博文
以工具为中心:神经元学说的三次转变  精选
精选
|
以工具为中心:神经元学说的三次转变
李双1,李侠2
(1.2. 上海交通大学 科学史与科学文化研究院,上海 200240)
摘 要:近年来,一些神经科学家提倡以“工具为中心”的神经科学发展新模式,但现有研究多局限于单一事件,缺乏对神经科学发展历程的系统性考察。梳理神经元学说的演进历程(1850-1950),可以发现神经元学说的三次重大理论革命都离不开技术层面的革新。首先,光学显微镜和组织学染色技术的进步解决了神经细胞结构的基本争议。其次,电压钳制技术的应用使霍奇金和赫胥黎得以纠正伯恩斯坦的膜假说,建立离子假说,从而揭示动作电位的机制。第三,玻璃微电极技术的发展促使埃克尔斯转向化学传递机制,并帮助卡兹最终阐明突触传递的本质。工具中心的视阈下,系统考察假说提出、仪器改进、争论终结和理论确证的完整过程,不仅能更全面地理解神经元学说的发展史,也为当代神经科学的发展提供了重要启示。
关键词:神经科学; 神经元学说; 工具; 科学思想史
中图分类号:N09 文献标识码:A
科学发现模式是科学理论研究的核心命题,对该问题的厘清意义非凡,影响着具体学科的发展方向、科研组织的构建、科学政策的制定,甚至是人类认识世界的方式,因而古今学者们从逻辑实证、史学例证、社会分析等角度切入,旨在廓清科学发展的路径。传统的观念里,科学的发展多以理论为中心。无论是逻辑实证主义者卡尔纳普(Rudolf Carnap)、批判理性主义者波普尔(Karl Popper),还是历史主义者库恩(Thomas Samuel Kuhn)和拉卡托斯(Imre Lakatos),他们都认为科学发展是以关键理论为节点,通过革命性新理论的出现实现跃迁。
然受哈金(Lan Hacking)科学实践观的影响,神经科学研究者开始关注技术工具的革新作用。该观点集中反映在《神经科学实验的工具——哲学和科学视野》(The tools of neuroscience experiment: philosophical and scientific perspectives)一书中,以比克尔(John Bickle)为代表的学者强调:理论往往产生于关键数据之后,科研更像是一场以改进科学设备为主的“游戏”,而非纯粹的理论探索。他们探究微电极、水迷宫、SCAPE显微镜、光遗传学等仪器对感官、记忆、注意力等意识研究的推进作用,主张“以工具为中心”的神经科学发展新模式。[1]这一视角富有新意,但现有研究多局限于个别案例分析,让人不免有“只见树木,不见树林”的遗憾。因此,本文将通过考察神经元学说的三次重大转变,系统揭示工具与理论在科学发展不同阶段的作用及其互动逻辑。
一 从网状理论到神经元学说:组织染色技术凸显神经细胞的独立性
1.1组织学染色法对光学显微镜局限性的突破
19世纪30年代,消色差显微镜的发明与传播为观察微观世界提供可能,但当时的显微镜技术仍无法清晰呈现大脑的内部结构,有时反而导致一些错误认识。利德尔(Liddell)深中肯綮:“显微镜是一剂良药,但就目前看来,其对生理学和解剖学的助益仍然有限。”[2]正因光学显微镜的局限性,神经科学研究者们转而寻求组织学方法上的突破,以获得更清晰的神经组织图像。
组织学染色技术的两次关键突破,极大推进了人们对神经细胞的认识。1858年,德国生理学家格拉赫(Joseph von Gerlach,1820-1896)在一次实验中偶然发现,将碳酸胺(carbamic acid)处理过的小脑组织浸泡在胭脂红(carmine)和明胶溶液(gelatin solution)中进行染色后,神经组织的结构变得更加清晰可见。[3]通过这一改进,格拉赫观察到神经细胞与神经纤维相连,这促使他提出神经网状假说。
然而,格拉赫染色法仅能呈现神经细胞模糊的局部图像。真正取得突破性进展的是1873年意大利医生高尔基(Camillo Golgi,1843-1926)创制的黑色反应染色法。他将硝酸银(silver nitrate)倒在重铬酸钾(potassium dichromate)硬化过的组织上,发现部分神经纤维被染为黑色,这使得神经细胞及其纤维以前所未有的清晰度显现。[4]在高尔基的观测下,神经纤维的末端呈蔓延状,据此,他提出“弥漫性神经网络”(Rete Nervosa Diffusa)假说。彼时尚无成像技术,所有观测结果都依赖研究者的手工绘制。高尔基凭借细致的观察绘制出大量结构清晰的神经细胞图谱,吸引了大批拥趸。
1887年,西班牙神经科学家卡哈尔(Santiago Ramón y Cajal,1852-1934)受高尔基启发,将黑色反应染色法改进为双重程序染色法(the procedure of double impregnation)。这一方法包含两个关键步骤:第一步,使用高尔基的银铬酸盐染色法,使神经组织结构在显微镜下变得清晰;第二步,脱水后,再次使用亚甲基蓝(azul metileno)、伊红(eosin)等染色剂进行二次染色,以增加细胞组织各成分的对比度。[5]经过卡哈尔的改良,染色效果更为稳定,也更易重复。
卡哈尔考虑到成熟的神经细胞可能难以被染色试剂完全渗透,因此转而开始研究新生神经细胞——幼年细胞未形成髓鞘,末端形态更清晰可见。[6]出人意料的是,卡哈尔发现新生细胞内的纤维并非如高尔基所描述的相互纠缠,而是彼此独立且有明确的终止点。卡哈尔遂得出两点与高尔基截然不同的结论。第一,神经细胞的树突和轴突末端均为自由终止。第二,轴突末端负责向具有导电性的细胞原生质传递神经冲动,而非提供营养物质。[7]这意味着神经组织并非由相互连接(吻合)的网状结构构成,而是由独立的单个细胞(即现今所称的神经元)组成。该结论几乎推翻网状理论的大多数观点,遭到以高尔基为代表的网状派的强烈反对。
1.2电子显微镜终结的神经细胞结构之争
1906年,诺贝尔生理学或医学奖颁奖典礼上,作为共同获奖者的高尔基和卡哈尔剑拔弩张。诺贝尔奖的演讲中,高尔基分别对卡哈尔神经元学说的两点关键性证据提出反驳,并抽丝剥茧地指出卡哈尔论证中的两点漏洞。其一,卡哈尔观察到的局部神经结构不足推及为神经细胞的普遍特征;其二,胚胎神经系统处于动态发展过程中,初期状态不足以用来推断成熟状态。高尔基以“马尔皮基氏层的上皮细胞”为例,指出卡哈尔看似合理的假说可能在若干年后被证明是完全错误的。[8]
有趣的是,高尔基从始至终都未否定卡哈尔的方法和观察证据,相反,他毫不吝啬地夸赞卡哈尔对其方法的改进,也认可卡哈尔通过该方法所获得的样本。高尔基的态度不仅反映了他对自创方法的自信,更体现当时研究者的实证主义精神——他们深信通过仪器和严谨方法获得的观察结果。正如高尔基反复强调的,他的科学立场完全“基于反复验证”、“基于解剖学的方法”,他主张弥漫性网络理论“不仅是理论,更是解剖事实”。[8]190-217
面对高尔基的质疑,卡哈尔并未当场反驳,而是返回酒店接受采访时才作出回应。卡哈尔首先赞美高尔基的染色法,称正是该方法燃起了他对神经科学的热情,使他在1888年后取得一系列重要的成果。[9]卡哈尔的自传中也提及该点,他写道:“当充分习得解剖学研究方法后,我以一种超乎寻常的热情投入神经系统的研究,高尔基的方法也开始在我的手中卓有成效”[6]324但卡哈尔认为他的成功主要得益于他对高尔基染色法的改进,及其思想和策略上的变化:转向新生神经细胞的研究。卡哈尔抱怨道,高尔基的科学偏见是如此之强,以至于其忽略了新生神经细胞如此重要的研究对象。
这表明,网状理论和神经元学说的支持者在研究方法和技术层面并无根本分歧,核心的矛盾在于对证据的解释不同。笔者认为,高尔基并非如后人所评价的那般保守固执[9],相反,他敏锐地意识到当时技术的局限性,并预见未来技术的进步可能促成不同理论融合。卡哈尔对高尔基的抱怨亦透露出惺惺相惜:“他有权选择他的研究主题,但不幸的是,他没有强调自己发现的宝贵事实,而是重新提出几乎被遗忘的弥漫神经网络理论……多么残酷的命运啊,就像连体婴儿一样,将截然不同的科学对手配对在一起!”[10]然而,高尔基直至1926年去世,都未接受卡哈尔的理论。
电子显微镜的发明为这场争论拉下帷幕。1954年,帕拉德(George Palade,1912-2008)和帕利(Sanford Palay,1918-2002)首次使用电子显微镜观察到神经元之间的结构,证实突触前膜和突触后膜之间存在突触间隙,这直接为卡哈尔的神经元学说提供直接证成。[11]
二 从膜假说到离子假说:电压钳制技术揭示动作电位的机制
2.1伯恩斯坦创制电流断续器提出膜假说
19世纪50年代,杜布瓦-雷蒙德(Du Bois—Reymond,1818-1896)和亥姆霍兹(Hermann von Helmholtz,1821-1894)初步观察到神经冲动时的电位变化,并发现在细胞内外存在显著的电压差,但他们当时尚未能对这些变化进行定量测定。正因如此,德国生理学家伯恩斯坦(Julius Bernstein,1839-1917)致力于研制一套能测量电位变化的仪器。
1868年,伯恩斯坦指出雷蒙德设备的两个主要缺陷:一是无法确定肌肉收缩时电位由正转负的具体时刻;二是无法测定负电位的实际变化情况。[12]为解决这些问题,伯恩斯坦考虑设计一种新型仪器。该仪器能够在特定时刻短暂闭合电路,并按固定时间间隔重复这一过程,通过观察电流变化来记录神经或肌肉的反应。[12]175为提高测量精度,伯恩斯坦还对仪器提出了几点关键性的要求:一是装置必须高速旋转以实现快速取样;二是需要引入独立的刺激控制机制;三是要尽量减少机械摩擦以降低测量误差。
基于以上的目标和设想,伯恩斯坦研制出电流断续器(Rheotome),其核心原理是利用快速的周期性取样来测量瞬时电位变化。该装置高13厘米、长26.5厘米,由左右两个功能单元组成。左侧单元负责控制电流刺激,通过中轴轮的匀速转动,使钢针周期性地接触铜导线,从而以固定时间间隔对青蛙肌肉施加电刺激。右侧单元负责电位测量,配备两个旋转黄铜臂,各带一个钢制槽型水银槽,钢针在旋转过程中依次接触水银面,与肌肉相连,记录神经受刺激后的瞬时电位变化,从而实现对神经冲动的定量测定。[12]176-177
电流刺激神经会产生负变化,但究竟何为负变化的直接导因?1889年,伯恩斯坦提出“膜假说”的猜想。伯恩斯坦推测,没有生物不浸润在液体中,因而所有的电流变化都可能受到某种化学物质的影响。在奥斯特瓦尔德(Friedrich Wilhelm Ostwald,1853-1932)半透膜实验的启发下,伯恩斯坦再次自制仪器,观察到青蛙肌肉的电位变化符合能斯特方程(Nernst equation)的预测,这表明电位变化源于电解质的流动。[13]据此,伯恩斯坦尝试提出假说:“纤维内部带负电,外部带正电。这种电双层在未受损的纤维中始终存在,但只有在损伤或受到刺激时才会表现出来(负变化)……为了简明起见,我将称其为 ‘膜理论’。”[13]542
然而,伯恩斯坦错误地将负变化完全归因于钾离子活动的影响。在缺乏精确测量手段的情况下,伯恩斯坦只能基于肌肉富含钾盐这一已知现象来推测其背后的机制。[14]
2.2霍奇金、赫胥黎运用电压钳制技术提出离子假说
1939年,霍奇金(Alan Hodgkin,1914-1998)和赫胥黎(Andrew Huxley,1917-2012)发现“超调”(Overshoot)现象,表明神经兴奋时膜电位不仅消失,还会短暂地变为正值,这与伯恩斯坦假说的预测不符。这一发现极大推动了膜电生理学的发展。
霍奇金和赫胥黎的成功得益于他们对电极技术的巧妙运用。1939年,他们在普利茅斯(Plymouth)海洋实验室对乌贼(Loligo)进行细胞内电极实验,发现动作电位峰值可超过静息电位约40毫伏,这表明神经兴奋时膜电位不仅不是降至零,反而会发生极性反转。[15]1939年12月,他们在《自然》(Nature)杂志上以简报形式发表了这一颠覆性发现,并强调成功的关键得益于杨(J. Z. Young,1907-1997)此前发现的乌贼巨型轴突(giant axon)——其直径约1毫米,是青蛙轴突直径的近100倍。[16]
电压钳制(voltage clamp)技术的应用使霍奇金等人能够精确控制膜电位并测量相应的离子电流。1945年,霍奇金及其同事发现钠和钾离子都会引起离子通道的开放,说明钠对膜电位也发挥着重要作用。1946至1948年间,霍奇金、赫胥黎和卡兹(Bernhard Katz,1911-2003)推测钠离子内流导致动作电位上升,而钾离子外流则使其恢复静息电位。然而,测量变化如此迅速的动作电位,难度极大,因而他们不得不暂时放下这一设想,直到电压钳制技术的发明。
霍奇金和赫胥黎发明的电压钳制技术的原理是:通过注入电流来抵消跨膜离子电流,从而实现对离子电流的间接测量。具体装置包括内外电极、反馈放大器和保护系统。其中,内外电极用于测量跨膜电位差;反馈放大器用于维持恒定的膜电位并测量补偿电流;保护系统为轴突提供屏蔽环境,确保测量准确性。[17]工作时,反馈放大器持续监测电压测量电极(b)和外部参考电极(c)之间的电位差,当膜电位偏离预设值时,放大器通过电流注入电极(a)输入补偿电流,以维持预设的膜电位。
1948年到1951年间,霍奇金和赫胥黎运用电压钳制技术,首次系统阐明动作电位的产生机制。如果静息电位(约-70毫伏)增加到约-55毫伏,钠通道开放导致钠离子内流,使膜电位迅速上升至约+50毫伏,达到峰值。随后钠通道失活,同时钾通道开放引起钾离子外流,使膜电位逐渐恢复至静息状态。他们大部分的数据于1948年末就已获得,但由于技术和时间的限制,相关研究成果直至20世纪50年代才陆续发表。霍奇金认为他们能够保留优先权的原因在于他们独特的方法未被他人发现:“幸运的是,没有其他人对电压钳制的分析特别感兴趣,我们能够慢慢来。”[18]
三 从电信号假说到化学递质假说:微电极技术阐明突触传递的本质
3.1埃克尔斯运用玻璃微电极证伪电信号假说
到20世纪30年代,神经元内部的电信号传导机制已基本明确,但神经元之间的信息传递方式仍存在争议。1936年,戴尔(Henry Dale,1875-1968)和勒维(Otto Loewi,1873-1961)因发现神经—肌肉接头处的化学传递而共同获得诺贝尔生理学或医学奖,但许多电生理学家仍坚持神经元间信息传递必须通过电信号完成。这场关于突触传递模式的争论在20世纪30年代达到顶峰,被形象地称为“汤派”(主张化学传递)与“火花派”(主张电信号传递)之争。
澳大利亚神经生理学家埃克尔斯(John Carew Eccles,1903-1997)起初是电传递理论的坚定支持者。1937年,他针对化学传递学说提出了两点主要质疑。其一,乙酰胆碱(acetylcholine)作为神经递质似乎作用时间过长。在迷走神经对心脏的作用中,传递者(乙酰胆碱)将会持续数秒,但神经节突触的递质必须在几毫秒内消失。其二,化学传递学说无法合理解释乙酰胆碱的快速清除机制。胆碱酯酶能催化乙酰胆碱的水解反应,使得乙酰胆碱被消除,从而终止其神经递质作用。但实验发现,即使毒扁豆碱(Eserine)抑制了胆碱酯酶(cholinesterases)活性,单个神经节前的齐放(volley)仍只引发短暂的“引爆器反应”(detonator response),突触反应的持续时间并未延长。这表明乙酰胆碱的清除可能存在其他机制。[19]
在考察完另一种解释突触传递的动作电流假说后,埃克尔斯承认两种假说都存在局限。他谨慎地指出:“目前对化学假说的研究较为深入,并获得了一些支持性证据,但我们必须正视它面临的严重困难,尤其是在解释已知的递质快速消失速率方面。在这些关键问题得到充分解决之前,这个假说显然不值得被普遍接受。”[19]553-554然而,即便面对大量支持化学递质的实验证据,埃克尔斯依然坚信电假说具有其合理性和解释空间。他援引好友波普尔的科学哲学观点,主张电信号假说具有更高的可证伪性,能提供更多被经验证据检验和反驳的机会。
1939年,埃克尔斯确定神经冲动在神经—肌肉接头处会产生两种兴奋作用。一种是持续时间不超过1毫秒的短暂兴奋作用所产生的“新生肌肉冲动”,类似于突触传递中的引爆器作用。另一种是独立于新生冲动产生的、持续时间更长的终板电位。经过对不同生物神经—肌肉接头的广泛测量,到1943年,埃克尔斯更加自信地得出结论:关于突触传递和易化所涉及的过程,特别是被称为“引爆器易化”的短暂现象,现有的大部分证据表明这些现象是由短暂的电递质作用引起的[20],这进一步支持了他的引爆器反应电假说。
然而,埃克尔斯在证伪过程中也发现了一系列反常证据。第一,箭毒(curarine)引起的突触阻断效应可以被化学传递假说很好地解释,但难以用电传递假说来阐明;第二,终板电位传导速度减慢的现象,可能源于抗胆碱酯酶(anti-Cholinesterases)对乙酰胆碱的抑制作用;第三,在箭毒化的哺乳动物神经元活动中,第二次突触前齐发产生的动作电位不仅没有减弱,在神经节中反而略有增强。尽管埃克尔斯试图用电假说来解释这些异常现象,但最终他不得不承认对化学假说和电假说都不甚满意。[21]
1951年8月,埃克尔斯及其团队进入最终的判决性实验阶段。他们采用了一种突破性的新方法——细胞内记录技术。[22]项该技术由格雷厄姆(Judith Graham)和杰拉德(R·W·Gerard)于1946年开创。埃克尔斯等人巧妙地将充满盐水的微电极(直径小于1微米)插入青蛙肌肉纤维中进行细胞膜穿刺记录,且不会干扰邻近的微电极。采用这种精密技术,是为了完成一场具有决定性的判决实验。通过检测正向或负向电位来验证电学模型与化学递质传递假说:若出现正向电位,则证实存在短暂的正向电场,支持电学模型;若出现负向电位,则验证了膜电位短暂升高的现象,支持化学递质传递假说。[23]这项实验的结果将最终裁定长期以来的理论之争!
针对猫的脊髓神经,埃克尔斯采取逐步推进的方式,每次移动几微米并间隔数秒,直至找到最佳记录位置。通过放大装置,他成功记录到单个神经元膜电位随时间的变化,且能持续观察长达2小时之久。[24]在观察动作电位记录时,埃克尔斯有了一个出人意料的发现:运动神经元的直接抑制现象与膜电位的短暂升高(表膜超极化)相关。这一现象令他惊讶不已:“电压确实下降了,这个结果不仅可以重复,而且会随刺激强度的增加而呈现分级变化,这是无可争议的事实。”[25]1952年,埃克尔斯携这一消息赴英,与之同行的,还有他作为化学递质研究者的“新手”热情!
关键证据的出现使埃克尔斯彻底改变了立场,称抑制和兴奋现象的唯一合理的解释就是化学突触传递假说。戴尔用一句富有戏剧性的比喻描述了埃克尔斯的这一转变:“突然间光芒四射,鳞片从他的眼睛里掉落。”[26]这个比喻源自《使徒行传》中保罗皈依基督教后重获光明的典故。然而,细究埃克尔斯的思想历程可以发现,他早在1946年就已产生了一种矛盾的心态,真正促使埃克尔斯皈依化学突触传递假说的是无数次的实验和玻璃微电极带来的无可辩驳的事实证据。
3.2卡兹用微电极技术和统计学方法解开化学递质传播之谜
卡兹早在1935年伦敦大学攻读博士期间,就亲眼目睹了埃克尔斯与戴尔等人在剑桥大学生理学会议上的学术争论,这给他留下了深刻的印象。[27]及至1950年春,神经肌肉传播的模式已基本确定:“神经冲动→乙酰胆碱→端板电位→肌肉冲动→收缩”,但“乙酰胆碱如何引起端板电位变化”的机制仍不明确。因此,卡兹和法特(Paul Fatts,1924-2004)利用细胞内电极检测青蛙终板电位,旨在为化学递质假说提供直接证据。
卡兹考虑到,“对终板电位的定量研究需要在离连接点的不同距离测试同一个纤维。此外,终板电位在活动时呈现多样性”,为了达到这一目的,“需要在肌肉纤维的不同点上连续插入电极,才能找到最大电位的位置。”[28]具体而言,他们将青蛙神经—缝匠肌标本安装在特制的观察室中,该室由石蜡制成并嵌入培养皿。标本置于透明有机玻璃块上,通过下方玻璃板提供照明。实验观察采用39倍放大的双目解剖显微镜。微电极(采用外接顶端直径在0.5微米以下的毛细管微电极并用锉刀锉削,充填3M氯化钾)通过橡胶管固定,部件置于微型机械手内。他们利用方形脉冲发生器对记录仪器进行时间常数测试,可随着微电极电阻的改变而调整,从而实现了对微电极不同部位电阻的测量。浴槽电极能够连续校准电压及脉冲。[28]
接下来,与霍奇金和赫胥黎的方法程序一致,卡兹和法特将微电极尖端从纤维表膜移动到纤维内部,记录电位下降约90毫伏,并用校准的电压输入补偿阴极射线测量这种下降。接着记录单一刺激的电反应(尖峰或终端电位),最后拔出电极,检查电位是否恢复到原水平。整个过程通常需要约15秒。[28]321
经过无数次实验,卡兹最终确定终端电位在3毫秒内达到峰值,并且乙酰胆碱在新斯的明(prostigmine)的刺激下反应显著。这些证据表明,端板(肌肉纤维的神经感受区)对化学刺激物特别敏感,而对电流缺乏敏感性。[28]362然而,卡兹不认为终端电位是由乙酰胆碱的离子直接引起的,因为乙酰胆碱的释放量不足以支持肌肉冲动。
尽管直接证明了乙酰胆碱假说并排除了一些错误假设,但卡兹仍未探明乙酰胆碱如何引发触发端板电位变化。直到1954年,卡兹与卡斯蒂略(J. Del Castillo)才正式提出化学突触传递的量子假说。[29]在探索青蛙的自发突触电位时,卡兹发现了“微型端板电位”(miniature e.p.p.),他推测这可能是端板反应的最小单位(量子)。无数个量子带来的微型电位结合起来,便可产生70-80毫伏的电子脉冲,这种量子变化是由乙酰胆碱使得肌肉膜短路所引发的。这一猜想有其合理性,但缺乏实质性的证据表明端板电位是由量子形式构成,因此有必要进行量化实验。
卡兹他们仍沿用细胞内电极的方法,用含不同浓度氯化钙(CaCl2)和氯化镁(MgCl2)组合的等渗溶液浸洗青蛙肢体,并统计端板电位的变化数据。结果发现,随着镁浓度升高或钙浓度下降,会产生非连续的电子脉冲,且波动越来越显著。卡兹用统计学方法验证了量子释放的基本假设。这种方法是重要的,如他所言:“显然,如果没有统计处理,这些结果就无法被分析,甚至无法获得令人满意的描述。”[29]563研究的结论是,在神经—肌肉连接处的传输由全有或全无的“量子”进行,其大小由同时发生的微型放电决定。量子单元的数量以随机方式波动,只能通过统计学进行预测。[29]571
英国药理学家大卫·科尔克霍恩(David Colquhoun)称赞卡兹是一位极具洞察力的科学家。在无数次看似枯燥的微型终板电位观测中,卡兹敏锐地注意到了一个关键现象:终板上那些微小而不起眼的自发电位波动并非实验记录的伪影,而是揭示突触传递量子化本质的重要线索。[30]化学递质量子假说的成功,离不开卡兹的个人努力,但这一假说的确立仍有赖于玻璃微电极、电子显微镜、统计学等技术方法的支持。
结 论
从“以工具为中心”的视角梳理神经元学说的三次历史转向,可初步得出三个结论。第一,工具促进对事物认识的广度和深度,并在不同学说之间扮演着裁判者的角色,它不自觉地充当了技术发展的助推器。第二,争论双方虽会存在分歧,但大多会认可对方的方法和工具。无论是高尔基和卡哈尔的争论,还是汤派和电火花派的争论,这其中所有的科学家都在具体的微观科技活动中扮演着实在论者的角色,这些基于实用主义的竞争无形中推动了科技的进步。第三,20世纪前,科学家更倾向独自创造、改进和完善技术手段,譬如高尔基、卡哈尔自创的组织学方法、伯恩斯坦制造的电流断续器;20世纪后,同一时期的研究者大多采用相似的工具和方法,工具的研发和实验的测试也逐渐分离开来,例如,霍奇金、赫胥黎、埃克尔斯和卡兹几乎都使用玻璃微电极技术。其原因可能在于大科学时代的到来,社会分工的逐步深化,工具制造日趋复杂化与昂贵化,从而使制造工具的费用大大超过了个人的承受能力。在大工业时代,工具的改进可以同时促成许多理论问题的解决。
至此,一阶结论明确:神经元学说研究离不开工具、方法和技术层面的进展。工具与方法的改进拓展了视野,提供丰富证据,化解理论纷争。然而,理论是使用工具和思考问题的先决条件,缺乏范式将无法从事常规的科学研究。继而可推导出两个二阶结论:一是如果理论完整(如物理学),工具不完整,则理论优先;二是如果理论不完整且工具改进容易(如神经生物学),则工具优先。这一认识对科技管理和政策制定具有重要的指导意义,有助于合理分配资源,优化研究策略,从而更有效地推动科学进步。
【参考文献】
[1] BICKLE J, CRAVER FC, BARWICH AS. The tools of neuroscience experiment: philosophical and scientific perspectives[M]. New York & London: Routledge, 2022.
[2] SHEPHERD GM. Foundations of the neuron doctrine: 25th anniversary edition[M]. New York: Oxford University Press, 2016: 22-23.
[3] GERLACH J. Mikroskopische studien aus dem gebiete der menschlichen morphologie[M]. Erlangen: Ferdinand Enke, 1858: 2.
[4] GOLGI C. Sulla struttura della sostanza grigia del cervello (Comunicazione preventiva)[J]. Gazzett Medica Italiana-Lombardia, 1873, 33: 244-246.
[5] CAJAL SRY. Estructura de los centros nerviosos de las aves[J]. Revista Trimestral de Histología Normal y Patológica, 1888, 1(1): 1-3.
[6] CAJAL SRY. Recollections of my life[M]. Cambridge: MIT Press, 1996: 324.
[7] CAJAL SRY. Texture of the nervous system of man and the vertebrates[M]. Vol 1. New York: Springer-Verlag Wien, 1999: 24.
[8] Golgi C. The neuron doctrine-theories and facts[A]//Nobel lectures, physiology or medicine 1901-1921[C]. Singapore: World Scientific Pub Co Inc, 1999:192.
[9] FERRARO A. Camillo Golgi (1843-1926)[A]//HAYMAKER W, SCHILLER F. Founders of neurology[C]. Springfield: Charles C Thomas, 1970: 35-39.
[10] JONES EG. The impossible interview with the man of the neuron doctrine[J]. Journal of the History of the Neurosciences, 2006, 15(4): 336.
[11] DE ROBERTIS ED, BENNETT H S. Some features of the submicroscopic morphology of synapses in frog and earthworm[J]. Journal of Cell Biology, 1955, 1(1): 47-58.
[12] BERNSTEIN J. Ueber den zeitlichen verlauf der negativen schwankung des nervenstroms[J]. Archiv für die Gesamte Physiologie des Menschen und der Tiere, 1868, 1(1): 174.
[13] BERNSTEIN J. Untersuchungen zur thermodynamik der bioelektrischen ströme[J]. Archiv für die Gesamte Physiologie des Menschen und der Tiere, 1902, 92(10): 531-533.
[14] BERNSTEIN J. Elektrobiologie[M]. Braunschweig: Vieweg & Sohn, 1912: 94.
[15] HODGKIN AL. Beginning: some reminiscences of my early life (1914-1947)[J]. Annual Review of Physiology, 1983, 45(1): 13.
[16] HODGKIN AL, HUXLEY AF. Action potentials recorded from inside a nerve fibre[J]. Nature, 1939, 144(3651): 710-711.
[17] HODGKIN AL, HUXLEY AF, KATZ B. Measurement of current-voltage relations in the membrane of the giant axon of loligo[J]. Journal of Physiology, 1952, 116(4): 424-448.
[18] HODGKIN AL. Sir Alan L. Hodgkin[A]//SQUIRE L R. The history of neuroscience in autobiography: Robert Galambos and Vernon Mountcastle[C]. Washington DC: Society for Neuroscience, 1996: 263.
[19] ECCLES J. Synaptic and neuro-muscular transmission[J]. Physiological Reviews, 1937, 17(4): 544-545.
[20] ECCLES J C. Synaptic potentials and transmission in sympathetic ganglion[J]. Journal of Physiology, 1943, 101(4): 465.
[21] ECCLES J C. An electrical hypothesis of synaptic and neuro-muscular transmission[J]. Annals of the New York Academy of Sciences, 1946, 46: 446-448.
[22] ECCLES J C. My living dialogue with Popper[A]//LEVINSON P. In pursuit of truth: essays on the philosophy of Karl Popper on the occasion of his 80th birthday[C]. Atlantic Highlands: Humanities Press, 1982: 224.
[23] ECCLES J C. From electrical to chemical transmission in the central nervous system: the closing address of the Sir Henry Dale centennial symposium[J]. Notes and Records of the Royal Society of London, 1976, 30(3): 219-230.
[24] BROCK LG, COOMBS J, ECCLES J. The recording of potentials from motoneurones with an intracellular electrode[J]. Journal of Physiology, 1952, 117(4): 432.
[25]ECCLES J C. From electrical to chemical transmission in the central nervous system:the closing address of the sir Henry Dale centennial symposium[J]. Notes and records of the Royal Society of London, 1976, 30(2):219-230.
[26] DALE H. The beginnings and the prospects of neurohumoral transmission[J]. Pharmacological Reviews, 1954, 6(1): 11.
[27] ORMEMRS BS. Bernard Katz: 26 March 1911-20 April 2003[J]. Biographical Memoirs of Fellows of the Royal Society, 2007, 53: 185-202.
[28] FATT P, KATZ B. An analysis of the end-plate potential recorded with an intra-cellular electrode[J]. Journal of Physiology, 1951, 115(3): 320-321.
[29] DEL CASTILLO J, KATZ B. Quantal components of the end-plate potential[J]. Journal of Physiology, 1954, 124(3): 560-573.
[30] COLQUHOUN D. Professor Sir Bernard Katz[J]. The Independent, 2003, 4: 19.
【基金项目】教育部人文社会科学重点研究基地项目:交叉科学引发的范式融合问题研究(22JJD720015)
【作者简介】李 双(1996-),女,湖南衡阳人,上海交通大学科学史与科学文化研究院博士生,研究方向为科学思想史、科学哲学;李侠(1967-),男,辽宁辽阳人,上海交通大学科学史与科学文化研究院教授,研究方向为科学哲学、科学政策与科学社会学。
【博主跋】李双来信告知这篇文章见刊了,发在《科学技术哲学研究》2025(2)。恍然感觉那仿佛是很久以前的事情了,现在一篇文章从构思、写作、修改再到最后的发表,如果顺利的话也得折腾一年到一年半左右,属实是折磨人的漫长旅途。作为科学思想史的主题,这篇文章探讨了一个很重要的问题;理论还是工具哪个在推动科学发展中作用更大?以波普尔为代表的传统哲学家们认为,理论更重要,但近年来一些学者认为,在某些学科工具中心论正在复兴。本文就是通过神经元理论的演变来探讨这个问题。如今,李双博士已经毕业,很欣赏她对学术的由衷热爱,以及对目标的坚定执行力,祝她在未来百尺竿头更进一步,前程似锦。
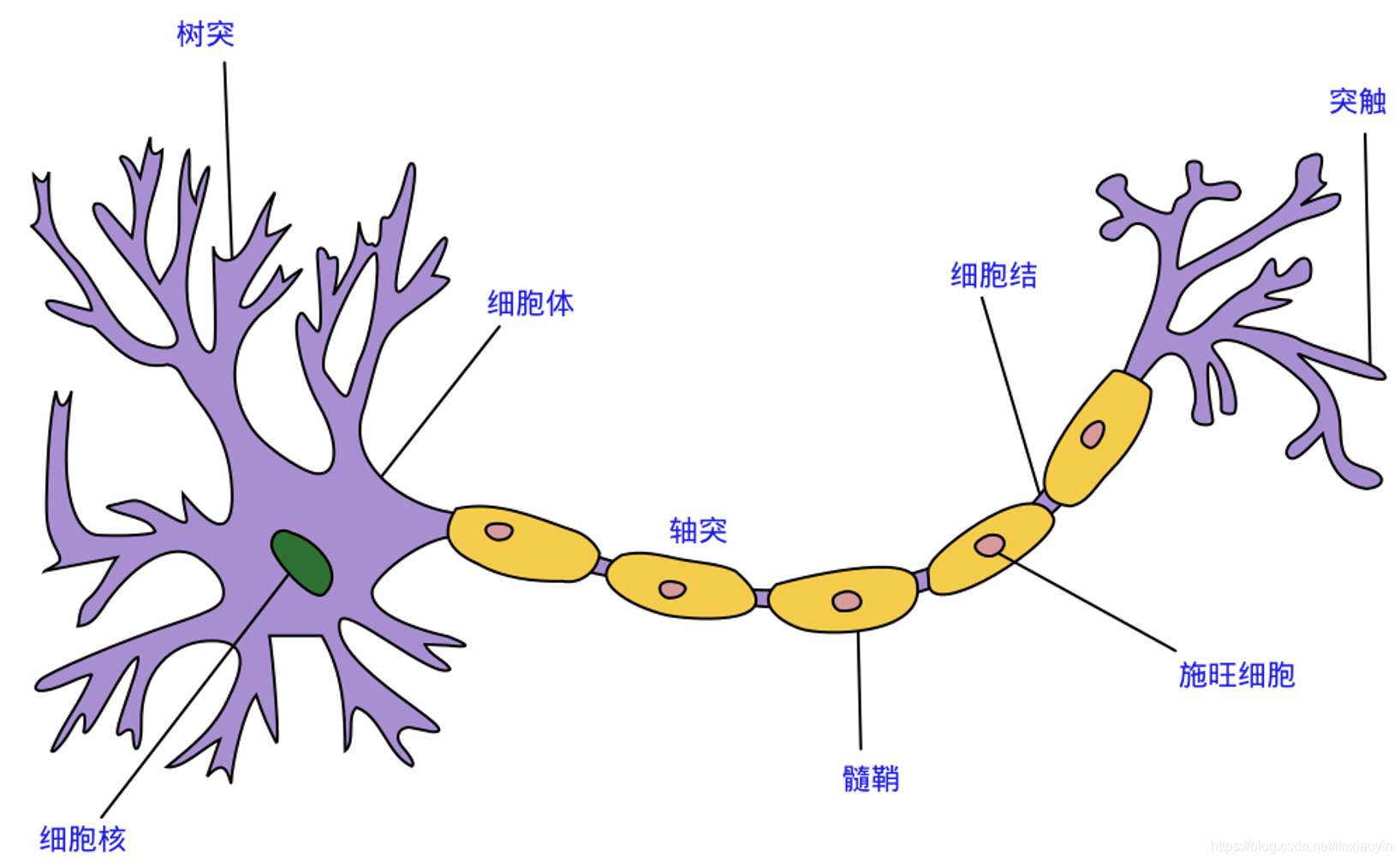
说明:文中神经元图片来自网络,没有任何商业目的,仅供欣赏,特此致谢!
2025-4-4凌晨于南方临屏涂鸦
https://blog.sciencenet.cn/blog-829-1480602.html
上一篇:执著与美丽:多萝西·克劳福特·霍奇金的晶体人生
下一篇:镜像与契机:中国应捍卫科研生态稳定乘势而上
