方法可筛选出更具代表性的特征,用于分类高亲和力氢氧化细菌,并明确其在土壤生态系统及氢气循环中的作用。
4 结论与未来展望
高亲和力氢氧化细菌使土壤成为大气氢气(一种间接温室气体)的主要汇。对高亲和力氢氧化细菌的研究,有助于深入理解其在土壤环境及全球氢气循环中的生物多样性、生物学特性与生态学功能。然而,高亲和力氢氧化细菌在系统发育上并不具有特异性,这可能是由于氢氧化基因在原核生物间发生水平转移所致。
关于高亲和力氢氧化细菌,仍有许多关键生态问题亟待解决,例如其空间分布、代谢功能、调控氢氧化菌活性的环境因子等,这些问题值得进一步关注。由于包括氢氧化菌在内的大多数土壤微生物难以培养,无法通过纯培养方法推导其生物学与生态学特征。
我们建议通过对高亲和力氢氧化菌的标志基因(hhyL(Giguere等,2021)、hucL(Islam等,2020)、第1f组基因WP_026441619.1(Myers和King,2016)、第1l组基因HylL(Ortiz等,2020))进行宏基因组测序,量化高亲和力氢氧化菌的数量、类型及氢化酶类别。该方法可根据高亲和力氢氧化细菌对环境的响应及其在全球氢气平衡中的潜在功能,明确其生态特征。
高亲和力氢氧化细菌的效应特征与响应特征,可作为评估和量化这类细菌在全球氢气循环中作用的依据。随着大气氢气浓度持续升高,通过扩大细菌分布与活性的研究尺度,我们能够更准确地估算不同土壤及陆地生态系统中大气氢气的氧化量。
https://blog.sciencenet.cn/blog-41174-1514259.html
上一篇:重组层粘连蛋白为帕金森病干细胞疗法铺平道路
下一篇:研究表明:老年女性过量饮用咖啡不利于骨健康
