博文
氢气的生理效应研究更重要
||
由于是预先进行氢气处理,本研究所观察到的现象更符合“生理学”效应,因为这些效应必需通过生物体相应发挥作用。这是本研究的重要贡献。自噬是一种应对外界应激的积极反应,有利于度过不良外界环境。本研究发现氢气能加强这种效应。提示氢气具有类应激效应,或者类似于外界不良刺激的作用。因此可推测,氢气具有类似应激效应分子的地位。补充氢气或预先进行氢气处理,对植物来说就具有类似胁迫的处理。本例中渗透的作用和氢气作用具有类似性。而生物体对胁迫的反应具有病理生理学或生理学地位。为理解氢气生理学作用细节,胁迫导致内源性氢气产生的分子过程也就显得极其重要。
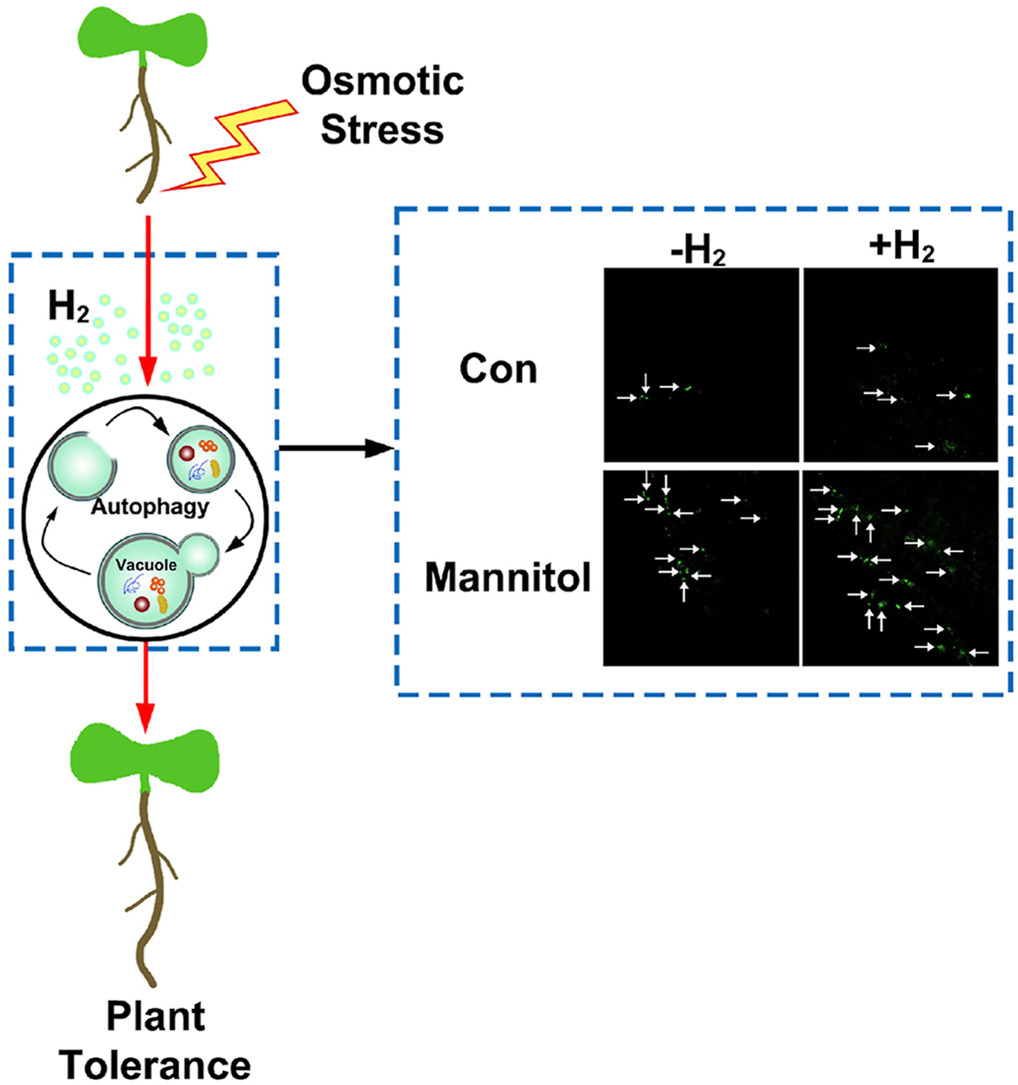
氢气熏蒸有助于植物抵抗渗透胁迫。
渗透胁迫诱导的自噬在氢气作用下得到加强,其中ATG基因作为下游靶标发挥作用。
内源性氢气介导的自噬在促进拟南芥的渗透耐受性中扮演重要角色。
氢气熏蒸和转基因植物是探索氢气在植物中作用更可靠的方法。
分子氢(H2)在哺乳动物细胞炎症反应中的自噬作用存在争议。尽管观察到植物响应渗透胁迫时产生氢气的刺激,但其合成途径及其诱导与植物自噬之间的相互关系尚不清楚。在这里,通过评估自噬体形成和自噬相关基因表达,观察到拟南芥在渗透胁迫下的自噬诱导。上述反应在氢气熏蒸下得到加强。同时,幼苗生长和根活力的减少明显被消除,伴随着氧化还原平衡的重新建立。这些氢气反应在T-DNA敲除株系atg2、atg5和atg18中明显受损。进一步的证据表明,通过遗传操作增加内源性氢气合成,不仅刺激了自噬体形成,还触发了植物对渗透胁迫的各种反应。相比之下,通过加入2,6-二氯靛酚钠盐破坏内源性氢气合成,这些反应明显被废除。综合的遗传和分子证据清晰地说明了自噬激活在氢气控制植物渗透耐受性中的必要性。
引言
由干旱和盐分引起的渗透胁迫,可以限制水分利用以及植物的生长和发育,成为限制作物生产力并威胁食品安全的关键环境因素[1], [2], [3]。这种胁迫不仅导致酶功能失调和膜损伤,还由于植物组织中活性氧种(ROS)的过度积累而导致细胞的氧化损伤[4]。
自噬在词源上意味着“自我吞噬”,是一种进化保守的过程,可以通过回收不必要的胞浆成分(包括过量蛋白质和受损的细胞器)到溶酶体或液泡中,从而密切调节细胞稳态的催化细胞机制[5], [6], [7]。通常,自噬与人类疾病和生理学有关,如神经退行性疾病、癌症、微生物感染和衰老[8]。植物中自噬的生物发生和功能已经得到了很好的阐述,包括几种不同的类型,即大自噬、小自噬和巨自噬。在这些类型中,大自噬,这里简称为“自噬”,涉及生成细胞质自噬体,吞噬不必要的胞浆成分[9], [10]。
在自噬诱导过程中,一个双膜结构,称为自噬体,吞噬胞浆成分并将其传递给液泡以进一步降解。自噬体与液泡对接后,外膜与液泡融合,内部小泡释放进入液泡腔。内部小泡(剩余的单层膜结构)被称为自噬体,参与液泡中的降解和回收[5], [11]。过去二十年中,已经在植物中鉴定出30多个自噬(ATG)相关基因。这些基因参与了自噬的诱导、延长、成熟以及与液泡的融合[6], [12], [13]。在植物中,自噬在特定发育阶段、营养回收、生物和非生物胁迫反应中有多重角色[14], [15], [16], [17]。
分子氢(H2)之前被认为是一种生物学上惰性的气体。然而,这一传统观点被日本科学家Ohsawa等人的一项里程碑报告所挑战[18]。他们观察到H2可能是一个候选的治疗性抗氧化剂,因为它能选择性地清除过氧亚硝酸盐和羟基自由基,从而缓解动物细胞中的缺血-再灌注损伤。进一步的研究发现H2在几种人类疾病中的有益功能,包括缺血-再灌注损伤[19]、败血症[20]、急性呼吸窘迫综合征[21]和动脉粥样硬化[22]。尽管对H2依赖的自噬在动物疾病和损伤反应中的分子理解日益增长,但其调控机制和功能仍然存在争议。例如,H2通过激活自噬被观察到可以减轻败血症[20]、动脉粥样硬化[22]和缺血-再灌注损伤[23]。然而,之前的研究也报告说H2可能通过抑制自噬来减轻创伤性脑损伤[24]。
在植物中,大量证据揭示H2的产生在响应环境刺激时显著增加,包括水分胁迫[25], [26]和盐分[27]。进一步显示外源性H2可能涉及多种植物生理反应,范围从根器官发生[28]到对抗环境刺激的保护,包括干旱[25]、盐分[27]、冷胁迫[29]和重金属暴露[30]。
氢气(H2)的代谢已经在原核生物和真核生物中观察到[31]。氢化酶是微生物和藻类中催化H2产生的重要模式,其催化过程依赖于光合作用系统[32]。尽管植物中尚未鉴定出氢化酶基因,但2,6-二氯靛酚钠盐(DCPIP)被认为是内源性H2合成的候选抑制剂,暗示其生产与光化学反应有关[33]。随后的结果证实,通过添加DCPIP,番茄[34]和拟南芥[26]中的H2产量显著受损。重要的是,由于H2的合成途径尚未阐明,使用转基因方法评估植物中内源性H2的作用是有效的,在这种方法中,拟南芥植物被转化了来自衣藻(Chlamydomonas reinhardtii)的氢化酶基因(CrHYD1),用于H2生物合成[26], [27], [35]。
在这项研究中,我们旨在调查内源性H2在渗透耐受性中的可能机制。我们的遗传和分子证据表明,H2激活的自噬对拟南芥的渗透耐受性至关重要。形态学和生化分析显示,通过H2熏蒸,拟南芥幼苗的渗透耐受性得到增强,这通过减轻主根生长抑制和加强自噬体形成来评估。重要的是,自噬参与了H2实现的这一反应。遗传和分子证据进一步证实,内源性H2可以通过正向调节拟南芥幼苗中的自噬来缓解渗透胁迫。因此,这项研究提高了对氢生物学以及H2在自噬调控中作用的理解。
植物材料和生长条件
本研究中的拟南芥(Arabidopsis thaliana)品系均以哥伦比亚-0(Col-0)为背景。CrHYD1转基因品系和空载体(EV)植物先前已有描述[26]。表达GFP-ATG8a的植物先前已有描述[36]。T-DNA插入突变体atg2(SALK_006994C)、atg5(SALK_020601C)和atg18(SAIL_690_B08)从欧洲拟南芥库存中心(NASC,诺丁汉,英国)获得。为了确认纯合T-DNA敲除品系,从莲座叶中提取了基因组DNA。
H2熏蒸赋予渗透耐受性
在用250 mM甘露醇(模拟渗透胁迫)处理5天后,与对照组相比,拟南芥幼苗的主根伸长减少了39.6%(图2A和B)。值得注意的是,无论是否存在甘露醇胁迫,H2熏蒸都促进了主根伸长(特别是在仅用甘露醇处理的情况下高出了23.2%)。在新鲜重量和干重方面,H2处理也取得了类似的积极效果。
讨论
关于植物对抗渗透胁迫时自噬的调控,特别是形成和动态,目前的理解仍然有限。尽管之前的研究结果发现渗透胁迫刺激了内源性H2的产生[26],但尚无文献显示H2是否参与植物中的自噬。这里,药理学和遗传学方法的结合揭示了内源性H2触发自噬以减轻拟南芥中的甘露醇诱导的渗透胁迫。
结论
总的来说,我们揭示了H2在植物响应渗透胁迫中的一个特定机制,即更高的内源性H2激活自噬以缓解渗透胁迫引起的抑制和损害。由于尚不清楚H2是直接还是间接调控自噬,H2的合成途径及其在植物中可能的信号传导尚未完全确定。
https://blog.sciencenet.cn/blog-41174-1453460.html
上一篇:比较基因组分析提供了心肌调节的机制线索
下一篇:儿童轻度脑震荡会不会增加成年后犯罪可能性【丹麦研究】
