博文
围绕DNA分子双螺旋结构模型的建立展开的一场激烈竞争  精选
精选
||
围绕DNA分子双螺旋结构模型的建立
展开的一场激烈竞争
冯永康
(生命科学史研究者)
书比人更有趣,阅读使人生充满意义!
——J.D.Watson
新事物只有建立在现存的传统之上,才最有生命力。
—— F.Crick
早在1912年,英国结构化学家亨利·布拉格(W.H.Bragg,1862—1942)和劳伦斯·布拉格(W.L.Bragg,1890—1971) 父子两人,提出并逐渐发展起来的X射线晶体衍射法,为40年后解开DNA的结构之谜提供了关键性的技术。
1930年代末到1950年代,奥地利出生的晶体学家佩鲁茨(M.F.Perutz,1914—2002)和英国化学家肯德鲁(J.C.Kendrew,1917—1997),在劳伦斯·布拉格领导的剑桥大学卡文迪什实验室,分别对马的血红蛋白和鲸的肌红蛋白进行晶体结构的研究。他们创立了把重原子引入蛋白质分子的分析方法,并应用计算机处理衍射资料,提高了分析精确度和速度。这些工作为DNA分子双螺旋结构的建立提供了直接的实验基础。
1944年,美国科学家艾弗里(O.T.Avery,1877—1955)及其同事, 以肺炎双球菌为材料,利用酶学分析实验技术,通过破坏失活的思路,以试图确定“转化因子”的目的。他们首先着手进行了含有R→S转化因子的S细菌的无细胞提取液的分馏工作。在使用一系列的化学和酶催化方法把各种蛋白质、脂质、多糖和RNA从提取液中去掉时,不断检测被纯化留下的提取物(转化因子)把R突变型转化为S野生型的能力。实验结果发现,当加入DNA水解酶后,便观察不到转化现象的发生(即转化因子的活性丧失),这说明DNA是R→S转化所必需的“转化因子”。这个重要发现,首次用实验证明了遗传物质就是DNA。9年后的1953年,因提出DNA分子双螺旋结构模型而声名大振的沃森(J.Watson,1928— )曾经赞叹道:“艾弗里的实验,使我们闻到了DNA是基础遗传物质的气息。”
1948年,美国生物化学家查伽夫(E.Chargalf,1905—2002)在读到艾弗里的论文后,深受启迪。他认为如果不同的生物种类是由于DNA的不同,则DNA的结构必定十分复杂,否则难以适应生物界的多样性,因此他对由美国生物化学家列文(P.A.Levene,1869—1940)提出的已统治科学界30多年的“四核苷酸假说”,产生了怀疑。1949—1952年,查伽夫采用纸层析法分离碱基,再用紫外吸收光谱作定量分析,发现DNA的碱基成分随生物种类的不同而有很大差异,而A和T、G和C的分子数总是相等。这就意味着,DNA分子中四种脱氧核苷酸的排列顺序可能蕴藏着大量的信息。查伽夫的实验研究,为沃森和克里克(F.C.Crick,1916—2004)建立DNA分子的双螺旋结构中起关键作用的碱基配对方式,奠定了坚实的实验基础。
到了1950年代初期,有三个实验室为了一个共同的目标——找到合理的结构模型来阐述DNA分子的遗传作用,展开了一场争分夺秒的激烈竞争。遗传学的研究,处在一个重大突破的前夜。
第一个实验室是美国加州理工学院著名化学家鲍林(L.C.Pauling,1901—1994)的实验室。1950年,鲍林等利用X射线晶体衍射技术研究蛋白质结构,提出蛋白质是长链分子,并发现了α螺旋结构。对蛋白质研究的成功,大大激励了他们把该技术用于DNA分子结构的研究。
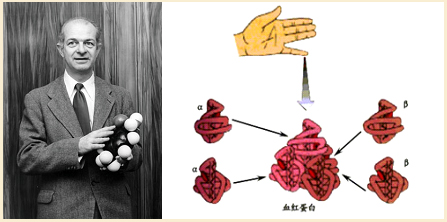
鲍林利用X射线晶体衍射技术,发现血红蛋白的α螺旋结构
1952年底,鲍林等人根据阿斯特伯里的研究照片,明确提出DNA分子并非单链结构,而可能是双链或三链的螺旋体。由于鲍林研究团队缺乏足够的资料,所用的X射线照片图像也不够清晰,便得出了在DNA分子结构中,磷酸-脱氧核糖骨架位于螺旋体内部、碱基排在外侧的错误认识。当时,鲍林的注意力主要集中在蛋白质结构研究上,根本没想到DNA需要他全力以赴去做研究。鲍林还非常自负地认为,其他人未必是他的竞争对手。当他最终为寻找一种合理的结果继续研究DNA分子的结构时,一切都已经晚了,成功与鲍林擦肩而过。
第二个实验室在英国皇家学院,由新西兰物理学家威尔金斯(M.Wilkins,1916—2004)和英国女物理学家弗兰克林(R.Franklin,1920—1958)领导,两人都专长于X射线结晶学的研究。
1950年,威尔金斯开始选择DNA纤维作为研究材料,弗兰克林于翌年加盟。他们于1952年设法制成了高度定向的DNA结晶纤维,并由弗兰克林拍摄出了非常清晰的X射线衍射照片。通过对这张照片的细致分析,他们推算出DNA分子是双链同轴排列的螺旋结构,磷酸根基团和脱氧核糖在螺旋外侧,碱基在螺旋内侧;并定量测定了DNA螺旋体的直径和螺距。
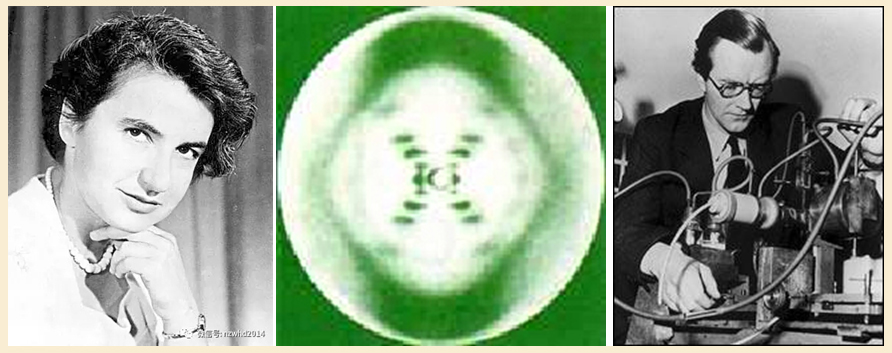
图左中为弗兰克林及拍摄出的DNAX射线衍射图;图右为威尔金斯
到1953年3月17日,DNA分子的结构问题已经差不多被弗兰克林所攻克。然而,他们却未能及时地将分析资料转变为一个合理的结构模型。作为物理学家,他们也未能真正理解DNA分子结构的重大生物学意义。再加上两人的关系一开始就磕磕碰碰,并发展到高度紧张,很难进行合作。
结果,让沃森和克里克领先了。
早在1946年,当沃森还是美国芝加哥大学的学生时,就被奥地利物理学家薛定谔(E.E.Schrödinger,1887—1961)出版的《生命是什么》小册子所吸引,对遗传学产生了浓厚的兴趣。
1947年,沃森来到了美国印地安那大学,这里有刚刚(1946年)获得诺贝尔生理学或医学奖的遗传学家缪勒(H.J.Muller, 1890-1967)在此任教。
沃森在美国遗传学家卢里亚(S.E.Luria,1912—1991)的指导下,开始研究X射线对噬菌体增殖的影响。卢里亚对DNA分子可能是遗传物质的预感,以及稍后与德国物理学家德尔布吕克(M.Delbrück,1906—1981)的第一次相见,都进一步影响了沃森,促使他产生了要了解DNA如何发挥遗传作用,就必须研究其分子结构的想法。
1951年春,已经取得博士学位的沃森,在参加意大利那不勒斯的一个关于生物大分子结构的学术会议上,有机会听取了威尔金斯所作的关于DNA的X射线衍射分析报告,便决定从事X射线晶体学的博士后研究。
是年秋,经卢里亚的引荐,沃森进入到英国剑桥大学由劳伦斯·布拉格领导的卡文迪什(Cavendish)实验室。在肯德鲁的指导下,沃森对植物病毒中提取的核酸分子结构进行研究。
也就是在英国的卡文迪什实验室,沃森与比他早两年进入该实验室跟随佩鲁茨作血红蛋白结构研究的克里克有幸相遇。克里克不仅是一位了解X射线结晶学,而且对DNA分子的结构与生物学功能也很感兴趣的物理学家。
两个年轻人的结识,开始了现代生物学最激动人心、最卓有成效的合作。
沃森和克里克决定共同研究DNA分子的结构,并确定了提出一个结构模型的目标。这个模型不仅要能够解释X射线衍射分析的资料,还必须能够阐明DNA的自我复制和控制蛋白质合成的机理。从1951年秋到1953年春,他们先后提出了三种结构模型。
1951年底,沃森和克里克根据X射线衍射照片和键距计算资料,提出了一个三链螺旋模型。由于他们所研究的X射线照片的质量太差,这个模型经威尔金斯和弗兰克林等人验证和核实,发现对实验资料的处理有明显错误,第一次模型的建立宣告失败。劳伦斯·布拉格将克里克调回蛋白质课题组,沃森被调回去继续做烟草花叶病毒的晶体衍射。但他们两人并没有就此罢休,继续偷偷地研究DNA。尤其是沃森热衷于DNA,“简直到了疯狂的地步”。
正当两人一筹莫展的时候,有三件事对沃森和克里克的研究思路,产生了十分重要的影响。
第一件事是沃森和克里克与剑桥大学年轻的数学家格里菲斯(J.Griffth)的偶遇。格里菲斯也对一些生物学问题比较感兴趣,便答应帮助他们计算DNA分子中碱基之间的吸引力。格里菲斯不久便告诉他们,理论计算表明碱基之间的结合力是A吸引T,G吸引C。这使克里克看到了碱基互补配对的可能性。
第二件事是查伽夫1952年7月到剑桥大学访问时,肯德鲁介绍他与沃森和克里克进行了一次非常重要的会面。当他们谈到核酸研究工作的进展时,查伽夫略微迟疑了一下回答道:“一句话说完,就是1:1。”克里克立即意识到,格里菲斯和查伽夫所说的碱基配对是完全一样的;不同类型的碱基配对,可能就是DNA分子结构的基础。
第三件事则是鲍林对蛋白质α螺旋结构的研究,为研究生物大分子提供了一个独特的物理方法,即先根据理论上的考虑建立模型,再用X射线衍射结果对模型进行检验。沃森和克里克两人深受启示,先后建立了一大堆有关嘌呤碱基和嘧啶碱基的模型,以确定什么样的维度和排列才既符合氢键的理论要求,又符合查伽夫碱基配对规则的经验要求。
1953年2月初,沃森和克里克看到了鲍林关于DNA研究论文的手稿,使他们大吃一惊的是,鲍林的模型与他们之前的那个三链结构很相似。他们清楚地意识到,待鲍林觉察出自己的错误再回过头来全力研究DNA,至少需要六周时间。
由于鲍林的论文,使沃森和克里克终于得到劳伦斯·布拉格的指示,重返DNA分子结构的研究,第二次模型建立又雄心勃勃地开始了。当时,可供考虑的研究方案已经缩小到是双链还是三链、碱基是排列在内侧还是外侧。
几天后,当沃森又一次访问英国皇家学院实验室,向威尔金斯讲述了鲍林的错误,并介绍了自己关于DNA分子螺旋结构的想法。威尔金斯随即谈到了弗兰克林拍摄的高度清晰的DNAX射线衍射照片。弗兰克林的研究已经发现,DNA存在着两种形式,一种处于缩合干燥状态,另一种处于扩张湿润状态。鲍林使用的阿斯特贝里的照片,是两种形式的混合,并且不太清楚。而弗兰克林新拍摄的DNAX射线衍射照片不仅清晰多了,而且纯粹是扩张的形式。也就是在该实验室里,在弗兰克林毫不知晓的那一刻,威尔金斯将弗兰克林新拍摄的B型DNAX射线衍射照片的拷贝取出给沃森看了一下。
正是弗兰克林拍摄出的这张清晰照片及其反映出的几个主要参数,为沃森和克里克的继续研究,提供了非常关键的信息。
于是,沃森和克里克又重新开始忙着一个双链螺旋结构模型的设计。但这个模型由于是采用了相同的碱基配对,当即便受到了在同一个实验室里工作的美国结构化学家多诺休(J.Donohue,1920—1985 )的质疑。多诺休建议他们将模型的构型由烯醇型改为酮型。这一合理的主张,使他们茅塞顿开。一直困扰他们的碱基配对问题,终于得到了解决。
克里克在后来的回忆录中写道:“太妙了!我们可以用这种特异的碱基配对方式,来说明查伽夫的1:1的碱基比例。就在那一刻(1953年2月20日),我们三个人都明白了,排列在DNA分子结构内侧的碱基,是靠氢键来形成A—T、G—C这两种碱基对的。”
在这种DNA的双螺旋结构模型中,A与T配对,G与C配对,两条相互缠绕的链上碱基顺序彼此互补,结合在一起,就能形成符合X射线衍射资料的螺旋。这样一来,查伽夫关于“碱基配对的1:1法则”,也就成了DNA双螺旋结构的必然结果。该模型还反映出,只要确定了其中一条链上的碱基排列顺序,另一条链上的碱基排列顺序也就确定了。沃森和克里克对这个双螺旋结构的模型经过逐项精确的检查,得到布拉格、威尔金斯和弗兰克林以及鲍林等学者的一致肯定,DNA分子的双螺旋结构模型便成功建立了。

沃森与克里克注视着刚刚建立的DNA分子双螺旋结构模型(1953)
从1951年11月到1953年4月,经过短短18个月紧张而有序的合作,一篇具有划时代意义的重要文献《核酸的分子结构——脱氧核糖核酸的结构》,于1953年4月25日在英国的《自然》(Nature)周刊上发了。就是在这篇不到1 000字的论文中,沃森和克里克不仅第一次从分子水平上,阐明了DNA分子的双螺旋结构和碱基互补配对的原则;基于该模型,他们还提出了一种DNA分子自我复制的可能机制。

沃森与克里克发表在Nature周刊上的研究论文
《核酸的分子结构——脱氧核糖核酸的结构》
这是一个在整个科学界都激动人心的时刻,称赞和喝彩从世界各地不断飞来,两位年轻人充满了喜悦。他们二人,一人是资历尚浅的博士后研究者,另一人是年龄偏大的研究生。然而,正是他们的亲密合作和锲而不舍的追求,才谱写出了生命科学史上的壮丽篇章。
DNA分子双螺旋结构模型的建立,为人们进一步探求遗传物质的自我复制和控制蛋白质合成的功能,奠定了坚实的基础,标志着人类对遗传和变异现象的研究进入到分子水平,也由此开创了遗传学研究的新时代。
半个世纪后的2003年,沃森和克里克又重逢在英国剑桥大学的卡文迪许实验室。凝视着当年搭建的DNA分子双螺旋结构模型,黑发已白,活力依然,感慨万千。

沃森与克里克50年后重访故地(2003)
1953年,沃森和克里克同心协力、执着求索,提出了DNA分子的双螺旋结构模型。随后的50年,他们又立于生命科学的潮头,亲身体验和见证了由DNA分子双螺旋结构模型所开创的分子生物学的大发展。
主要参考文献:
[1] J.D.Watson著.刘望夷等译.双螺旋——发现DNA结构的故事.北京:科学出版社,1984.
[2] 李佩珊,许良英主编.20世纪科学技术发展简史. 北京:科学出版社,1999.
[3] G E.Allen著.田洺译.20世纪的生命科学史.上海:复旦大学出版社,2000.
[4] R.Olby著.赵寿元、谌民家译.通往双螺旋之路: DNA的发现.上海:复旦大学出版社,2012.
[5] 庚镇城.纪念DNA双螺旋结构模型发表70周年.科学, 2023 (75) 4:58-61.
(选摘自《科学》杂志2003年第2期《生命科学史上的划时代突破——DNA分子双螺旋结构模型的建立》一文。本次发布时,作者进行了个别文字和插图的补充。)
(2024年4月15日)
https://blog.sciencenet.cn/blog-294644-1429774.html
上一篇:绿色教育志愿者的足迹 ——追求人与自然的和谐共存
下一篇:中国水稻遗传育种学的先驱赵连芳博士——写在赵连芳诞辰130周年之际
