博文
孔雀鱼MHC II基因的平衡选择和超类进化
||
对自然选择是达尔文进化论中的核心内容,表示在种群中促进有利变异的保留和扩散、消除有害变异的力量。但自然界中大多数性状不是只有“有或无”两种状态,而是呈现出一种连续的分布。作用于性状分布的自然选择,根据其作用的位置和方向,可以再分为定向选择(directional selection)、歧化选择(diversifying selection)和稳定选择(stabilizing selection)三种。
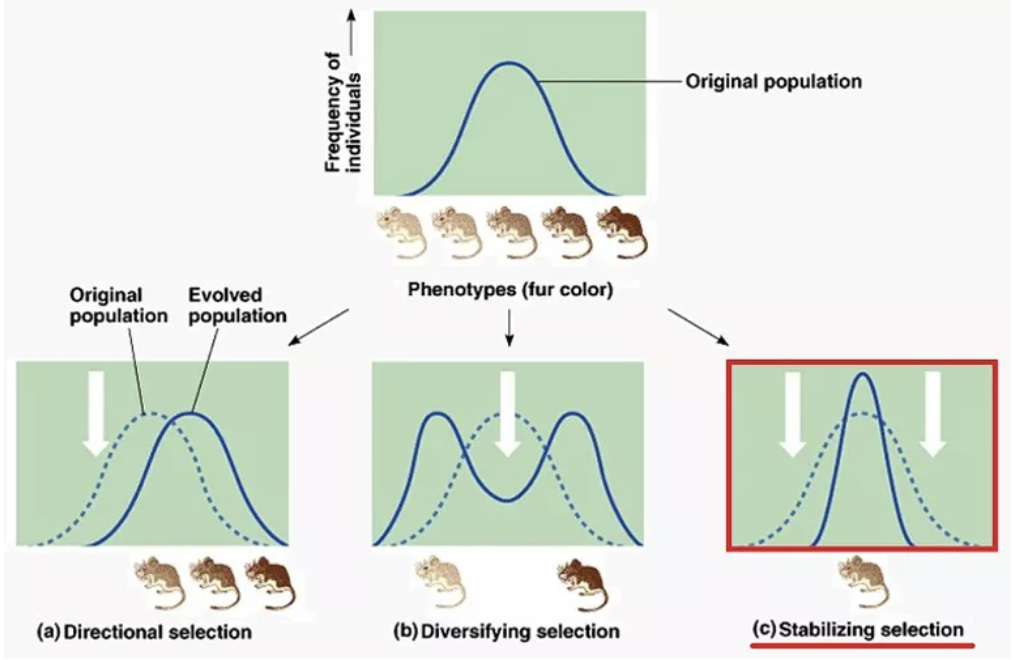
图片来源:wareslab.genetics.uga.edu
定向选择导致种群向某一方向变异,歧化选择保留了种群中的极端变异,而稳定选择则保留中间类型的个体,使得生物类型保持相对的稳定。
以上三种概念都是在表型水平的,而我们今天讨论的平衡选择则是关于具体的位点的。在具体到位点水平时,可以分为正选择(positive selection)、纯化选择(purifying selection)和平衡选择(balancing selection);有时候把没有受到自然选择的情况称为中性选择(neutral selection)。
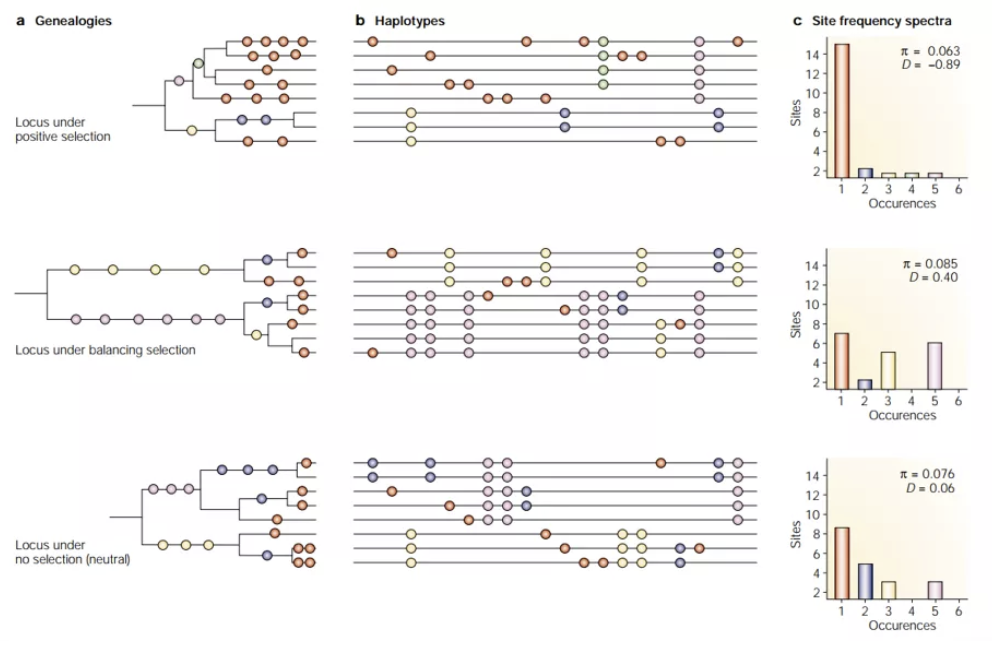
The effects of selection on the distribution of genetic variation (Bamshad M, et. Al., 2003)
如果用A表示野生型、a表示突变型,那么这几种自然选择表示如下:野生型的适合度最高时,为纯化选择;突变型的适合度最高时,为正选择;野生型位点与突变型位点杂合的个体适合度最高时,为平衡选择。从种群角度来看,受到平衡选择的位点能够长期维持多态性,例如导致人类镰状细胞贫血的基因位点。
前面讲的杂合子适合度最高,其实只是平衡选择的表现形式之一。如果将平衡选择的概念扩展一下,表示一种维持种群内位点多态性的力量,那么目前来看至少还包含另外两种机制。
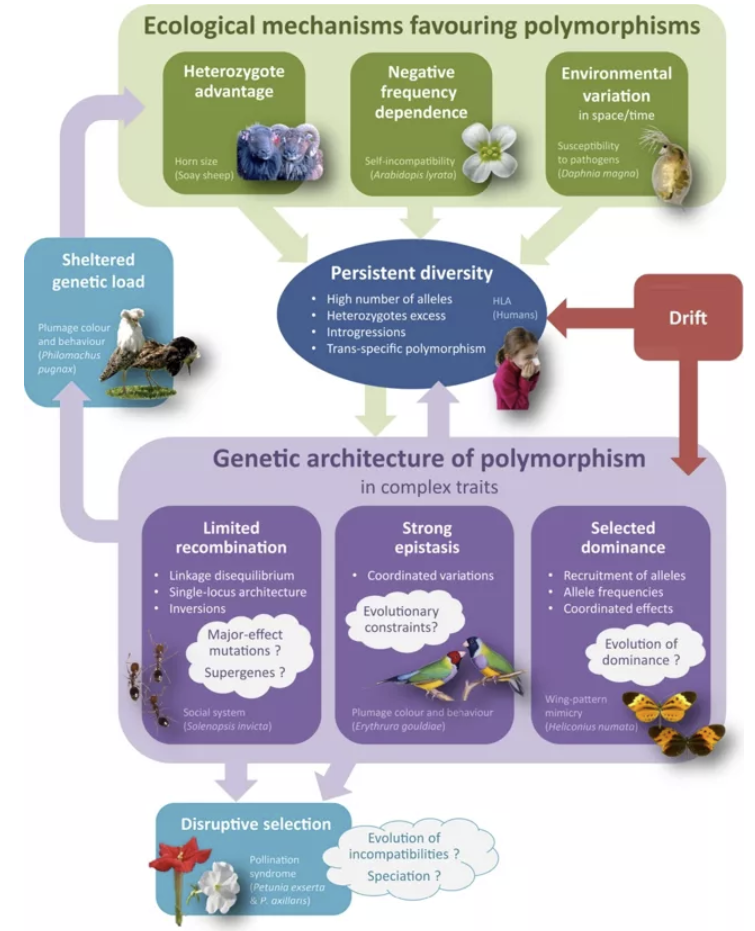
Balancing selection on complex traits (LlaurensV. et.al.,2017)
负频率依赖选择(negative frequency dependent selection, NFDS)是指,在群体中占少数的基因位点具有更高的适合度。例如有研究发现女性对男性胡须分布的偏好与人群中这种分布的比例呈负相关——胡子形状越稀少的男性,越受到青睐。
另一种情况是环境的波动,有时也可以称为波动选择(fluctuating selection, FS)。当生物所处的环境发生变化,最适应当前环境的基因型也随之变化,进而导致基因频率的此消彼长,从长期来看维持了种群内各种基因的多样性。
本期主要介绍的MHC II基因,是二类主要组织相容性复合体的缩写。
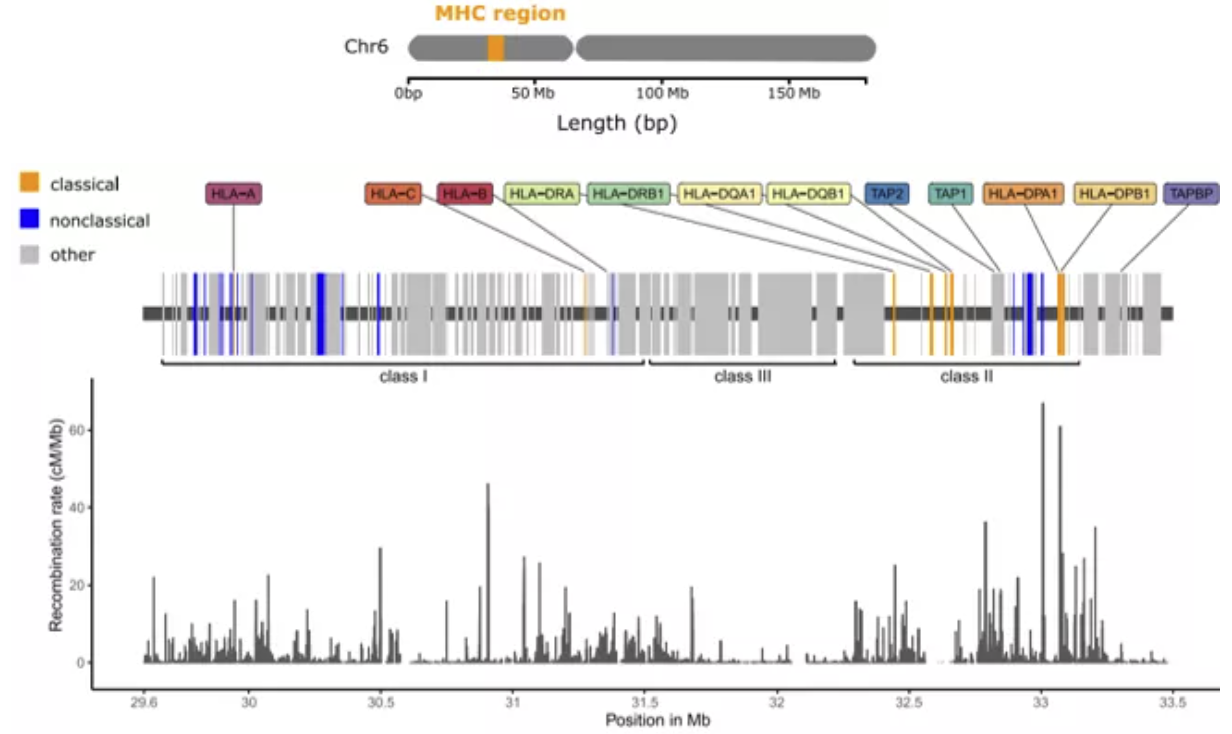
Genomic Map of Classical Human MHC (HLA) Region with Associated Recombination Rate (RadwanJ. et.al., 2020)
MHC II由多个紧密相邻的基因组成,这些基因的编码产物具有相同或相似的功能。此类基因主要参与免疫过程,在生物体抵抗病原体过程中发挥重要作用,不论在种间还是种内都具有极高的多态性。除此之外,它还受到强烈的正选择,并且具有跨物种多态性(trans-species polymorphism,TSP)。
那么,什么是TSP呢?
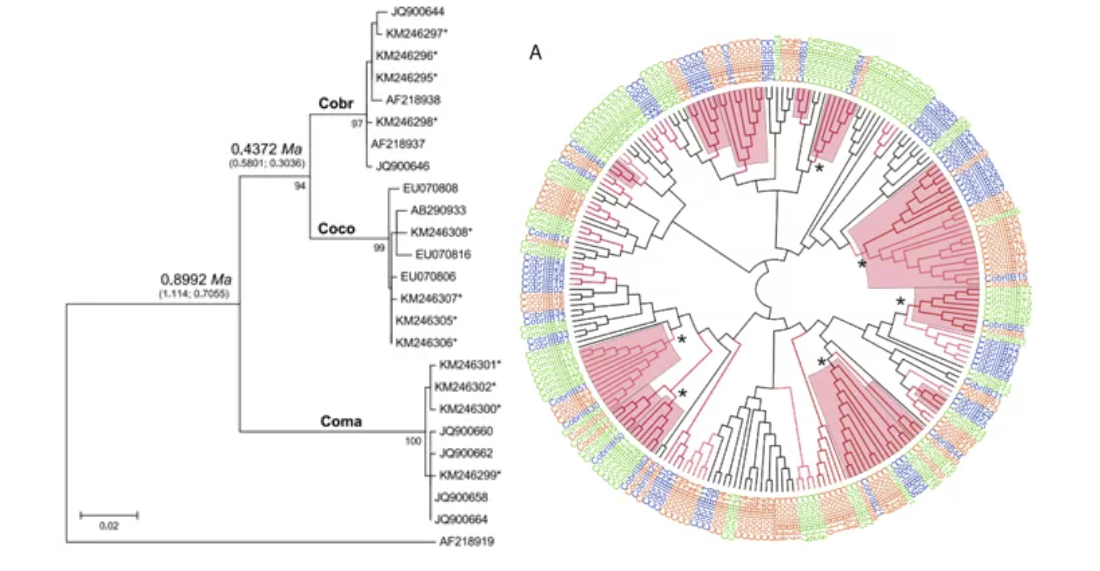
Patterns of evolution of MHC class II genes of crows suggest trans-species polymorphism (Eimes J. A. et.al., 2015)
这张图是对TSP的阐释。首先通过左边的线粒体控制区ML树可以看出,这三个乌鸦物种可以明显分开。右边展示了它们MHC II 某个区段的NJ树,可以看到不同物种的序列混杂在一起,甚至某些分支(*标出)同时包含了三个物种。也就是说,即使物种发生分化,此MHC II区段也没有发生分化,而是继续在不同物种中保留。
前面也提到过了,MHC II基因的另一个特征是即使在种内也具有高度的多态性。这是可以理解的,因为环境中的病原体复杂多样,MHC II必须维持较高的多样性才能抵抗复杂的病原组合。那么具体是哪些机制导致了这种多态性呢?回到刚才讲的平衡选择的三种机制,这里我们将它具象化为寄生虫对MHC II基因的平衡选择作用。

首先是杂合子优势(heterozygote advantage, HA)。MHC II基因参与寄生虫的识别和免疫过程,因此杂合MHC II基因的个体能够更广泛地识别环境中的寄生虫,因而具有优势。
另一种可能是负频率依赖选择(NFDS)——环境中的寄生虫也在进化,因此它们会提高对群体内频率最高免疫组成的抗性。这就像是一场军备竞赛,当宿主某一种免疫组成占优势之后,寄生虫产生了相应的抵抗对策,此时占少数的宿主反而不容易被寄生虫感染,进而占据优势。
第三种假说就是环境中寄生虫的组成因为其他因素发生变化,进而改变了对宿主的选择方向,即波动选择(FS)。下面的图将HA和FS放在了一起,这幅图的作者认为这两种机制是一个尺度问题——如果只看一代,那么平衡选择通过HA发生作用;如果在多代中暴露于不同的寄生虫,那么平衡选择就是通过FS发挥作用。
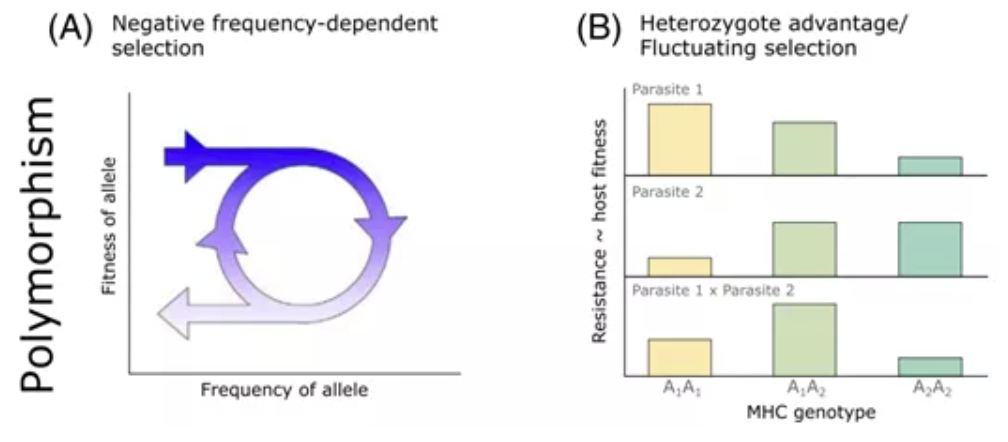
MHC polymorphism (RadwanJ. et.al., 2020)
寄生虫对MHC II的具体选择机制介绍完了,那么这种选择对群体产生了什么样的影响呢?
从理论上来看,平衡选择会降低不同种群之间的分化。因为这种选择压力倾向于维持多样性,而非让不同种群向着不同方向分化。但是在实证研究中,只有少数研究支持了这种假设。为此,有人提出是不同寄生虫组成导致不同的种群发生了不同方向的本地适应,进而选择出了不同的MHC基因。但是这种解释与前面提到的TSP发生了冲突——即使不同物种都可能保持相同的MHC,更何况是同一种内的不同种群呢?
于是人们又提出了一些其他的假设。
首先是,平衡选择作用的对象不是某个特定的位点,而是一组具有相似功能的位点——称为超类;另一种解释认为,跨物种多态性来源于种间的MHC渐渗,因此不能单纯从自然选择的角度考虑;还有人认为平衡选择(基于NFDS和FS)的空间异步性导致小尺度上种群明显分化,而大尺度上谱系分选减弱。
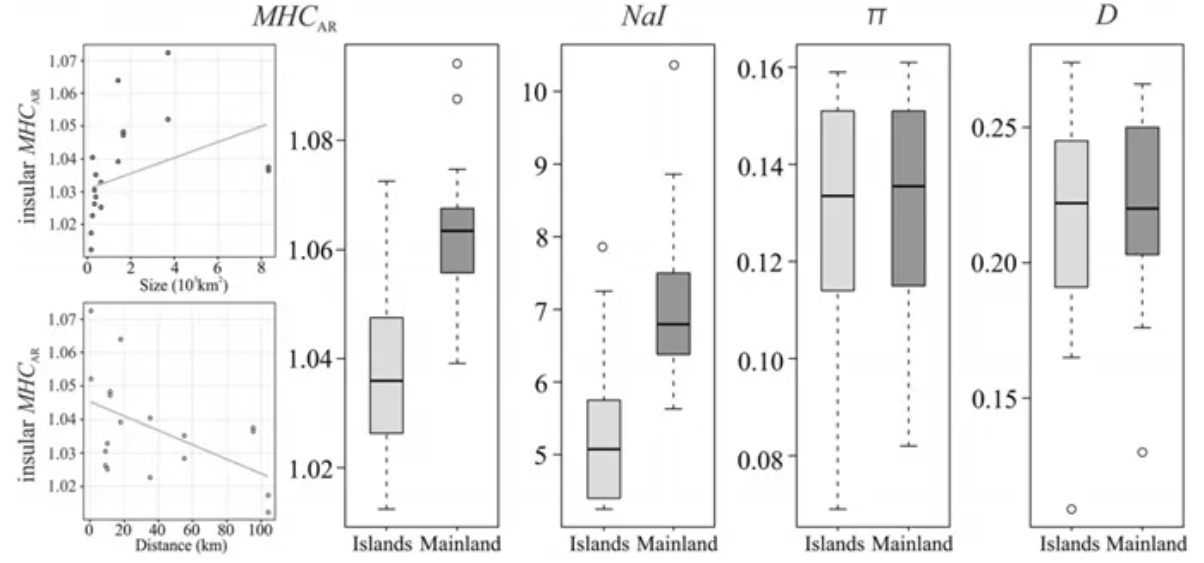
MHC diversity indices between mainland and island populations of Lacerta trilineata (SagonasK, et.al, 2019)
最后一个理论在一些实证研究中得到了支持,例如这一个关于蜥蜴MHC基因变异的研究中就发现,同一地区内的遗传分化高于岛屿之间。
但是自然界的历史通常是非常复杂、各种因素交织在一起的。如果能存在一个体系从一开始就隔绝某些因素的影响,就可能更有目的性地进行研究。
不同岛屿的湖泊鱼类就是一个非常好的模型,因为它们难以跨越地理隔离的障碍——如果海平面下降将它们隔离开了,那么它们就只能各自生活在自己的湖里,基本可以排除基因流的影响。

孔雀鱼 guppy 图片来源:www.ovas.ca
本文选取的研究体系,就是生活在Tri和Tob两个岛屿上的孔雀鱼,这两个岛屿在距今1-1.5万年前相互隔离开,在此之后可以认为两个种群之间完全没有基因交流。
Geographic distribution of sampledpopulations
本文主要想研究四个问题:
1、 MHC受到的是平衡选择还是歧化选择。这一点主要通过MHC II基因与微卫星位点的岛内-岛间分化来判断
2、 平衡选择是作用于超类还是位点
3、 分化的位点是否具有某种生存优势。一般认为,具有多态性的MHC位点具有更强的免疫力;所以它们是否更具有生存优势,可以通过种群内和单倍型内的MHC是否更多样来判断
4、 被隔离的两个岛屿是否共同保留了一些谱系的MHC基因。这一点主要通过MHC II的系统发育树进行判断,查看有哪些单系群在两个岛屿中都存在。
作者主要分析的数据包括MHC II基因和微卫星位点两类,主要的实验设计包括系统发育分析、种群结构分析,以及MHC位点的功能分化。需要提的一点是,孔雀鱼本身研究比较充分,已经有使用多个MHCII位点构建单倍型的方法。但这种方法需要养鱼和杂交,因此很难使用在其他非模式动物上。

这是作者在这篇文章中使用的方法,主要包括微卫星和MHC的分型、系统发育分析、种群遗传学分析,以及MHC位点功能分化的计算。这种功能分化,主要是通过比较当前位点与随机生成位点在氨基酸序列和物理化学参数空间中的差异性来进行区分的。如果当前位点有明显的适应性意义,那么就应该偏离随机生成的分布。
接下来来介绍一下文章的主要结果。
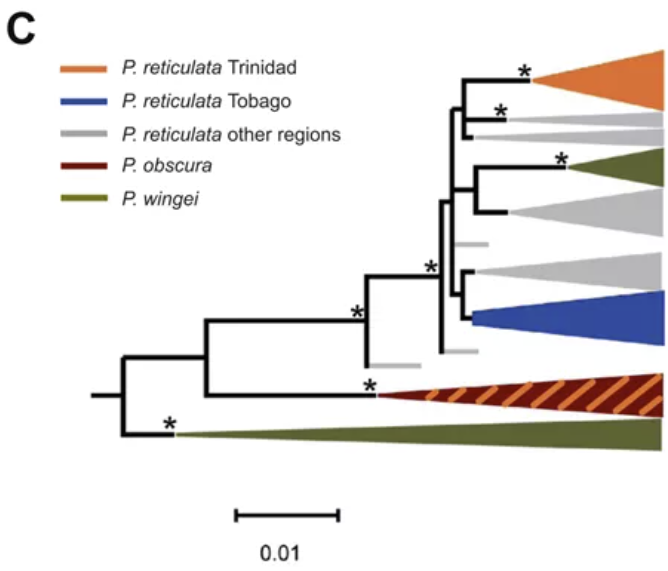
NJ tree showing relationships between mtDNA cytochrome b sequences (945 bp) from
P.obscura,P.reticulata and P.wingei,rooted with sequences from Micropoecilia picta and P.latipinna
使用线粒体cytb构建NJ树可以看出Tob的个体聚在一支,Tri的个体有两支,其中一支与另一个物种P.obscura嵌合在一起(橙色斜线)。Tob构成一个单系群,与其他孔雀鱼分开的遗传距离约为0.4%;假设线粒体的突变率为每百万年2%,那么Tob分化出来的时间约为距今20万年前。所以Tob和Tri分化的时间早于海平面下降、两个岛屿分隔开的时间。
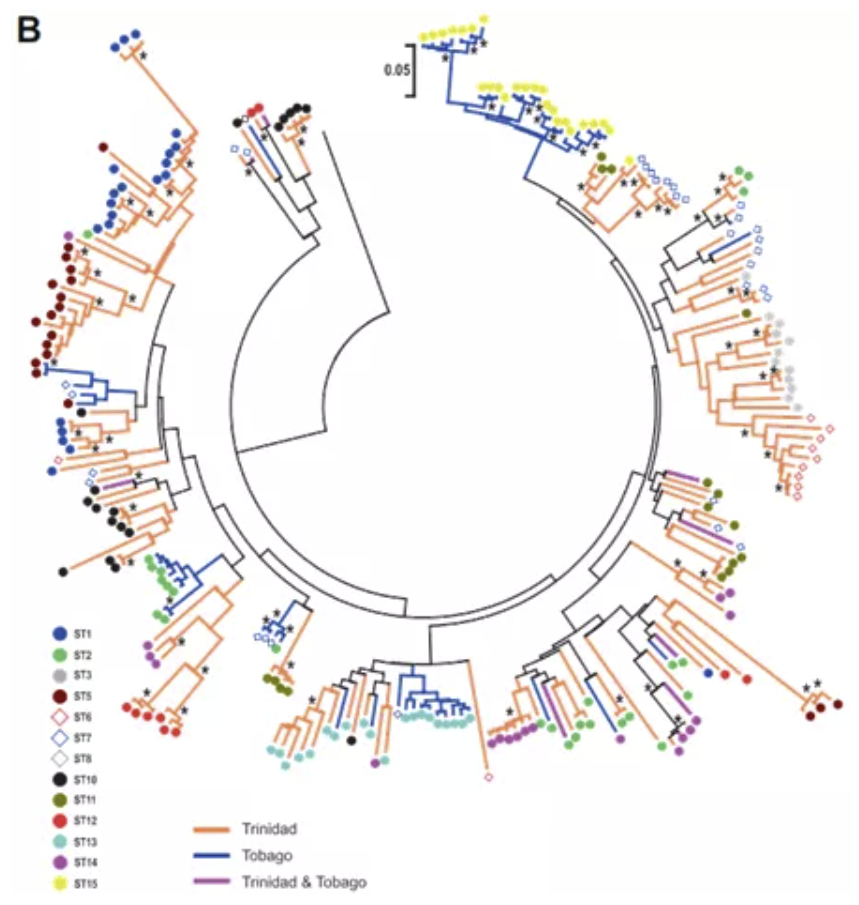
Neighbor Joining (NJ) tree showing relationships between MHC class II
上图是使用MHC II构建的NJ树。只有9个MHC超类在Tri和Tob中共有;支持率高的谱系大多都不是两个岛共有的——除了一个包含ST02和ST07的超类谱系;最大的岛屿特异位点包含23个只出现在Tob的位点,为ST15。
随后,作者分析了MHC和微卫星位点在种群中的变异情况。
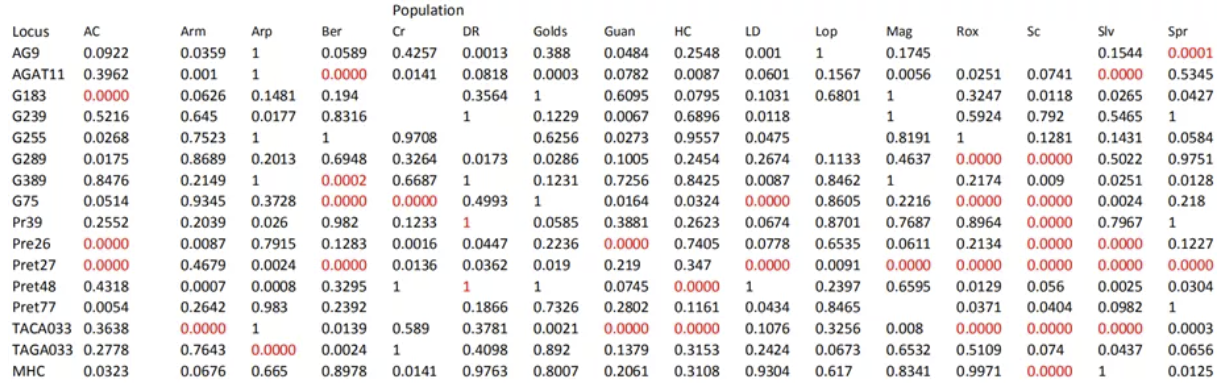
Results of Hardy-Weinberg equilibrium tests for each locus in each population
作者对微卫星数据和MHC数据进行了哈温平衡检测,发现有34个位点偏离了HW平衡,也包括一个MHC。Null allele和LD可能导致了偏离HW平衡,但这些现象只在少数种群中出现,而非全部种群,因此更有可能是漂变导致的,而非物理上临近。因此后续的分析中,作者没有去掉这些偏离HW平衡的位点。
Allelic richness of all mst loci and MHC haplotype richness in each population,
together with means perpopulation for microsatellite markers
作者发现,MHC的单倍型多样性比微卫星高3倍,且Tri的MHC和微卫星多样性都比Tob高。
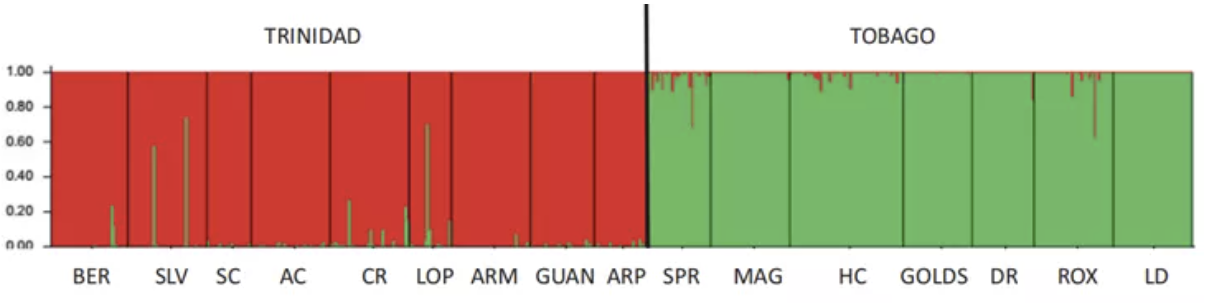
STRUCTURE plot based on 15 microsatellite loci with the optimal number of clusters k = 2
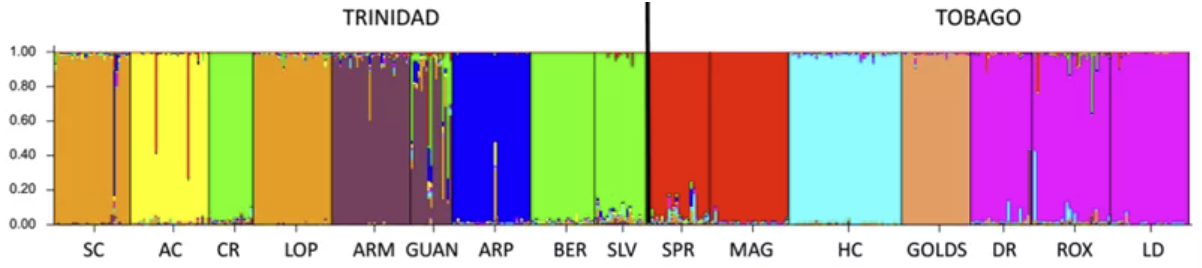
STRUCTURE plot based on 15 microsatellite loci with the optimal number of clusters K=9,
supported by Pritchard’s maximum probability method
作者使用STRUCTURE进行群体结构分析,采用了两种不同的方法对K值进行选取:使用Evanno的方法K=2,与岛屿划分一致;使用Pritchard的方法K=9,但没有跨越岛屿的界限,表明Tri和Tob还是有明显的分化。
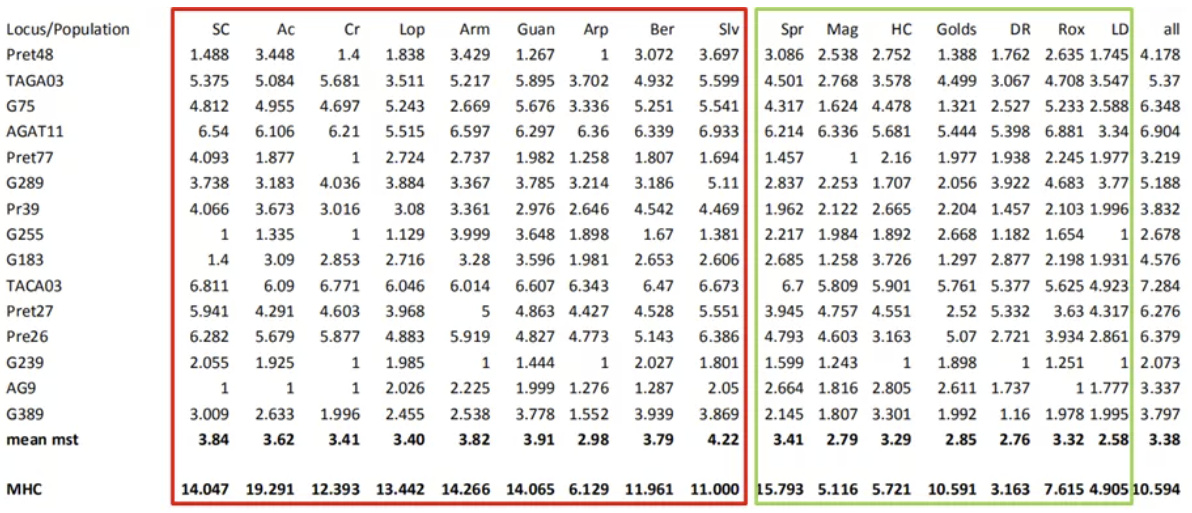
Gst是Nei提出的一种可以用于计算种群分化程度的指标,它与Wrigt提出的Fst相似,但可以用于计算复等位基因的遗传多样性。使用Gst计算种群对分化时,微卫星中Tri平均为0.41,Tob平均为0.33,岛屿间平均值为0.35;使用MHC时,Tri平均为0.19,Tob平均为0.27,岛屿间平均为0.22。
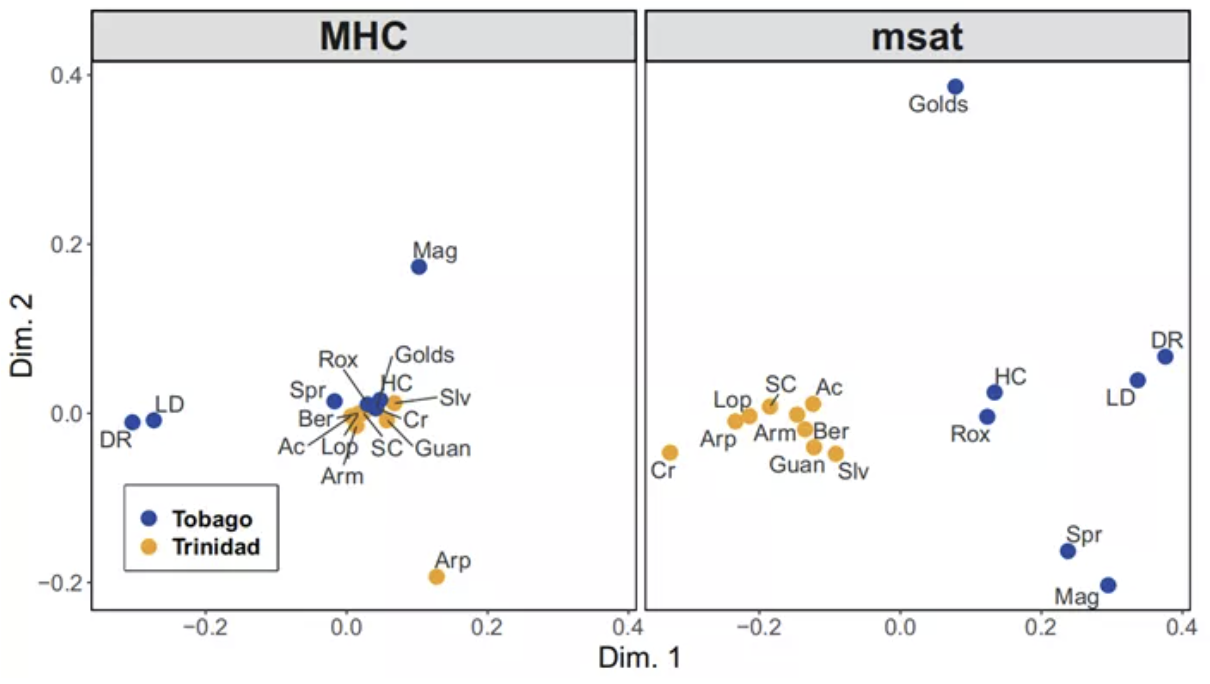
Two-dimensional scaling of the matrix of pairwise GST values between all sampled populations
based on MHC (left) and microsatellite (right) allele frequencies
作者对成对Gst矩阵进行二维尺度分析(可以理解为一种矩阵降维处理)。在微卫星数据中,dim1上两个岛屿明显分开;而在dim2上,Tob分开,Tri没有分开——这表明Tob上存在强烈的漂变。而MHC数据则明显不同,大多数的种群都聚在一起——表明它们的分化程度相差无几。
AMOVA(Analysis of Molecular Variance,分子方差分析)的结果也表明,微卫星的变异中,51.1%的为种群内水平,岛间占26.7%;而MHC的变异中,种群内占78.4%,岛间仅占2.3%。
以上结果表明,MHC在种群之间、岛屿之间的变异模式与微卫星位点不同。随后,作者使用了两种方法(分层Fst和贝叶斯)进行Fst离群点分析。

Fst outlier analysis on MHC and microsatellite loci, using Bayescan and Arlequin software
分层Fst离群点分析表明,MHC和微卫星没有明显差异;但贝叶斯分析却发现MHC的Fst明显较低,微卫星位点中有7个(TAGA033、AGAT11、TACA033、Pret77、Pre26、G239、AG9)明显偏离了中性假设。上图中,A是用全部位点分析得到的结果,B是去除了7个偏离中性的微卫星位点后得到的结果,可以看出去除之后MHC的Fst明显不同于其他微卫星位点。
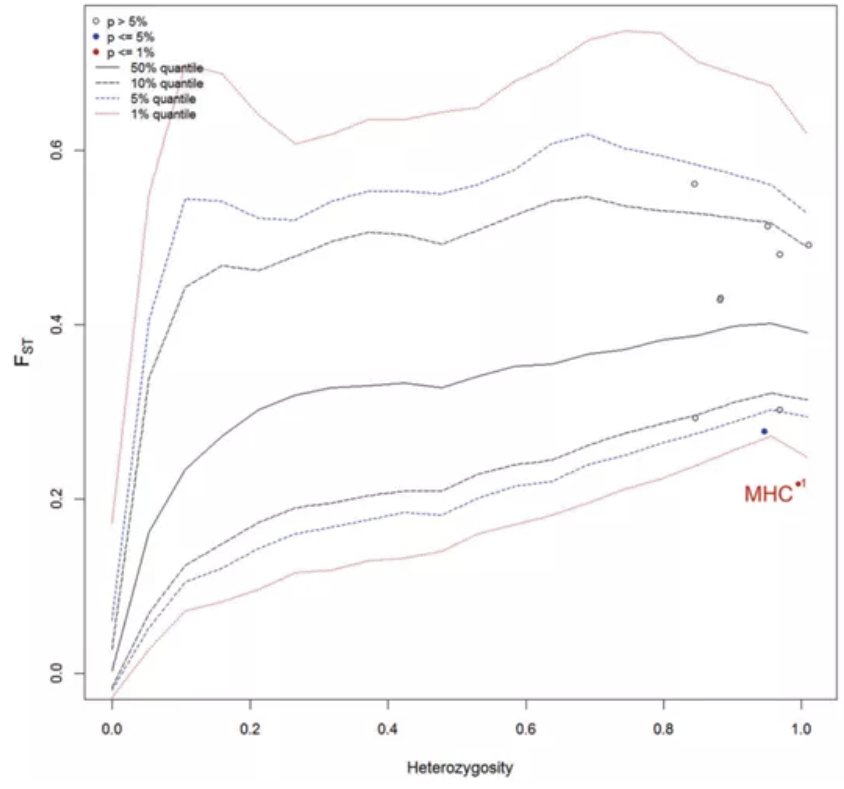
FST outlier analysis based on MHC andeight microsatellite loci
去除这7个微卫星位点后,两种方法得出的一致结果为MHC分化明显较低。
前文中提到过,大多数实证研究并不支持平衡选择降低了种群间的分化程度,为此有人提出选择压力并非作用于位点,而是作用于超类。为了对这种假说进行检验,作者将MHC II划分为了15个超类进行分析。
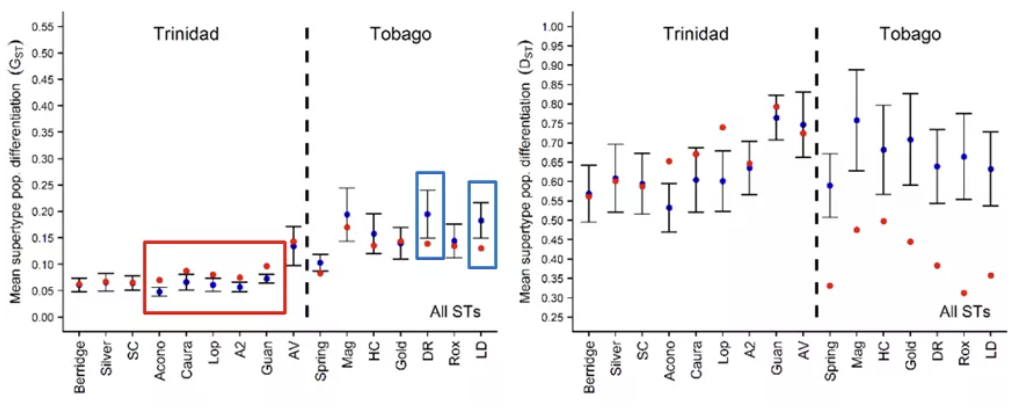
Pairwise MHC supertype-based population genetic differentiation
通过对同一岛内其他种群的Gst与估计值比较,作者发现,Tri有5个的观测值高于预测值(红框),Tob中则是有两个更低(蓝框)。如果更换评价分化程度的指标,改为Dst,则Tri有2个更高,Tob都普遍更低。这表明,作用于超类的选择压力在两个岛屿上是不同的。Tri中,选择压力提高了超类的分化情况,因此为歧化选择;Tob中选择压力降低了超类的分化,为平衡选择。
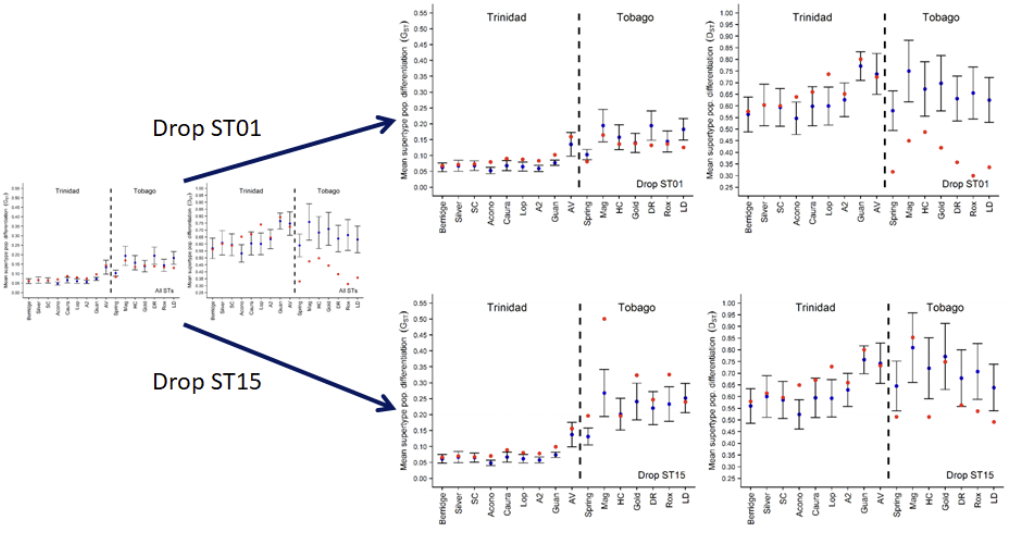
Pairwise MHC supertype-based population genetic differentiation
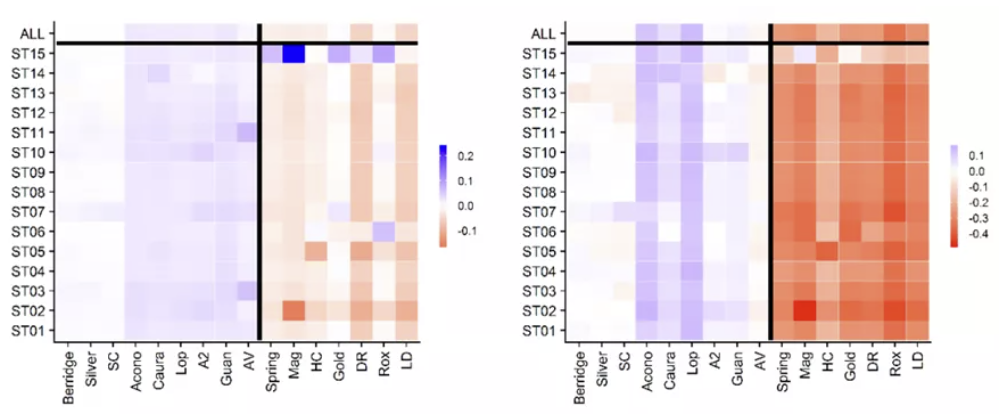
为了确定不同超类对分析结果的影响,作者分别将它们从数据集中去掉,观察Gst和Dst的变化,发现ST15对Tob的结论有重要影响。
另外还有假说认为种群间明显分化的MHC II位点具有明显的进化优势。但这种优势是很难衡量的,于是作者采用了一种计算模拟的方法:如果这些位点具有优势,那么它们的物理化学参数应该明显不同于随机产生的组合。
Distribution of physicochemical Euclidean distances between pairs of MHC alleles
但是结果表明,种群内分化的位点并没有比随机产生的分化更明显:受正选择的15个氨基酸位点的功能欧氏距离没有明显高于随机组合,因此分化位点并不具有明显的生存优势。

Per-population mean number ofprivate MHC alleles (Private alleles),
fraction of alleles that could be explained as derived by single nucleotide substitution (Local substitutions )
from or recombination between (Local chimaeras) other allele/s found in the same population
作者统计发现,这种结果可能是局部变异的影响。Tri的本地替换(单核苷酸替换)和嵌合(同一种群其他位点的重组)都很高,这表明存在很多局部地区产生的变异降低了种群内的功能欧氏距离。
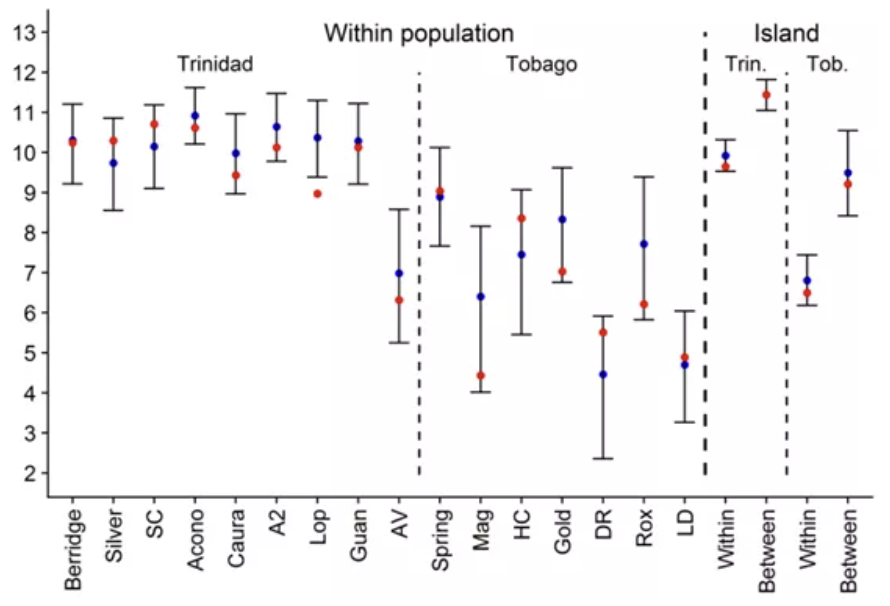
另一种检测MHC II是否具有优势的方法是观测同一单倍型中的变异情况。
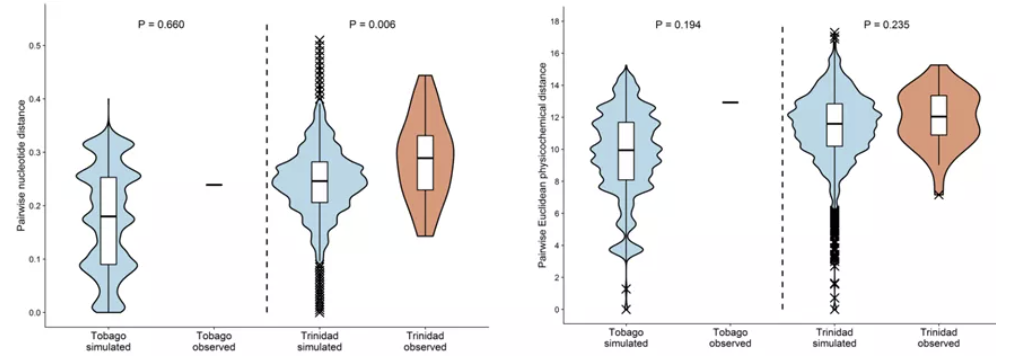
Comparison of the distributions of observed distances between pairs of MHC variants
segregating in the same haplotype to random pairs of variants inthe respective populations
作者发现同一单倍型内的核苷酸遗传距离在Tri中明显更高,而Tob则没有明显变化;但是对物理化学参数的统计发现,Tri中明显分化的单倍型没有更明显的功能分化。
接下来是文章的讨论部分,作者讨论了五部分内容。
一、岛内和岛间的分化。线粒体NJ树和微卫星种群结构分析的结果都表明,Tri和Tob在遗传上明显分化了。Tri的MHC单倍型多样性和微卫星多样性都高于Tob,这可能反应了两个岛屿种群历史的差异。Tob的面积比Tri更小、适合孔雀鱼生存的河流也少,而且作者的经验表明Tob河流中孔雀鱼的密度也低于Tri——这些因素导致了Tob的有效群体比Tri更小。
二、作用于MHC的选择压力。Fst离群点分析表明,MHC的空间变异比微卫星位点更低,揭示了孔雀鱼中MHC基因受到的是平衡选择,但Tob的信号较弱(在Fst outlier分析中,单独使用Tob无法得出MHC分化较低的结论)。Tri受到的平衡选择更强的原因可能包括:Tob受到漂变的影响更强,Tri的河流连通性更高、寄生虫选择压力更强,以及微卫星位点偏离中性假设。
三、分化位点是否具有优势。前言中提到的平衡选择三种机制中,有一种假设了新位点具有生存优势。但本研究表明,种群内分化的位点中没有发现分化位点适合度更高的证据,单倍型水平也是相似的结果。之前有研究表明,尽管新MHC可以在寄生虫压力下具有明显的优势,但这些新位点带来的功能变化并没有高于随机突变的组合。
四、作用于超类的选择压力。作用于MHC超类的选择压并没有强于位点上,而是呈现出一种复杂的模式:Tob表现为平衡选择,Tri表现为歧化选择。但这个结果依赖于对群体遗传学参数的选择,例如Tob的超类分化更浅是通过Dst得出的结论,而Gst则不支持这种结果。另外,ST15对Tob的遗传模式分析结果有很大的影响。
五、MHC谱系在岛屿中的保留情况。理论上,平衡选择会导致不同区域的分化水平低于区域内各个种群的水平。本研究也发现了岛屿水平MHC分化较低,但只有九个位点在岛屿中共享,因此作者认为这种分化较低可能是来源于大多数MHC变异来源于较低水平(种群内或同一岛内的不同种群)。除此以外,作者认为MHC的进化是一个动态的过程。首先,系统发育分析的结果中,大多数单系群都分布在某一岛上,因此MHC在岛屿隔离之后经历了快速的谱系分选;其次,ST15的系统发育位置表明,此超类在Tob快速扩散。
总结一下,这篇文章表明,孔雀鱼MHC II基因的进化受到平衡选择作用,这种选择作用与位点的优势无关,并且也并不更多地发生在超类。而且,MHC II基因进化是一个动态的过程,伴随着MHC单系的丢失、新位点的产生和扩散。
参考文献
[1] Herdegen-Radwan M, Phillips K P, Babik W, et al. Balancing selection versus allele and supertype turnover in MHC class II genes in guppies[J].Heredity, 2020: 1-13.
[2] Llaurens V, Whibley A, Joron M. Genetic architecture and balancing selection: the life and death of differentiated variants[J]. Molecular ecology,2017, 26(9): 2430-2448.
[3] Radwan J, Babik W, Kaufman J, et al. Advances in the evolutionary understanding of MHC polymorphism[J]. Trends in Genetics, 2020, 36(4): 298-311.
[4] Sagonas K, Runemark A, Antoniou A, et al. Selection, drift, and introgression shape MHC polymorphism in lizards[J]. Heredity, 2019, 122(4):468-484.
[5] Eimes J A, Townsend A K, Sepil I, et al. Patterns of evolution of MHC class II genes of crows (Corvus) suggest trans-species polymorphism[J].PeerJ, 2015, 3: e853.
[6] Bamshad M, Wooding SP. Signatures of natural selection in the human genome. Nat Rev Genet. 2003 Feb;4(2):99-111.
https://blog.sciencenet.cn/blog-3461164-1264496.html
上一篇:进化拯救:杂交渐渗使物种适应环境污染
下一篇:论稳定的邻居关系对北美红松鼠生存的重要性