ВЉЮФ
ЧтЦјМѕЩйЦпЗњЭщгеЕМаТЩњЖљЦЄВуЩёОдЊЕђЭіЃЌЧтЦјгааЇХЈЖШ1-8%  ОЋбЁ
ОЋбЁ
||
ЧтЦјМѕЩйЦпЗњЭщгеЕМаТЩњЖљЦЄВуЩёОдЊЕђЭі
БОбаОПФПЕФЪЧЙлВьЧтЦјЮќШыЪЧЗёФмМѕЩйТщзэЕМжТЕФФдЩёОЫ№ЩЫЃЌЗЂЯж1-8%ЧтЦјЮќШы3аЁЪБФмМѕЩйТщзэвЉЕМжТЕФЩёОдЊЧАЬхЯИАћЕђЭіЃЌетРяЕФЧАЬхЯИАћЪєгкФмЗжЛЏЮЊЩёОдЊЕФзцЯИАћЃЌОЭЪЧЩёОИЩЯИАћЁЃШЛКѓзїепбаОПСЫЩёОдЊЕђЭіЕФаХКХЗжзгСзЫсЛЏЃЌЗЂЯжЧтЦјФмМѕЩйетаЉЗжзгЕФСзЫсЛЏЫЎЦНЁЃШчЯТЭМЮЊИУбаОПТлЮФЕФЭМаЮеЊвЊЁЃзїепШЯЮЊЧтЦјЮќШыЭЈЙ§МѕЩйСЫбѕЛЏгІМЄЃЌДгЖјМѕЩйСЫJunЕААзЕФСзЫсЛЏЫЎЦНЃЌМѕЩйСЫЯИАћЕђЭіЁЃБОбаОПКмживЊЕФаХЯЂЪЧЃЌЭЈЙ§ЯИАћЕђЭіБЃЛЄЃЌжЄУї1-8%ЕФЧтЦјЪЧаЇЙћзюКУЕФЃЌЕЋ16%ЕФаЇЙћКмвЛАуЁЃетвЛаХЯЂЧПСвЬсЪОЃЌЧтЦјЮќШыЕФаЇЙћВЂВЛЪЧдНЖрдНКУЁЃетКЭЮвУЧНёЬьСїааЕФзіЗЈЭъШЋВЛЭЌЃЌИјЮвУЧвЛИіЗЧГЃживЊЕФЬсабЁЃ
ЮќШыЧтЦј(H2)ЖдФдбЊЙмМВВЁЁЂЩёОЭЫааадМВВЁКЭаТЩњЖљДѓФдеЯАЃЌАќРЈгЩТщзэЦјЬхв§Ц№ЕФВЁРэзДЬЌОпгажЮСЦаЇЙћЁЃЮЊСЫРэНтЧтЦјЖдДѓФдБЃЛЄзїгУБГКѓЕФЛњжЦЃЌЮвУЧбаОПСЫдкЦпЗњУбгеЕМЕФЩёОдЊЫРЭіжаЪмЕНH2гАЯьЕФЗжзгаХКХЁЃЮвУЧШЗШЯЃЌЩёОЧАЬхЯИАћЖдЦпЗњУбУєИаЃЌВЂЧвдкгзЪѓЕФКѓПлДјЦЄВужаЗЂЩњЕђЭіЁЃНЋ1-8%ЕФH2ЦјЬхгыЦпЗњУбвЛЭЌИјвЉ3аЁЪБЃЌПЩвдвжжЦвђбѕЛЏгІМЄЖјЩ§ИпЕФcaspase-3НщЕМЕФЕђЭіадЯИАћЫРЭіЃЌВЂЭЌЪБМѕЩйc-JunСзЫсЛЏКЭc-JunЭЈТЗЕФМЄЛюЁЃТщзэв§Ц№ЕФжЌжЪЙ§бѕЛЏКЭбѕЛЏDNAЫ№ЩЫЭЈЙ§H2ЮќШыЕУЕНЛКНтЁЃСзЫсЕААзжЪзщЗжЮіНвЪОЃЌдкБЉТЖгкЦпЗњУбЕФаТЩњДѓФджаЃЌгаЗсИЛЕФВювьадСзЫсЛЏЕААзДиМЏЃЌАќРЈВЮгыЩёОдЊЗЂг§КЭЭЛДЅаХКХДЋЕМЕФЕААзЁЃH2ЮќШыаоИФСЫвРРЕИпСзЫсЛЏЕААзЕФЯИАћдЫЪфЭООЖЃЌАќРЈЮЂЙмЯрЙиЕААзМвзхЁЃетаЉаоИФПЩФмЩцМАH2ЖдПЙЦпЗњУбгеЕМЩёОдЊЫРЭіЕФБЃЛЄЛњжЦжаЁЃ
ИУТлЮФЭЈбЖзїепЮЊДѓдѓгєРЩНЬЪкЃЌЫћвВЪЧ2007ФъЧтЦјвНбЇЕьЛљадТлЮФЕФЕквЛзїепЁЃ
Iketani M, Hatomi M, Fujita Y, Watanabe N, Ito M, Kawaguchi H, Ohsawa I. Inhalation of hydrogen gas mitigates sevoflurane-induced neuronal apoptosis in the neonatal cortex and is associated with changes in protein phosphorylation. J Neurochem. 2024 Jun 7.
ШЋЮФдМ12000зжЁЃ
1 ЧАбд
ЗжзгЧтЃЈH2ЃЉзїЮЊвЛжжПЙбѕЛЏКЭПЙбзМСЃЌдкЖЏЮяМВВЁФЃаЭКЭСйДВбаОПжаЕФИјвЉЭООЖДѓжТПЩвдЗжЮЊСНжжЃКЮќШыЧтЦјКЭвћгУИЛЧтЫЎЁЃЧтЦјЮќШыЖдФдВПШБбЊ-дйЙрзЂЫ№ЩЫОпгаЯджјЕФЩёОБЃЛЄзїгУЁЃзюНќгаБЈЕРГЦЃЌФдШБбЊКѓКЭаФдржшЭЃКѓЕФЛЄРэЦкМфЮќШыЧтЦјФмдіМгЮоЩёОШБЯнДцЛюТЪЁЃвћгУИЛЧтЫЎПЩФмЛКНтАќРЈХСН№ЩВЁКЭАЂЖћДФКЃФЌВЁдкФкЕФЩёОЭЫааадМВВЁЁЃЫќЛЙПЩвдМѕЩйЪгЭјФЄМВВЁЁЂаФбЊЙмМВВЁЁЂИЮдрМВВЁЁЂДњаЛзлКЯеївдМАШеГЃЩњЛюжаЕФОЋЩёЦЃРЭЃЌДгЖјПЩФмЬсИпРЯФъШЫЕФЩњЛюжЪСПЁЃЮЊСЫНЋетаЉЗЂЯжНјвЛВНзЊЛЏЮЊСйДВЪЕМљЃЌгІВћУїЧтЦјЙІФмЕФЗжзгЛњжЦЁЃ
дкЖЏЮяФЃаЭжадЄЯШИјгшЧтЦјПЩвддЄЗРбзжЂадМВВЁЃЌетБэУїЧтЦјПЩФмМфНгзїгУгкбѕЛЏгІМЄЁЃзюНќЕФбаОПЯдЪОЃЌгУЧтЦјДІРэ1аЁЪБПЩвдвжжЦгЩЗ№ВЈДМ12-ШтЖЙоЂЫсѕЅ13-ввЫсѕЅгеЕМЕФжаадСЃЯИАћАћЭтЯнкхЕФаЮГЩЃЌЖјВЛЛсИФБфЛљвђБэДяЕФжївЊБфЛЏЁЃЮвУЧЗЂЯжЃЌНЋЩёОФИЯИАћБЉТЖгкЧтЦјжа1аЁЪБЃЌЛсднЪБадЧвПЩФцЕидіМгМИжжСзжЌЕФЫЎЦНЃЌвжжЦгыФмСПЩњВњЯрЙиЕФДњаЛЭООЖЃЌВЂЭЌЪБдіМгЮЂШѕЕФбѕЛЏгІМЄЁЃЪЕМЪЩЯЃЌМИКѕЫљгаШБбЊ-дйЙрзЂЫ№ЩЫФЃаЭжаЖМЙлВьЕНСЫЧтЦјжЮСЦ1-2аЁЪБЕФБЃЛЄаЇЙћЁЃетаЉРДздЯИАћКЭЖЏЮяЪЕбщЕФНсЙћБэУїЃЌЧтЦјЕФПЙбѕЛЏКЭПЙбзЙІФмЪЧЦфдчЦкЯИАћЕїНкЕФНсЙћЁЃ
аэЖрЦјЬхЗжзгЃЌАќРЈЧтЦјЃЌдкжЌжЪжаЕФШмНтЖШБШдкЫЎжаИпЁЃШмНтдкжЌжЪжаЕФЦјЬхПЩФмЛсгАЯьЯИАћКЭЩњРэЙІФмЁЃОЁЙмТщзэМСЕФШЗЧаЗжзгЛњжЦдкКмДѓГЬЖШЩЯШдШЛЮДжЊЃЌЕЋMeyer-OvertonЙиСЊЬсЙЉСЫТщзэЦјЬхаЇСІгыЦфдкРржЌЗЧМЋадНщжЪжаШмНтЖШжЎМфЕФСЊЯЕЁЃШмНтдкжЌжЪжаЕФТщзэЦјЬхЭЈЙ§ЪшЫЎЯрЛЅзїгУИФБфЯИАћФЄНсЙЙЁЃОЁЙмТщзэЦјЬхЪЧЗЧГЃАВШЋЕФвЉЮяЃЌЕЋЫќУЧЖдЮДГЩЪьДѓФдЕФЪЙгУЛсв§Ц№ВЁРэБфЛЏКЭГЄЦкЕФааЮЊвьГЃЁЃДЫЭтЃЌаТЩњаЁЪѓдкЮќШыЦпЗњУбЦкМфЮќШыЧтЦј6аЁЪБЃЌПЩвдвжжЦЦпЗњУбв§Ц№ЕФбѕЛЏгІМЄдіМгЁЂЩёОдЊЕђЭівдМАЫцКѓЕФЩёОааЮЊШБЯнЁЃЖјЧвЃЌЮќШыЧтЦјЛЙФмзшжЙГЩФъКѓгЩгкаТЩњЦкБЉТЖгкЦпЗњУбЖјв§Ц№ЕФвьГЃФИадааЮЊЃЌетБэУїЧтЦјОпгаМѕЩйТщзэвЉЮяв§Ц№ИБзїгУЕФОоДѓЧБСІЁЃдкетЯюбаОПжаЃЌЮвУЧЕїВщСЫЧтЦјдкЦпЗњУбгеЕМЕФЩёОдЊЯИАћЫРЭіжагАЯьЕФЗжзгаХКХЃЌвдИќКУЕиРэНтЧтЦјЖдДѓФдБЃЛЄзїгУЕФЛњжЦЁЃ
2 ВФСЯгыЗНЗЈ
2.1 ЖЏЮя
ЫљгаЖЏЮяЪЙгУКЭЪЕбщЗНАИОљзёбЁЖЪЕбщЪвЖЏЮяЛЄРэддђЁЗЃЈNIHГіАцЮяБрКХ86-23ЃЌ1985ФъаоЖЉЃЉЁЃЫљгабаОПавщОљгЩЖЋОЉЖМРЯФъвНбЇбаОПЫљЕФЖЏЮяЛЄРэЮЏдБЛсЩѓВщВЂХњзМЃЈХњзМДњТыЃЌ23027ЃЉЁЃдкЪЕбщЙ§ГЬжаЃЌЮвУЧОЁвЛЧаХЌСІМѕЩйЖЏЮяЪ§СПМАЦфЭДПрЁЃДгSLCЃЈЖЋОЉЃЌШеБОЃЉЙКТђЕФC57BL/6NЖЈЪБЛГдааЁЪѓБЛАВжУдк22ЁуCЕФЛЗОГжаЃЌЙтеежмЦкЮЊ12аЁЪБЙтее/12аЁЪБКкАЕЃЌздгЩЛёШЁЮоОњЪГЮяКЭЫЎЁЃГіЩњШеБЛБъМЧЮЊP0ЁЃЮДШЗЖЈгзсЬЕФадБ№ЁЃбаОПжаЮДНјааЫцЛњЗжХфЃЌвВЮДдЄЯШШЗЖЈХХГ§БъзМЁЃзмЙВЪЙгУСЫ18жЛФИЪѓКЭ142жЛгзсЬНјааЪЕбщЁЃИУбаОПЮДНјаадЄзЂВсЁЃ
2.2 БЉТЖгкКЌЧтТщзэЦјЬхжа
дкГіЩњКѓЕк6ЬьЃЌНЋаЁЪѓгзсЬДгФИЪѓС§жаШЁГіЃЌСЂМДЗХШыдЄШШжС37ЁуCЕФЪЊШѓБћЯЉЫсЪвЃЈФкВПЬхЛ§6.8Щ§ЃЉжаЁЃУПИіЪвФкЕФгзсЬЪ§СПЮЊ3-6жЛЁЃЙиБеВЂУмЗтИЧзгКѓЃЌвд1.0Щ§/ЗжжгЕФСїСПЯђЪвФкГфШыТщзэЦјЬхЛьКЯЮя20ЗжжгЃЌШЛКѓдке§ГЃбЙСІЯТНЋЦфжУгк37ЁуCЕФЗѕЛЏЦїжа3аЁЪБЃЈЭМS1ЃЉЁЃЫцКѓвЛаЉЪвБЛДђПЊЃЌСЂМДгУПеЦјЭЈЗчЃЌВЂдк37ЁуCЯТдйБЃГж3аЁЪБЁЃИљОнжЎЧАЕФбаОПЃЌТщзэЦјЬхЛьКЯЮяАќКЌ3%ЦпЗњУбЁЂ30%бѕЦјКЭ0-16%ЧтЦјЃЌЦфгрЮЊЕЊЦјЦНКтЁЃРДздВЛЭЌФИЪѓЕФгзсЬБЛЛьКЯВЂЫцвтЗжЮЊЮхзщЃЌЗжБ№БЉТЖгквдЯТЛЗОГЃКЃЈ1ЃЉПеЦј3аЁЪБЃЈЖдеезщЃЉЃЌЃЈ2ЃЉВЛКЌЧтЦјЕФЛьКЯТщзэЦјЬх3аЁЪБЃЈЦпЗњУбзщЃЉЃЌЃЈ3ЃЉКЌ2%ЧтЦјЕФЛьКЯТщзэЦјЬх3аЁЪБЃЈЦпЗњУб+ЧтЦјзщЃЉЃЌЃЈ4ЃЉВЛКЌЧтЦјЕФЛьКЯТщзэЦјЬх3аЁЪБЃЌШЛКѓПеЦј3аЁЪБЃЈЦпЗњУб/ПеЦјзщЃЉЃЌвдМАЃЈ5ЃЉКЌ2%ЧтЦјЕФЛьКЯТщзэЦјЬх3аЁЪБЃЌШЛКѓПеЦј3аЁЪБЃЈЦпЗњУб/ПеЦј+ЧтЦјзщЃЛМћЭМS1ЃЉЁЃЮЊСЫШЗЖЈеЙЪОзюЧППЙЕђЭіЛюадЕФЧтЦјХЈЖШЃЌНЋгзсЬБЉТЖгкКЌга0-16%ЧтЦјЕФЛьКЯТщзэЦјЬхжа3аЁЪБЃЌШЛКѓдйБЉТЖгкПеЦјжа3аЁЪБЁЃжЎКѓЮвУЧдкРфгеЕМТщзэЯТНЋгзсЬЖЯЭЗЃЌСЂМДШЁГіДѓФдЃЌВЂНЋзѓАыЧђЕФДѓФдЦЄВудкИЩБљЩЯЫйЖГЁЃбљБОДЂДцгк-80ЁуCЁЃгвАыЧђгУ4%ЖрОлМзШЉдкСзЫсбЮЛКГхбЮЫЎЃЈPBSЃЉжаЙЬЖЈЙ§вЙЃЌдк4ЁуCЯТгУ30%есЬЧ/PBSБЃЛЄЃЌгУзюМбЧаИюЮТЖШЃЈOCTЃЉАќТёНщжЪЃЈФПТМКХ45833ЃЌSAKURAЃЌЖЋОЉЃЌШеБОЃЉРфЖГЃЌВЂДЂДцгк-80ЁуCЁЃ
ЮвУЧЪЙгУGIGAМЦЫуЦїЭјеОЕФPower & Sample Size CalculatorШЗЖЈСЫУПзщЕФбљБОСПЮЊn=19ЁЃЪЕбщЩшМЦВЮЪ§ЃКIаЭДэЮѓТЪЃЈІСЃЉ= 5%ЃЌЙІаЇЃЈ1−ІТЃЉ= 85%ЃЌжЮСЦзщЪ§ = 7ЁЃЪ§ОнВЮЪ§ЃКH0ЯТЕФОљжЕ = 100ЃЌБъзМВю = 40ЃЌзюаЁПЩМьВтаЇгІЃЈMDEЃЉ= 30ЃЌгХдНадБпНч = 0ЁЃетаЉВЮЪ§ЛљгкЯШЧАбаОПЃЈYonamineЕШШЫЃЌ2013ФъЃЉЕФЪ§ОнбЁдёЁЃ
2.3 УтвпШОЩЋКЭФЉЖЫЭббѕКЫмеЫсзЊвЦУИНщЕМЕФdUTPШБПкФЉЖЫБъМЧЃЈTUNELЃЉВтЖЈ
OCTАќТёЕФДѓФддкРфЖГЧаЦЌЛњЩЯЧаГЩ20ІЬmКёЕФЙкзДЧаЦЌЁЃУПИіЧаЦЌгУЩНбђбЊЧхЗтБеЃЌШЛКѓдк4ЁуCЯТгывдЯТеыЖдаЁЪѓПЙдЕФвЛПЙЙ§вЙЗѕг§ЃКЕЅПЫТЁЭУПЙСбНтаЭcaspase-3ЃЈCC3ЃЉЃЌгУгкМьВтгЩAsp175ИННќЧаИюВњЩњЕФДѓЦЌЖЮcaspase-3ЃЛЕЅПЫТЁДѓЪѓПЙГВЕААзЁЂПЙGFAPЁЂПЙNeuNЁЂПЙCD31ЁЂПЙOlig-2ЁЂПЙF4/80ЁЂПЙНсЕААзЃЛЕЅПЫТЁаЁЪѓПЙc-JunКЭЕЅПЫТЁаЁЪѓПЙСзЫсЛЏc-JunЃЌгУгкМьВтSer63СзЫсЛЏЕФc-Jun p39ЁЃШЛКѓУПИіЧаЦЌдкЪвЮТЃЈRTЃЉЯТгУвдЯТЪЪЕБЕФЖўПЙIgGПЙЬхЗѕг§60ЗжжгЃКЩНбђПЙаЁЪѓНсКЯAlexa Fluor 594ЁЂПЙЭУНсКЯAlexa Fluor 594ЁЂПЙЭУНсКЯAlexa Fluor 647ЃЌвдМАПЙДѓЪѓНсКЯAlexa Fluor 488ЁЂПЙЩНбђНсКЯAlexa Fluor 488КЭПЙЭУНсКЯCF680RЁЃбљБОдкМЄЙтЙВОлНЙЯдЮЂОЕЩЯГЩЯёЁЃЮЊСЫЖЈСПУтвпзщжЏЛЏбЇШОЩЋCC3ЃЌЮвУЧЪЙгУСЫЩњЮяЫиЛЏЕФПЙЭУIgGКЭЧзКЭЫи-РБИљЙ§бѕЛЏЮяУИзККЯЮяЁЃУтвпШОЩЋгУ3,3'-ЖўАБЛљСЊБНАЗЯдЩЋЃЌЧаЦЌгУЫеФООЋИДШОЁЃдкаЈЧАЦЄжЪжаМЦЪ§бєадЯИАћЁЃ
ЖдгкTUNELВтЖЈЃЌЮвУЧЪЙгУApopTag®дЮЛЕђЭіМьВтЪдМСКаАДеежЦдьЩЬЕФавщНјааЁЃМђЖјбджЎЃЌЧаЦЌдкввДМЃКДзЫс2:1жаКѓЙЬЖЈЁЃдкгУPBSжаЕФ3%Й§бѕЛЏЧтДуУ№ФкдДадЙ§бѕЛЏЮяУИКѓЃЌЧаЦЌдк37ЁуCЯТгыФЉЖЫЭббѕКЫмеЫсзЊвЦУИЗѕг§ЃЌЫцКѓгыПЙЕиИпаСЙ§бѕЛЏЮяУИзККЯЮяЗѕг§ЁЃгУDABЯдЩЋЕФTUNELбєадЯИАћБЛПЩЪгЛЏЁЃУПИіЧаЦЌЕФаЈЧАЦЄжЪФкЕФбєадЯИАћдк1-2ИіЪгвАЃЈУПИі0.3ЕН0.5 mm²ЃЉФкЪжЖЏМЦЪ§ЁЃ
2.4 бѕЛЏгІМЄБъжОЮяЕФУтвпзщжЏЛЏбЇ
4%ЖрОлМзШЉЙЬЖЈЕФOCTАќТёДѓФдЕФЙкзДЧаЦЌЃЈ20ІЬmКёЃЉШчЧАЫљЪіЖдбѕЛЏгІМЄБъжОЮя8-єЧЛљ-2'-ЭббѕФёмеЃЈ8-OHdGЃЉКЭ4-єЧЛљ-2-ШЩЯЉШЉЃЈ4-HNEЃЉНјааШОЩЋЁЃМђЖјбджЎЃЌЧаЦЌдкФћУЪЫсЛКГхвКжаМгШШ5ЗжжгвдДйНјПЙдЛжИДЃЌгУЩНбђбЊЧхЗтБеЃЌВЂдк4ЁуCЯТгыеыЖд8-OHdGКЭ4-HNEЕФвЛПЙЕЅПЫТЁПЙЬхЙ§вЙЗѕг§ЁЃЯДЕгКѓЃЌЧаЦЌдкЪвЮТЯТгУЩњЮяЫиЛЏЕФПЙаЁЪѓIgGЖўПЙЗѕг§60ЗжжгЃЌВЂНјвЛВНдкЪвЮТЯТгУМюадСзЫсУИЗѕг§30ЗжжгЁЃЪЙгУ5-фх-4-ТШ-3-пХпсСзЫс/ЯѕЛљРЖЫФпђзїЮЊЩњЩЋЕзЮяЪЕЯжбеЩЋЗДгІЁЃУПИіЧаЦЌЕФаЈЧАЦЄжЪФкЕФбєадЯИАћдк1-2ИіЪгвАЃЈУПИі0.3ЈC0.5 mm²ЃЉФкЪжЖЏМЦЪ§ЁЃ
2.5 ЕААзжЪУтвпгЁМЃЗЈ
НЋРфЖГЕФДѓФдЦЄВудкЗХЩфадУтвпГСЕэЗжЮіЃЈRIPAЃЉЛКГхвКжадШНЌЃЌИУЛКГхвККЌгаЕААзУИвжжЦМСЛьКЯЮяЃЌВЂдк4ЁуCЯТвд15000gРыаФ20ЗжжгЁЃЪеМЏЩЯЧхвКВЂДЂДцгк-80ЁуCЁЃНтЖГКѓЃЌЖдгкc-JunЃЌШШБфадЕААзЃЈУПЕР5ІЬgЃЉдк10%БћЯЉѕЃАЗФ§НКЩЯЗжРыВЂЕчзЊвЦЕНОлЦЋЗњввЯЉЃЈPVDFЃЉФЄЩЯЁЃгУЭбжЌХЃФЬЗтБеФЄЃЌШЛКѓдк4ЁуCЯТгыПЙc-JunЃЈ1:3000ЃЉКЭСзЫсЛЏc-JunЃЈ1:3000ЃЉЕФвЛПЙЕЅПЫТЁаЁЪѓПЙЬхЙ§вЙЗѕг§ЁЃЖдгкCC3ЃЌШШБфадЕААзЃЈУПЕР10ІЬgЃЉдк15%БћЯЉѕЃАЗФ§НКЩЯЗжРыВЂЕчзЊвЦЕНPVDFФЄЃЈ0.2ІЬmЮЂПзЃЉЩЯЁЃгУЭбжЌХЃФЬЗтБеФЄЃЌШЛКѓдк4ЁуCЯТгывЛПЙЭУПЙCC3ЃЈ1:3000ЃЉЙ§вЙЗѕг§ЁЃЯДЕгКѓЃЌФЄдкЪвЮТЯТгУРБИљЙ§бѕЛЏЮяУИХМСЊЕФПЙаЁЪѓIgGЗѕг§1аЁЪБЁЃЪЙгУдіЧПЛЏбЇЗЂЙтЪдМСКаМьВтЕААзДјЃЌВЂгУCCDГЩЯёЯЕЭГПЩЪгЛЏЁЃзїЮЊФкВЮЃЌФЄБЛАўРыВЂгыЖрПЫТЁЭУПЙИЪгЭШЉ-3-СзЫсЭбЧтУИПЙЬхКЭРБИљЙ§бѕЛЏЮяУИХМСЊЕФПЙЭУIgGЗѕг§ЁЃ
2.6 ЖЈСПPCR
ЪЙгУRNeasy Plus MiniЪдМСКаДгРфЖГЕФДѓФдЦЄВужаЬсШЁзмRNAЃЌВЂАДеежЦдьЩЬЕФавщЪЙгУЕквЛСДКЯГЩЯЕЭГНјааФцзЊТМЁЃЭЈЙ§ЪЙгУЛљгкSYBR Green IЕФqPCRЛьКЯвККЭStepOne PlusЪЕЪБPCRЯЕЭГЃЌвдШ§жиИДВтЖЈзЊТМЫЎЦНЁЃгУгкРЉдіЕФв§ЮяСагкБэ1ЁЃЛљвђБэДяСПЙщвЛЛЏжСУПИібљБОжаЕФRpl4БэДяСПЁЃ
TABLE 1. List of primers used for quantitative real-time PCR.
Gene | Fwd/rev | Primer sequence (5ЁфЈC3Ёф) |
Fas | Forward | GAGAATTGCTGAAGACATGACAATCC |
Reverse | GTAGTTTTCACTCCAGACATTGTCC | |
Fasl | Forward | CAGCCCATGAATTACCCATGT |
Reverse | ATTTGTGTTGTGGTCCTTCTTCT | |
Bcl2 | Forward | TAAGCTGTCACAGAGGGGCT |
Reverse | TGAAGAGTTCCTCCACCACC | |
Jun | Forward | ACGACCTTCTACGACGATGC |
Reverse | CCAGGTTCAAGGTCATGCTC | |
Fos | Forward | GAATGGTGAAGACCGTGTCAG |
Reverse | CTGCAACGCAGACTTCTCATC | |
Egr1 | Forward | CTGCTTCATCGTCTTCCTCT |
Reverse | CCTGTTGTTGTGGAAACAGA | |
Ddit3 | Forward | CTGCCTTTCACCTTGGAGAC |
Reverse | CGTTTCCTGGGGATGAGATA | |
Ppp1r15a | Forward | GGTGGTCCAGCTGAGAATGA |
Reverse | TCTTCCGTGGCTTGATGTTC | |
Hmox1 | Forward | TGCTAGCCTGGTGCAAGATA |
Reverse | GCCAACAGGAAGCTGAGAGT | |
Rpl4 (internal control) | Forward | GCCAAGACTATGCGCAGGAAT |
Reverse | GTAGCTGCTGCTTCCAGCTT |
2.7 бЊСїКЭбѕЦјЫЎЦНВтСП
ЪЙгУМЄЙтЩЂАпЖдБШГЩЯёвЧВтСПбЊСїЃЌИУЩшБИгЩвЛИіКьЭтАыЕМЬхМЄЙтЦїЃЈВЈГЄ785 nmЃЉКЭвЛИіЕчКЩёюКЯЦїМўЯрЛњЃЈmoor FLIP2ЃЉзщГЩЁЃЩшБИЩшжУдкаЁЪѓИЙВПЩЯЗНЃЌЕїећЫѕЗХвдАќРЈећИіЩэЬхЁЃЪгвАдМЮЊ3136 mm²ЃЈ64.0 ЁС 49.0 mmЃЉЁЃвЛИіЦНУцЭМЯёгЩ752 ЁС 580ЯёЫизщГЩЃЌЯёЫиДѓаЁдМЮЊ85 ІЬmЁЃЮЊСЫСПЛЏЧјгђбЊСїЃЌДгИЙЙЩЙЕЧјгђЕФИааЫШЄЧјгђдМ16.4 mm²ЃЈ4.0 ЁС 4.1 mmЃЉЬсШЁЪ§ОнЁЃЪЙгУЗЧЧжШыЪНТіВЋбЊбѕвЧВтСПбЊбѕЫЎЦНЁЃ
2.8 СзЫсЕААзжЪзщбЇЗжЮі
ЭЈЙ§ФЩУзвКЯрЩЋЦз-ДЎСЊжЪЦзЃЈLC-MS/MSЃЉНјааЕФСзЫсЕААзжЪзщбЇЗжЮіКЭЪ§ОнДІРэдкProteobiologicsНјааЁЃ
2.8.1 LC-MS/MSЗжЮіЕФбљЦЗжЦБИ
НЋРфЖГЕФаЈЧАЦЄжЪдкКЌгаЕААзУИвжжЦМСЛьКЯЮяКЭСзЫсУИвжжЦМСЛьКЯЮяЕФЯрзЊвЦБэУцЛюадМСЛКГхвКжадШНЌЃЌ95ЁуCЗѕг§7ЗжжгЃЌГЌЩљДІРэВЂРыаФЁЃШЛКѓНЋЩЯЧхвКзЊвЦЕНаТЕФЙмзгжаЁЃУПИібљЦЗгУЯрзЊвЦБэУцЛюадМСЛКГхвКШмНтЁЃУПИібљЦЗЙВ200 ІЬgзЊвЦЕН1.5 mLЙмжаЃЌ95ЁуCжѓЗа5ЗжжгЁЃЪЙгУBioruptorГЌЩљЦїЖдСбНтвКНјааГЌЩљДІРэЁЃШЛКѓЃЌгУ10 mM2-єШввЛљШ§ьЂЛЙдбљЦЗЃЌгУ20 mMЕтввѕЃАЗЭщЛљЛЏЃЌВЂЪЙгУSP3авщДПЛЏЕААзжЪЁЃДПЛЏЕФЕААзжЪгУвШЕААзУИЃЈЕААзжЪжиСПЃК1/50ЃЉКЭжЪСПЙтЦзМЖЕФРЕАБѕЃФкыФУИдк37ЁуCЯТЯћЛЏ16аЁЪБЁЃ
2.8.2 ШЅбЮЁЂИЛМЏСзЫсыФЁЂДЎСЊжЪСПБъЧЉЃЈTMTЃЉБъМЧ
ИљОнЯШЧАУшЪіЕФавщЃЌЪЙгУOASISЧзЫЎ-ЪшЫЎЦНКтЖдыФЖЮНјааШЅбЮЃЌВЂгУFe3+ЙЬЖЈЛЏН№ЪєЧзКЭВуЮіЃЈFe-IMACЃЉЪїжЌИЛМЏСзЫсыФЁЃдкБОбаОПжаЃЌЮвУЧдкНзЖЮМтЖЫгУFe-IMACЪїжЌИЛМЏСЫСзЫсыФЁЃМђЖјбджЎЃЌНЋC18ХЬЩшжУдк200 ІЬLвЛДЮадМтЖЫжаЃЌВЂдкC18ХЬЩЯЩшжУFe-IMACЪїжЌЁЃШЅбЮЕФыФЖЮЭЈЙ§Fe-IMAC/C18НзЖЮМтЖЫЁЃгУ60%ввыцМг0.1%Ш§ЗњввЫсЯДЭбСзЫсыФЃЌВЂдкЯТВПC18ХЬЩЯДПЛЏЁЃЮЊСЫЖЈСПУПИібљЦЗжаЕФСзЫсЮЛЕуЃЌИљОнжЦдьЩЬЕФавщЖдУПИіИЛМЏСзЫсыФЕФбљЦЗНјааTMTБъМЧЁЃбљЦЗЫцКѓБЛецПеРыаФжСНќИЩЁЃУПИізщЗждк2%ввыцЃЌ0.1%Ш§ЗњввЫсжажиЙЙЃЌгУгкLC-MS/MSДІРэЁЃ
2.8.3 LC-MS/MSЗжЮі
LC-MS/MSЗжЮіЪЙгУгыUltimate 3000КЭHTC-PALёюКЯЕФQ Exactive PlusНјааЁЃФЩУзвКЯрЩЋЦзЬнЖШгЩЛКГхвКAЃЈ0.1%МзЫсЃЉгы5-30%ЛКГхвКBЃЈ0.1%МзЫсКЭ99.9%ввыцЃЉдк85ЗжжгФкаЮГЩЃЛСїЫйЮЊ280 nL/minЁЃQ ExactiveвЧЦївдЪ§ОнвРРЕФЃЪНВйзїЁЃдкРызгРлЛ§жС3ЁС10^6ФПБъжЕКѓЃЌдкOrbitrapжавд70,000ЗжБцТЪЛёШЁЕїВщШЋЩЈУшжЪЦзЃЈm/z 350ЈC1800ЃЉЁЃЖЏЬЌХХГ§ЩшжУЮЊ30УыЁЃзюЧПСвЕФ12ИіЖрЕчКЩРызгЃЈzЁн2ЃЉБЛЫГађРлЛ§жС1ЁС10^5ФПБъжЕЃЌВЂдкХізВГижаЭЈЙ§ИпФмСПХізВНтРыЃЈHCDЃЉНјааЫщЦЌЛЏЃЌзюДѓзЂЩфЪБМфЮЊ120КСУыЃЌЗжБцТЪЮЊ35,000ЁЃЕфаЭЕФжЪЦзЬѕМўШчЯТЃКХчЮэЕчбЙЃЌ2 kVЃЛМгШШУЋЯИЙмЮТЖШЃЌ250ЁуCЃЛБъзМЛЏHCDХізВФмСПЃЌ30%ЁЃLC-MS/MSРызгбЁдёуажЕЩшжУЮЊ2.5ЁС10^4МЦЪ§ЁЃбЁдёСЫ0.7 DaЕФИєРыПэЖШЁЃ
2.8.4 СзЫсыФЕФМјЖЈКЭЖЈСП
ЪЙгУMaxQuant1.6.14.0НјааСзЫсыФЕФМјЖЈЁЃНсКЯСЫ253жжГЃМћЮлШОЮяЕФШЫРрЕААзжЪUniProtЪ§ОнПтЃЈ2017_01АцБОЃЉгУгкЗжЮіLC-MS/MSЙтЦзЁЃУИЬивьадЩшжУЮЊвШЕААзУИ/PЁЃдЪаэзюЖрСНИіЮЛЕуЕФВЛе§ШЗЧаИюЁЃМйЩшАыызАБЫсВаЛљЕФЙЬЖЈаоЪЮЮЊєШМзЛљЛЏЃЌЖјЕААБЫсбѕЛЏКЭЫПАБЫсЁЂЫеАБЫсЁЂРвАБЫсЕФСзЫсЛЏБЛЩшЖЈЮЊПЩБфаоЪЮЁЃЕААзжЪзщЁЂыФЖЮКЭСзЫсЮЛЕуЕФМйЗЂЯжТЪЃЈFDRЃЉаЁгк0.01ЁЃTMTБЈИцРызгЧПЖШЕФЖЈСПжЕгЩMaxQuantзмНсЁЃБЛЪЖБ№ЮЊЗДЯђЛђЧБдкЮлШОЮяЕФыФЖЮВЛгУгкКѓајЗжЮіЁЃУПИіСзЫсЮЛЕуЕФЖЈЮЛИХТЪЕФНиЖЯБъзМГЌЙ§0.75ЁЃ
2.8.5 Ъ§ОнДІРэ
ЪЙгУPerseus 1.6.14.0НјааЪ§ОнДІРэЁЃЖЈСПTMTБЈИцРызгЧПЖШОЙ§log2зЊЛЛВЂЭЈЙ§УПИіTMTЭЈЕРжаЕФжЕЕФжаЮЛЪ§жааФЛЏНјааЙщвЛЛЏЁЃ
2.9 ЪЙгУPhos-tagММЪѕИЛМЏКЭЕААзжЪУтвпгЁМЃЗжЮіСзЫсЕААз
БљЖГЕФаЈЧАЦЄжЪдкВЛКЌSDSЕФRIPAЛКГхвКжадШНЌЃЌИУЛКГхвККЌгаЕААзУИвжжЦМСЛьКЯЮяКЭСзЫсУИвжжЦМСЛьКЯЮяЁЃдШНЌбљБОдк4ЁуCЯТвд15000ЁСgРыаФ20ЗжжгЁЃЪеМЏЩЯЧхвКВЂДЂДцгк-80ЁуCЁЃЪЙгУPhos-tagЧэжЌЬЧДгНтЖГбљБОжаЬсШЁСзЫсЛЏЕААзжЪЁЃPhos-tagавщЪзЯШЭЈЙ§гУ21GеыЭЗДЬДЉ0.5 mLЙмЕзВПДДНЈвЛИіаЁПзЃЌВЂНЋДЫЙмжУгк1.5 mLЙмФкПЊЪМЁЃШЛКѓЃЌЯђ0.5 mLЙмжаМгШы40 ІЬLЕФ50% (v/v) Phos-tagЧэжЌЬЧаќИЁвКЃЌНЋЕЅдЊРыаФЃЈ2000 gЃЉ20УыКѓЖЊЦњТЫвКЁЃНгзХЃЌМгШы40 ІЬLЦНКтЛКГхвКЃЌдйДЮРыаФВЂдк5ЗжжгКѓЖЊЦњТЫвКЁЃФ§НКгУ40 ІЬLНсКЯ/ЯДЕгЛКГхвКЃЈ0.1 M Tris-CH3COOH (pH 7.5) КЭ 1 M CH3COONaЃЉЯДЕгШ§ДЮЃЌАщЫцРыаФЁЃЫцКѓЃЌЯђУПИіКЌга20 ІЬgЕААзжЪЕФдШНЌбљБОжаМгШы10 ІЬLЃЌЦНКт5ЗжжгЃЌШЛКѓРыаФвдЪеМЏСїЙ§ЮяЁЃШЛКѓНЋгыСзЫсЕААзНсКЯЕФФ§НКгУ0.1 mLНсКЯ/ЯДЕгЛКГхвКЯДЕгВЂРыаФЁЃЯђФ§НКжаМгШыжСЩй40 ІЬLеєСѓЫЎКѓЃЌНЋЛьКЯЮязЊвЦЕНаТЙмжаВЂРыаФвдЖЊЦњЩЯЧхвКЁЃзюКѓЃЌЯђФ§НКжаМгШы10 ІЬLбљЦЗЩЯбљШОСЯЃЌШЛКѓдк95ЁуCМгШШ5ЗжжгЁЃШШБфадбљБОдк3%БћЯЉѕЃАЗ/0.5%ЧэжЌЬЧФ§НКЩЯЗжРыЃЌВЂдк4ЁуCЯТЪЊзЊЕНPVDFФЄЩЯ16аЁЪБЁЃФЄгУЭбжЌХЃФЬЗтБеЃЌВЂдк4ЁуCЯТгыMAP1AКЭMAP1BЕФвЛПЙЙ§вЙЗѕг§ЁЃЯДЕгКѓЃЌФЄдкЪвЮТЯТгыРБИљЙ§бѕЛЏЮяУИХМСЊЕФПЙаЁЪѓIgGЃЈ1:10000ЃЉКЭПЙЭУIgGЃЈ1:10000ЃЉЗѕг§1аЁЪБЁЃЕААзжЪДјгУдіЧПЛЏбЇЗЂЙтЪдМСКаМьВтЃЌВЂгУCCDГЩЯёЯЕЭГПЩЪгЛЏЁЃ
2.10 ЭГМЦЗжЮі
ЪЙгУPrism 8НјааЭГМЦЗжЮіЁЃЫљгажЕБэЪОЮЊЦНОљжЕЁРБъзМЦЋВюЃЈSDЃЉЁЃЭЈЙ§Shapiro-WilkКЭKolmogorov-SmirnovВтЪдШЗШЯЪ§ОнЕФе§ЬЌадЁЃЭЈЙ§ДјгаTukeyВтЪдЕФЕЅвђЫиЗНВюЗжЮіЃЈANOVAЃЉШЗЖЈЯджјадЁЃp<0.05ЪгЮЊЯджјЁЃЪ§ОнЮДНјааРыШКжЕВтЪдЁЃЮвУЧЪеМЏСЫАќРЈFжЕКЭpжЕдкФкЕФЫљгаЭГМЦЪ§ОнЃЌгУгкЮвУЧЕФANOVAКЭtМьбщЗжЮіЃЈЪ§ОнS1ЃЉЁЃЖдгкLC-MS/MSЗжЮіЃЌЪЙгУWelchЕФtМьбщШЗЖЈИааЫШЄЬѕМўжЎМфЕФЭГМЦЯджјадЁЃpжЕЭЈЙ§Benjamini-HochbergЗНЗЈНјааFDRаЃе§ЁЃ
3 НсЙћ
3.1 ЦпЗњУбгеЕМЕФаТЩњЪѓДѓФдЩёОзцЯИАћЕђЭі
ЮвУЧЪзЯШдкБЉТЖгкЦпЗњУбЕФаТЩњаЁЪѓДѓФджаЪЖБ№ГіЕђЭіЯИАћЁЃЮвУЧдк6ЬьДѓЕФаЁЪѓБЉТЖгк3%ЦпЗњУб3аЁЪБКѓЃЌжївЊдкЗЂг§ЛКТ§ЕФаЈЧАЦЄжЪЃЈЫцзХЖЏЮябЇЛсдкЛЗОГжаЕМКНЖјЗЂг§ЃЉЙлВьЕНПЙCC3ПЙЬхШОЩЋЛђTUNELбєадЕФЕђЭіЯИАћЃЈЭМ1ЃЉЁЃЕђЭігЩФкдкЛђЭтдкЭООЖНщЕМЃЌЕМжТcaspase-3ЕФЧаИюЁЃCC3бєадЯИАћгыПЙГВЕААзПЙЬхЙВШОЃЈЭМ1cЃЉЃЌЕЋВЛгыNeuNЃЈГЩЪьЩёОдЊЕФБъжОЮяЃЛЭМ1aЃЉЁЂНКжЪЯЫЮЌЫсадЕААзЃЈGFAPЃЛаЧаЮНКжЪЯИАћЕФБъжОЮяЃЛЭМ1bЃЉЁЂCD31ЃЈФкЦЄЯИАћЕФБъжОЮяЃЛЭМ1dЃЉЁЂПЙOlig2ЃЈЩйЭЛНКжЪЯИАћЕФБъжОЮяЃЛЭМ1eЃЉЁЂПЙF4/80ЃЈЮЂНКжЪЯИАћКЭОоЪЩЯИАћЕФБъжОЮяЃЛЭМ1fЃЉЛђПЙНсЕААзЃЈжмЯИАћЕФБъжОЮяЃЛЭМ1gЃЉЙВШОЁЃгЩгкДцдкГВЕААзбєадЕФжмЯИАћЃЌCC3бєадЯИАћЭЌЪБгУПЙГВЕААзКЭПЙНсЕААзПЙЬхЙВШОЁЃЮвУЧЗЂЯжЫЋбєадЯИАћВЛЪЧCC3бєадЁЃетаЉНсЙћБэУїЃЌЦпЗњУбБЉТЖПЩгеЕМЕђЭіЕФЯИАћЪЧЩёОзцЯИАћЁЃ
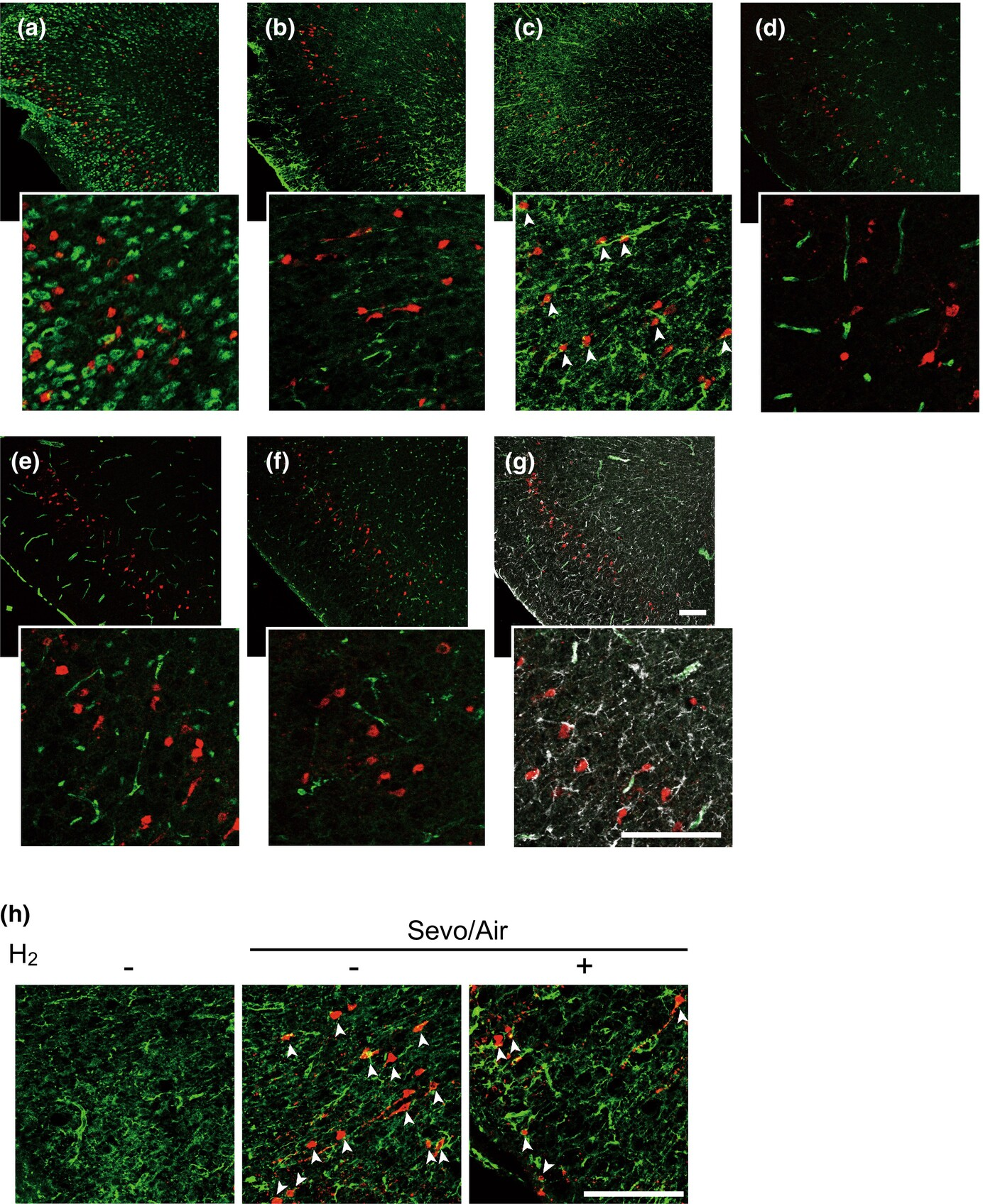
ЭМ1ЦпЗњУбгеЕМаЈЧАЦЄжЪЩёОзцЯИАћЕђЭіЁЃ(aЈCg) ДњБэадЙВОлНЙЯдЮЂОЕЭМЯёЯдЪОЃЌЦпЗњУбгеЕМЕФCC3бєадЯИАћЃЈКьЩЋЃЉЃЌгыПЙNeuN (a)ЁЂПЙGFAP (b)ЁЂПЙГВЕААз (c)ЁЂПЙCD31 (d)ЁЂПЙOlig2 (e)ЁЂПЙF4/80ЃЈТЬЩЋЃЛfЃЉЛђПЙНсЕААзЃЈТЬЩЋЃЉКЭПЙГВЕААзЃЈАзЩЋЃЛgЃЉЙВШОЁЃгзЪѓБЉТЖгкЛьКЯТщзэЦјЬх3аЁЪБЃЌШЛКѓБЉТЖгкПеЦј3аЁЪБЁЃЃЈhЃЉДгБЉТЖгкКЌЛђВЛКЌ2%ЧтЦјЕФЛьКЯТщзэЦјЬх3аЁЪБМАЫцКѓБЉТЖгкПеЦј3аЁЪБЃЈSevo/AirЃЉЕФгзЪѓжаЬсШЁЕФаЈЧАЦЄжЪЃЌгУПЙCC3ЃЈКьЩЋЃЉКЭПЙГВЕААзЃЈТЬЩЋЃЉПЙЬхЙВШОЁЃМ§ЭЗжИЪОГВЕААзКЭCC3ЫЋбєадЯИАћЁЃБШР§ГпЃК100ІЬmЁЃ
3.2 ЧтЦјЖдЦпЗњУбгеЕМЕђЭіЕФЩёОБЃЛЄзїгУ
ЮЊСЫЦРЙРЧтЦјЖдЦпЗњУбгеЕМЕђЭіЕФБЃЛЄзїгУЃЌаТЩњаЁЪѓБЛЭЌЪБИјгш3%ЦпЗњУбЃЌгаЛђЮо2%ЧтЦјЙВДІРэ3аЁЪБЃЈЗжБ№ЮЊSevoзщКЭSevo+H2зщЃЉЁЃЯШЧАЕФбаОПБэУїЃЌдкФдШБбЊдйЙрзЂЦкМфЮќШыЕФЧтЦјзюгааЇЕФХЈЖШЪЧ2%ЁЃУтвпШОЩЋКЭTUNELМьВтЯдЪОЃЌЧтЦјЧуЯђгкМѕЩйЦпЗњУбгеЕМЕФCC3КЭTUNELбєадЯИАћдіМгЃЈЭМ2a-cЃЉЁЃЮЊСЫБугкЧјЗжЕђЭіЯИАћЃЌНЋаЁЪѓНјвЛВНжУгке§ГЃПеЦјжа3аЁЪБЃЌвдЪЙДѓФджаЕФЕђЭіЕУвдЗЂеЙЃЈSevo/AirКЭSevo+H2/AirзщЃЉЁЃЮвУЧЙлВьЕНЃЌгыSevoзщЯрБШЃЌSevo/AirзщжаЦпЗњУбгеЕМЕФCC3КЭTUNELбєадЯИАћдМдіМгСЫСНБЖЃЈЭМ2a-cЃЉЁЃдкетаЉЪЕбщЬѕМўЯТЃЌ2%ЧтЦјЕМжТSevo+H2/AirзщЕФCC3КЭTUNELбєадЯИАћЪ§СПЯджјЕЭгкSevo/AirзщЃЈЭМ2a-cЃЉЁЃЮїЗНгЁМЃЗжЮіНвЪОЃЌЧтЦјвжжЦСЫЦпЗњУбдкаЈЧАЦЄжЪжагеЕМЕФCC3ЦЌЖЮдіМгЃЈЭМS2ЃЉЁЃЮвУЧНјвЛВНШЗШЯЃЌЫцзХ2%ЧтЦјЕФЪЙгУЃЌаЈЧАЦЄжЪжаЕђЭіЩёОзцЯИАћЕФЪ§СПМѕЩйЃЈЭМ1dЃЉЁЃ
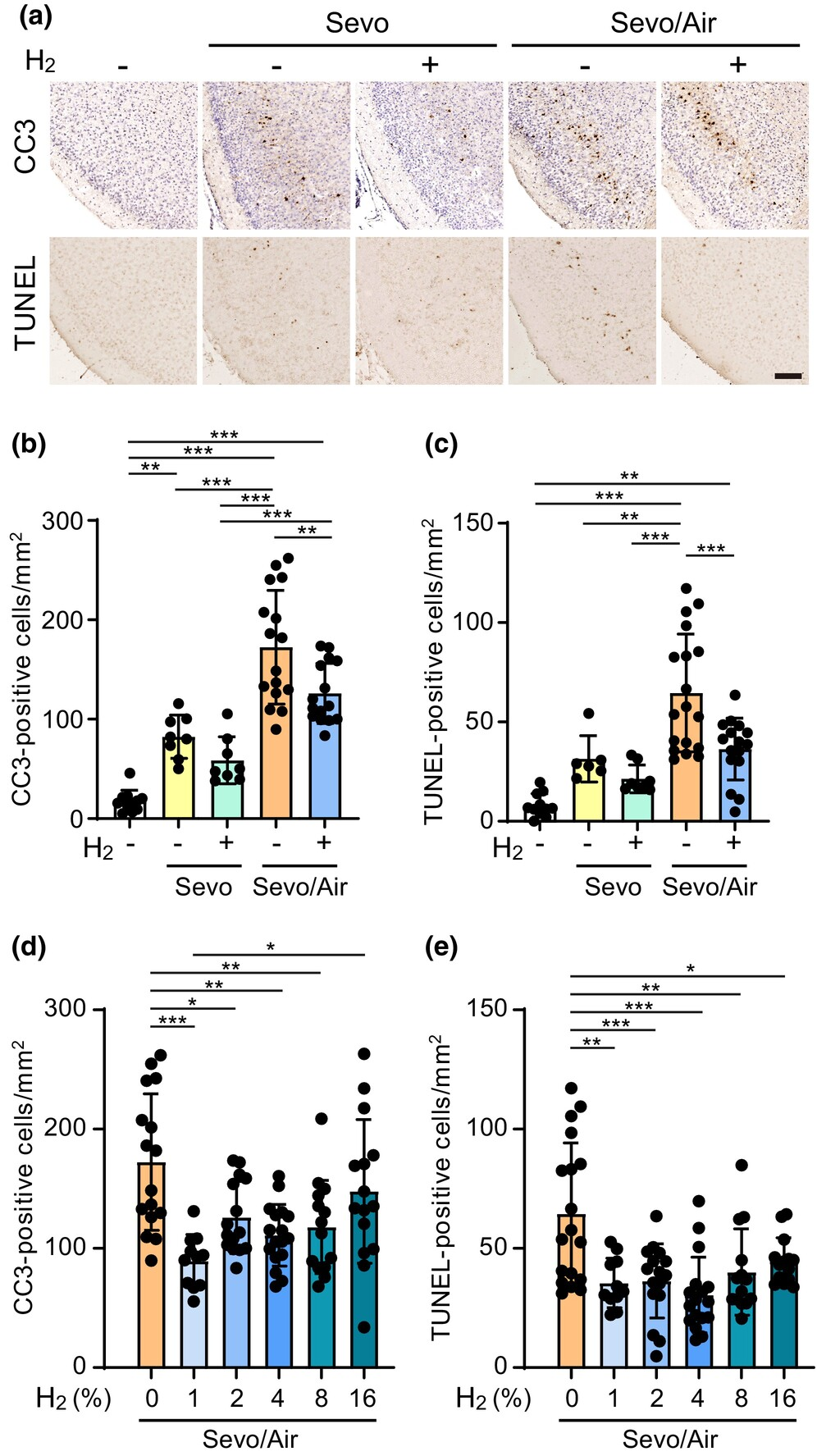
ЭМ2ЧтЦјдкаЈЧАЦЄжЪЖдПЙЦпЗњУбгеЕМЕФЕђЭіжаЗЂЛгЩёОБЃЛЄзїгУЁЃгзЪѓБЉТЖгкЛьКЯТщзэЦјЬхЃЈSevoЃЉ3аЁЪБЃЌгаЛђЮоЧтЦјЁЃЦфжавЛаЉгзЪѓНјвЛВНБЛЗХжУдке§ГЃПеЦјжа3аЁЪБЃЌвдЪЙЕђЭіЕУвдЗЂеЙЃЈSevo/AirЃЉЁЃЃЈaЃЉДњБэадЯдЮЂОЕЭМЯёЯдЪОЃЌгзЪѓБЉТЖгкКЌЛђВЛКЌ2%ЧтЦјЕФЛьКЯТщзэЦјЬхКѓЃЌаЈЧАЦЄжЪжаЦпЗњУбгеЕМЕФCC3бєадЯИАћгыКЫШОЩЋЫиИДШОЃЈЩЯЗНЃЉКЭTUNELбєадЯИАћЃЈЯТЗНЃЉЁЃБШР§ГпЃК100ІЬmЁЃЃЈb-eЃЉЖдБЉТЖгкКЌЛђВЛКЌ2%ЧтЦјЃЈb, cЃЉЛђКЌ0ЈC16%ЧтЦјЃЈd, eЃЉЕФЛьКЯТщзэЦјЬхЕФгзЪѓжаCC3бєадЯИАћЃЈb, dЃЉКЭTUNELбєадЯИАћЃЈc, eЃЉЕФЪ§СПНјааЖЈСПЗжЮіЁЃМЦЫуУПИігзЪѓЕФЙкзДЧаУцжааЈЧАЦЄжЪЕФбєадЯИАћЪ§СПЁЃИїзщЁАЖдееЁБЁЂЁАВЛКЌ2%ЧтЦјЕФSevoЁБЁЂЁАКЌ2%ЧтЦјЕФSevoЁБЁЂЁАВЛКЌ2%ЧтЦјЕФSevo/AirЁБЁЂЁАКЌ2%ЧтЦјЕФSevo/AirЁБЕФбљБОСПЗжБ№ЮЊЃКCC3 n=11, 8, 8, 16, КЭ15жЛгзЪѓЃЛTUNEL n=11, 6, 8, 18, КЭ16жЛгзЪѓЃЈb, cЃЉЁЃИїзщЁАВЛКЌЧтЦјЕФSevo/AirЁБЁЂЁАКЌ1%ЧтЦјЕФSevo/AirЁБЁЂЁАКЌ2%ЧтЦјЕФSevo/AirЁБЁЂЁАКЌ4%ЧтЦјЕФSevo/AirЁБЁЂЁАКЌ8%ЧтЦјЕФSevo/AirЁБЁЂЁАКЌ16%ЧтЦјЕФSevo/AirЁБЕФбљБОСПЗжБ№ЮЊЃКCC3 n=16, 11, 15, 17, 14, КЭ15жЛгзЪѓЃЛTUNEL n=18, 11, 16, 17, 14, КЭ14жЛгзЪѓЃЈd, eЃЉЁЃЪ§ОнБэЪОЮЊЦНОљжЕЁРБъзМВюЁЃ*p<0.05, **p<0.01, ***p<0.001ЃЛЕЅвђЫиЗНВюЗжЮігыTukeyМьбщЁЃ
НгЯТРДЃЌЮвУЧбаОПСЫдкSevo/AirзщжаЃЌФФжжХЈЖШЕФЧтЦјзюФмБЃЛЄЩёОзцЯИАћУтЪмЦпЗњУбгеЕМЕФЕђЭіЁЃЧтЦјЕФЪЙгУЯдЪОГіЧтЦјХЈЖШгыЕђЭіЯИАћЪ§СПжЎМфДцдкЗЧЯпадЕФUаЭЙиСЊЁЃЃЈетИіНсЙћЬсЪОЃЌЧтЦјЕФзїгУВЂВЛЪЧдНЖрдНКУЃЁЃЁЃЁЃЉПЙCC3ПЙЬхШОЩЋЯдЪОЃЌ1ЈC8%ЕФЧтЦјЯджјМѕЩйСЫCC3бєадЯИАћЕФЪ§СПЃЌЖј16%ЕФЧтЦјдђУЛгаЃЈЭМ2dЃЉЁЃЃЈМйШчетИіХЈЖШдкШЫРрвВЪЧШчДЫЃЌдђЪЧЧтЦјХЈЖШзюКЯРэЕФХЈЖШЪЧЕЭгк10%ЃЌАДееЭМжаЫљБэЪОЃЌРэЯыХЈЖШ1-4%ЃЌДѓдМУПЗжжг300КСЩ§ЃЌЖјВЛЪЧИќИпЩуШЁЧтЦјЁЃЫфШЛетжЛЪЧЦфжавЛИіжИБъЃЌУЛгаИќЖрЕФаХЯЂЃЌЩаВЛФмзїЮЊгІгУЕФВЮПМЁЃЕЋЧтНЁПЕВњвЕЙ§гкзЗЧѓИпСїСПЕФзіЗЈВЂВЛЪЧКЯРэЕФзіЗЈЁЃЃЉ
TUNELШОЩЋНвЪОЃЌ1ЈC16%ЕФЧтЦјЯджјМѕЩйСЫTUNELбєадЯИАћЕФЪ§СПЃЌЖј16%ЕФЧтЦјаЇЙћТдВюЃЈЭМ2eЃЉЁЃетаЉНсЙћБэУїЃЌдкЦпЗњУбгеЕМЕФДѓФдЫ№ЩЫКЭШБбЊдйЙрзЂЪЕбщжаЃЌ1ЈC8%ЕФЧтЦјОпгазюЧПЕФЩёОБЃЛЄзїгУЁЃ
3.3 ЧтЦјЮќЪеЭООЖМАЦфЖдбЊСїКЭбЊбѕЫЎЦНЕФгАЯь
НЋаТЩњаЁЪѓжУгкГфТњЧтЦјКЭЦпЗњУбЕФЧЛЪвжаЁЃЧтЦјМЋОпЩјЭИадЃЌвђДЫВЛНіЭЈЙ§КєЮќЮќЪеЃЌЛЙЭЈЙ§ЬхБэЮќЪеЁЃЮЊСЫжЄУїЧтЦјЪЧЭЈЙ§КєЮќЩуШыЕФЃЌЮвУЧЛЙНіНЋаЁЪѓЕФЩэЬхЛђЭЗВПжУгкЧЛЪвжаЃЌШЛКѓгУеыаЭЧтЦјЮЂДЋИаЦїВтСПИЮдржаЕФЧтЦјХЈЖШЃЈЭМ3aЃЉЁЃНіЭЗВПЛђШЋЩэБЉТЖгк2%ЧтЦјЕФаЁЪѓжаЃЌЧтЦјХЈЖШЯрЫЦЃЈЭМ3bЃЉЁЃВтЕУЕФХЈЖШдМЮЊ15mMЃЌетгы2%ЧтЦјдкЫЎжаШмНтЕФХЈЖШЗЧГЃЯрЫЦЃЌМД15.6mMЁЃЯрЗДЃЌНіДгОБВПвдЯТБЉТЖгк2%ЧтЦјЕФаЁЪѓЬхФкЧтЦјХЈЖШвЊЕЭЕУЖрЃКдМ3mMЁЃетаЉНсЙћБэУїЃЌМИКѕЫљгаЧтЦјЖМЪЧЭЈЙ§ЗЮКєЮќЩуШыЕФЁЃЮќШыЦјЬхжаЧтЦјХЈЖШЙщСуКѓЃЌЬхФкЧтЦјХЈЖШбИЫйНЕЕЭЃЈЭМ3cЃЉЁЃ
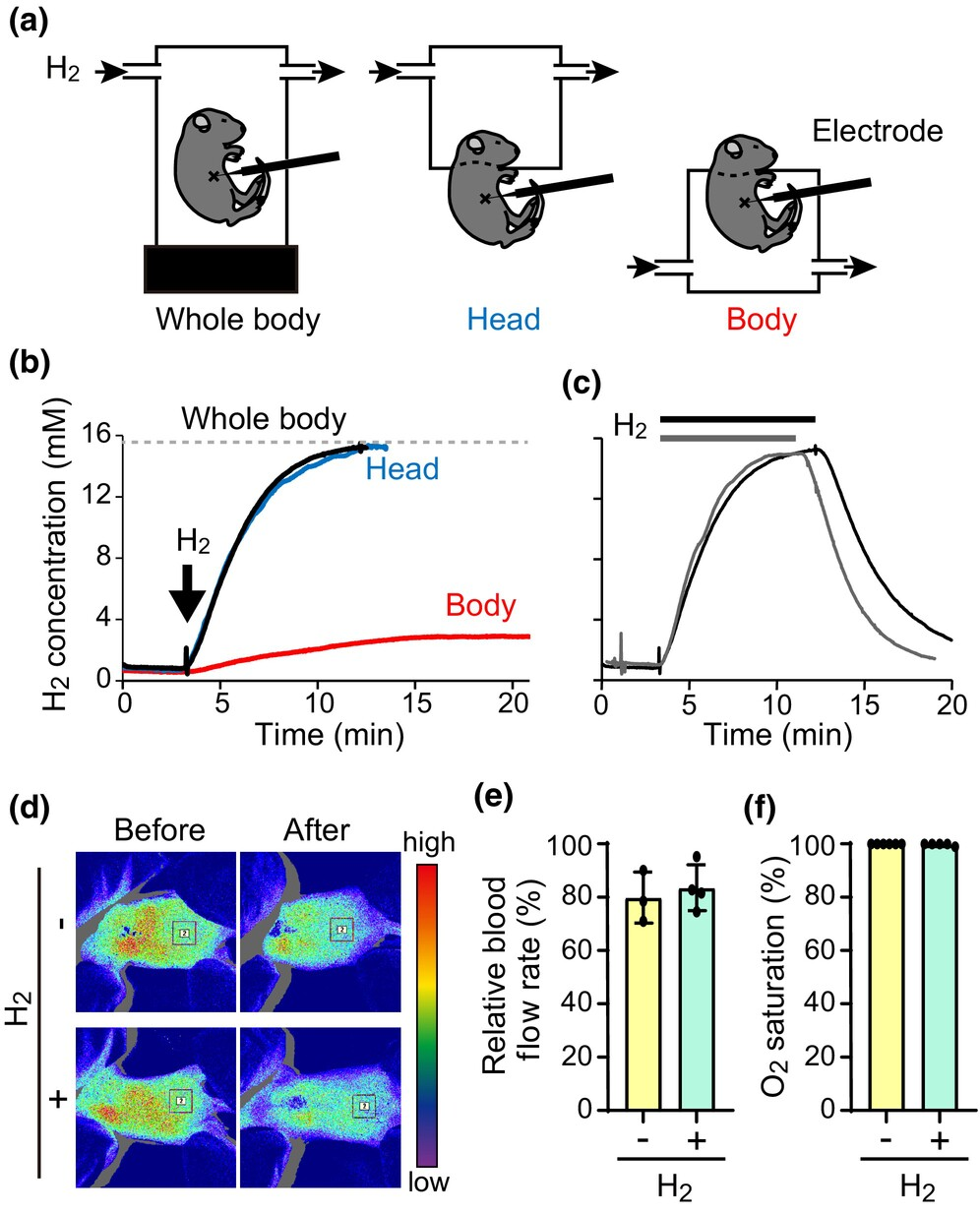
ЭМ3ЧтЦјЮќШыЖдбЊСїКЭбѕЦјЫЎЦНЕФгАЯьЁЃ(a-c) ДѓВПЗжЧтЦјЪЧЭЈЙ§КєЮќЖјВЛЪЧЭЈЙ§ЬхБэЮќЪеЕФЁЃ(a) БЉТЖВеЕФЪОвтЭМЃЌВеФкГфТњЦјЬхСїЃЌИВИЧећИіЩэЬхЁЂНіЭЗВПЛђНіЩэЬхЁЃ(b) дкУПИіВежаПЊЪМБЉТЖгк2%ЧтЦјЃЈМ§ЭЗЫљЪОЃЉКѓЃЌИЮдржаЧтЦјХЈЖШЕФЪБМфБфЛЏЁЃащЯпБэЪОГфТњ2%ЧтЦјЕФЫЎжаЧтЦјХЈЖШЁЃзЂвтЃЌЭЈЙ§ЬхБэЮќЪеЕФЧтЦјХЈЖШдіМгЛКТ§ЁЃ(c) ЭЃжЙЙЉгІЧтЦјЃЈДжЯпЫљЪОЃЉКѓЃЌИЮдржаЕФЧтЦјХЈЖШбИЫйНЕЕЭЁЃЯдЪОСЫСНДЮжиИДВтСПЕФНсЙћЁЃ(d, e) дкТщзэБЉТЖЦкМфЃЌбЊСїСПВЛЪм2%ЧтЦјЕФгАЯьЁЃЮќШыЦпЗњУбЧАКѓЃЌШЋЩэЕФМЄЙтЩЂАпЖдБШЭМЯёЃЌЮоТлЪЧЗёКЌгаЧтЦјЁЃДгУПеХЭМЯёЕФаЫШЄЧјгђПђжаЬсШЁбЊСїСПЖЈСПЗжЮіЪ§ОнЃЈdЃЉЁЃЮќШыЦпЗњУбКѓбЊСїСПЕФЖЈСПЗжЮіЃЌЮоТлЪЧЗёКЌга2%ЧтЦјЁЃбљБОСПШчЯТЃКЁАВЛКЌ2%ЧтЦјЕФЦпЗњУбЁБn=3жЛгзЪѓЃЌЁАКЌ2%ЧтЦјЕФЦпЗњУбЁБn=4жЛгзЪѓЃЈeЃЉЁЃ(f) дкТщзэЦкМфЃЌбѕЦјЫЎЦНВЛЪм2%ЧтЦјЕФгАЯьЁЃбљБОСПШчЯТЃКЁАВЛКЌ2%ЧтЦјЕФЦпЗњУбЁБn=6жЛгзЪѓЃЌЁАКЌ2%ЧтЦјЕФЦпЗњУбЁБn=5жЛгзЪѓЁЃЪ§ОнБэЪОЮЊЦНОљжЕЁРБъзМВюЁЃ
ФдЯИАћЕФДцЛюПЩФмЪмЕНФГаЉВЮЪ§ЕФгАЯьЃЌАќРЈбЊСїКЭбЊвКбѕЦјЫЎЦНЁЃЮЊСЫХХГ§ЮќШыЧтЦјгАЯьетаЉВЮЪ§ЕФПЩФмадЃЌЮвУЧВтСПСЫЮќШы2%ЧтЦјКЭЦпЗњУбЕФаЁЪѓЕФбЊСїСПЃЈЭМ3d,eЃЉКЭбЊвКбѕЦјЫЎЦНЃЈЭМ3fЃЉЁЃетаЉВЮЪ§ОљЮДЪмЕНЮќШы2%ЧтЦјЕФгАЯьЁЃ
3.4 ЧтЦјЖдЦпЗњУбМЄЛюЕФc-JunЭЈТЗОпгавжжЦзїгУ
ЮЊСЫВћУїЧтЦјЖдЦпЗњУбгеЕМЕђЭіБЃЛЄзїгУЕФЗжзгЛњжЦЃЌЮвУЧЪзЯШЭЈЙ§qPCRМьВщСЫЦЄВужаЛљвђБэДяЕФБфЛЏЁЃМгШыЦпЗњУбКѓЃЌFasЁЂFaslЁЂDdit3ЁЂPpp1r15aЁЂHmox1КЭJunЕФзЊТМЯджјдіМгЃЌЖјBcl2КЭEgr1ЕФзЊТМЯджјМѕЩйЃЈЭМ4aЃЛЭМS3ЃЉЁЃдкSevo/AirзщжаЃЌЧтЦјЯджјвжжЦСЫFasЁЂDdit3ЁЂHmox1КЭJunЕФдіМгзЊТМвдМАBcl2ЕФМѕЩйзЊТМЃЈЭМ4aЃЉЁЃетаЉЛљвђЪмc-JunЕФЕїПиЃЌc-Junгыc-FosНсКЯаЮГЩAP-1дчЦкЗДгІзЊТМвђзгЁЃc-JunЕФМЄЛюНјвЛВНгеЕМc-JunБОЩэЕФБэДяЁЃЪЕМЪЩЯЃЌМгШыЦпЗњУбКѓЃЌГВЕААзбєадЕФЩёОзцЯИАћВПЗжГЪЯжc-JunбєадЃЌЖјетжжЕААзБэДяБЛЧтЦјЫљвжжЦЃЈЭМ4bЃЉЁЃЮвУЧНјвЛВНМьВщСЫc-JunЕФСзЫсЛЏЁЃУтвпгЁМЃЯдЪОЃЌЧтЦјЧуЯђгкМѕЩйЦЄВужаЦпЗњУбгеЕМЕФc-JunСзЫсЛЏКЭЕААзжЪЗсЖШЕФдіМгЃЈЭМ4cЃЉЁЃ
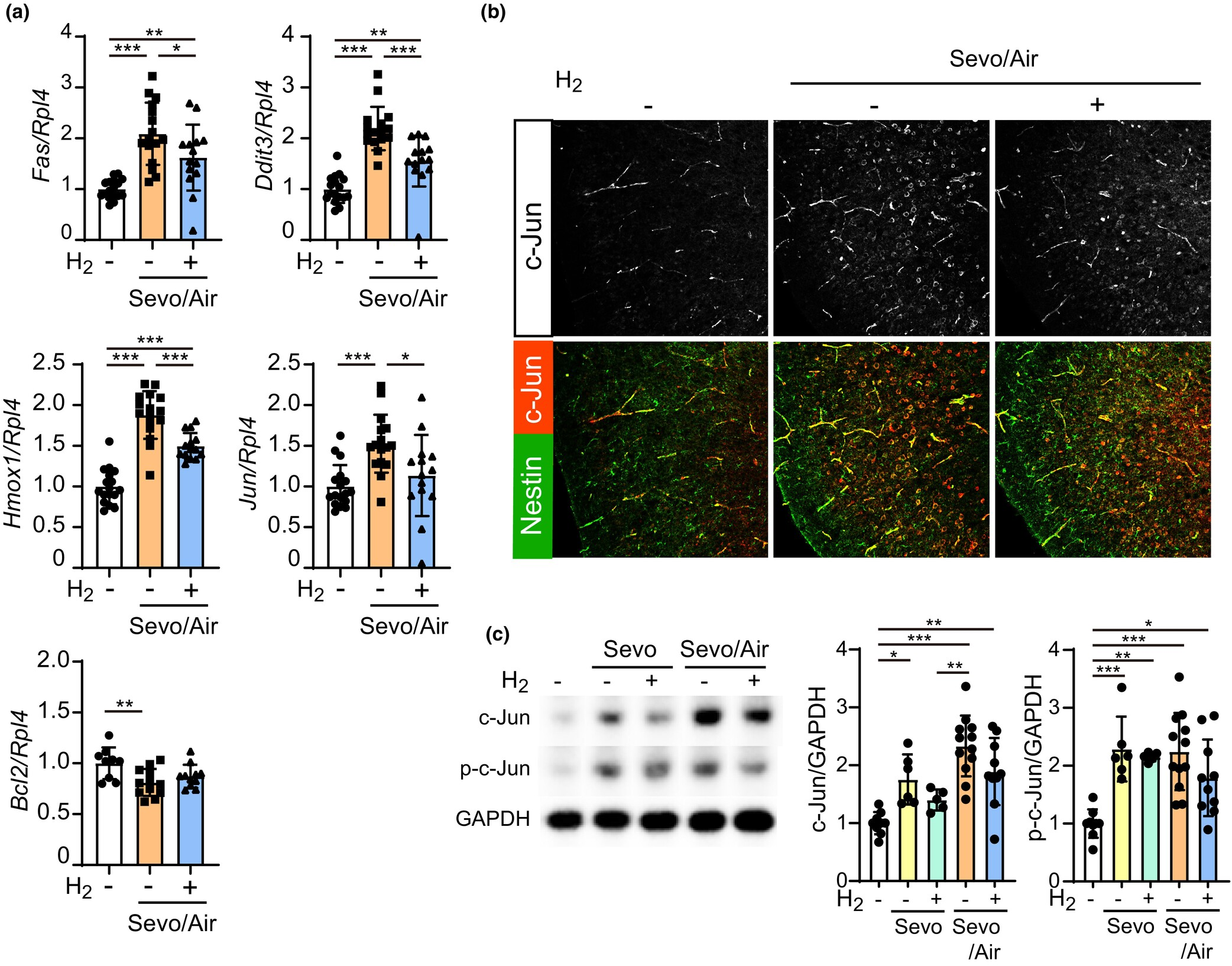
ЭМ4ЧтЦјЖдЦпЗњУбМЄЛюЕФc-JunЭЈТЗЕФгАЯьЁЃгзЪѓБЉТЖгкКЌгаЛђВЛКЌга2%ЧтЦјЕФЛьКЯТщзэЦјЬхЃЈSevoЃЉ3аЁЪБЃЌШЛКѓБЉТЖгкПеЦјжаСэЭт3аЁЪБЃЈSevo/AirЃЉЁЃ(a) ЧтЦјвжжЦСЫЦпЗњУбгеЕМЕФFasЁЂDdit3ЁЂHmox1КЭJunзЊТМдіМгвдМАBcl2зЊТММѕЩйЁЃДгаЈЧАЦЄВуЗжРыЕФзмRNAНјааСЫqPCRЗжЮіЁЃRpl4гУзїФкВЮЁЃИїФПБъЛљвђдкЁАЖдеезщЁБЁЂЁАВЛКЌ2%ЧтЦјЕФSevo/AirЁБКЭЁАКЌ2%ЧтЦјЕФSevo/AirЁБзщжаЕФбљБОСПШчЯТЃКFas n=17, 16, КЭ14жЛгзЪѓЃЛDdit3 n=17, 16, КЭ14жЛгзЪѓЃЛHmox1 n=17, 16, КЭ14жЛгзЪѓЃЛJun n=17, 16, КЭ14жЛгзЪѓЃЛBcl2 n=9, 12, КЭ11жЛгзЪѓЁЃ(b) ЙВОлНЙЯдЮЂОЕЯТЕФДњБэадЭМЯёЃЌЯдЪОаЈЧАЦЄВужагыПЙГВЕААзЙВШОЩЋЕФc-JunбєадЯИАћЁЃЧтЦјвжжЦСЫЩёОзцЯИАћжаЦпЗњУбгеЕМЕФc-JunБэДядіМгЁЃ(c) ЧтЦјЖдc-JunБэДяКЭСзЫсЛЏЃЈp-c-JunЃЉЕФгАЯьЁЃЪЙгУаЈЧАЦЄВуЕФСбНтвКНјааЮїЗНгЁМЃЗжЮіЁЃЭМБэЃЈгвВрЃЉЯдЪОСЫУПИіЬѕДјЕФЖЈСПЫЎЦНЁЃИЪгЭШЉ-3-СзЫсЭбЧтУИЃЈGAPDHЃЉгУзїФкВЮЁЃc-JunКЭp-c-JunЕФбљБОСПШчЯТЃКЁАЖдеезщЁБn=9ЃЌЁАВЛКЌ2%ЧтЦјЕФSevoЁБn=6ЃЌЁАКЌ2%ЧтЦјЕФSevoЁБn=5ЃЌЁАВЛКЌ2%ЧтЦјЕФSevo/AirЁБn=12ЃЌЁАКЌ2%ЧтЦјЕФSevo/AirЁБn=10жЛгзЪѓЁЃЪ§ОнБэЪОЮЊЦНОљжЕЁРБъзМВюЁЃ*p<0.05, **p<0.01, ***p<0.001ЃЛВЩгУTukeyМьбщЕФЕЅвђЫиЗНВюЗжЮіЁЃ
3.5 ЧтЦјвжжЦЦпЗњУбгеЕМЕФбѕЛЏгІМЄ
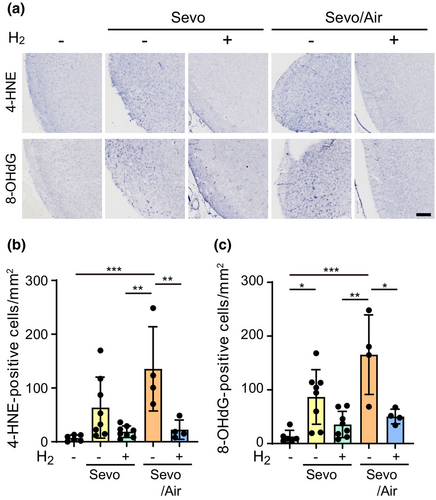
c-Jun NЖЫМЄУИЃЈJNKЃЉФЃПщдкЕђЭіжаЦ№зХживЊзїгУЃЌВЂЧвгЩбѕЛЏгІМЄМЄЛюЁЃДЫЭтЃЌЦпЗњУбЕФЩёОЖОадВПЗжЪЧгЩбѕЛЏгІМЄдіМгв§Ц№ЕФЁЃЮвУЧЙлВьЕНЃЌЦпЗњУбБЉТЖЃЈ3аЁЪБЃЉдіМгСЫаТЩњДѓФджаЕФжЌжЪЙ§бѕЛЏЃЈ4-HNEЃЉКЭбѕЛЏDNAЫ№ЩЫЃЈ8-OHdGЃЉЃЈЭМ5ЃЉЁЃ4-HNEКЭ8-OHdGбєадЯИАћЪ§СПЕФдіМгБЛЧтЦјЯджјНЕЕЭЃЌБэУїЧтЦјгеЕМЕФc-JunЭЈТЗМЄЛювжжЦжСЩйВПЗжПЩвдЭЈЙ§МѕЩйбѕЛЏгІМЄРДНтЪЭЁЃ
ЭМ5
ЧтЦјвжжЦЦпЗњУбгеЕМЕФбѕЛЏгІМЄЁЃгзЪѓБЉТЖгкКЌгаЛђВЛКЌга2%ЧтЦјЕФЛьКЯТщзэЦјЬхЃЈSevoЃЉ3аЁЪБЃЌШЛКѓБЉТЖгкПеЦјжаСэЭт3аЁЪБЃЈSevo/AirЃЉЁЃгУеыЖдбѕЛЏгІМЄБъМЧЮя4-HNEКЭ8-OHdGЕФПЙЬхЖдаЈЧАЦЄВуНјааУтвпзщжЏЛЏбЇШОЩЋЁЃЃЈaЃЉДњБэадЯдЮЂОЕЭМЯёЯдЪОСЫгУЫеФООЋКЫИДШОЕФ4-HNEКЭ8-OHdGбєадЯИАћЁЃБШР§ГпЃК100ІЬmЁЃЃЈb, cЃЉЖдБЉТЖгкКЌЛђВЛКЌЧтЦјЕФЛьКЯТщзэЦјЬхЕФгзЪѓжа4-HNEЃЈbЃЉКЭ8-OHdGЃЈcЃЉбєадЯИАћЕФЖЈСПЗжЮіЁЃМЦЫуУПИігзЪѓЕФаЈЧАЦЄВуЙкзДЧаУцжабєадЯИАћЕФЪ§СПЁЃ4-HNEКЭ8-OHdGЕФбљБОСПШчЯТЃКЁАЖдеезщЁБn=6ЃЌЁАВЛКЌ2%ЧтЦјЕФSevoЁБn=8ЃЌЁАКЌ2%ЧтЦјЕФSevoЁБn=8ЃЌЁАВЛКЌ2%ЧтЦјЕФSevo/AirЁБn=4ЃЌЁАКЌ2%ЧтЦјЕФSevo/AirЁБn=4жЛгзЪѓЁЃЪ§ОнБэЪОЮЊЦНОљжЕЁРБъзМВюЁЃ*p<0.05, **p<0.01, ***p<0.001ЃЛЕЅвђЫиЗНВюЗжЮігыTukeyМьбщЁЃ
3.6 ЧтЦјИФБфгЩЦпЗњУбаоЪЮЕФСзЫсЕААзжЪзщ
МјгкШЋЩэТщзэЕМжТДѓФджааэЖрЕААзжЪбИЫйСзЫсЛЏЃЌЕААзжЪСзЫсЛЏПЩФмЪЧаТЩњЖљаЁЪѓДѓФджаТщзэКЭЯИАћЕђЭіЛњжЦЕФЛљДЁЁЃШчЭМ2КЭЭМ5ЫљЪОЃЌ3аЁЪБЕФЦпЗњУбБЉТЖЧуЯђгкгеЕМЕђЭіКЭбѕЛЏгІМЄЃЌетаЉЭЈЙ§ИјгшЧтЦјВПЗжЪмЕНвжжЦЁЃЮЊСЫНвЪОЧтЦјЖдЕМжТЕђЭіЕФСзЫсЛЏБфЛЏЕФгАЯьЃЌЮвУЧЫцКѓбаОПСЫгыБЉТЖгк3%ЦпЗњУбЃЈКЌЛђВЛКЌ2%ЧтЦјЃЉ3аЁЪБЯрЙиЕФСзЫсЛЏЪТМўЃЌВЂгыЮДОДІРэЕФЖдееДѓФдНјааСЫБШНЯЁЃДгЦЄВуЬсШЁЕААзжЪВЂНјааДІРэгУгкФЩУзвКЯрЩЋЦз-жЪЦзСЊгУЗжЮіЃЈnano-LC-MS/MSЃЉЁЃдкНјааЭГМЦЗжЮіжЎЧАЃЌЖдАќКЌ5808ИіСзЫсыФЕФЪ§ОнНјааСЫЙ§ТЫЃЌвдБуНіАќРЈдкЫљгабљБОжаЗЂЯжЕФыФЁЃЮвУЧМјЖЈСЫ5789ИіСзЫсаоЪЮЮЛЕуЃЌЖдгІгк1962ИіЕААзжЪЁЃЫљгаЪ§ОнЕФЭъећФПТМПЩдкData S2жаЛёЕУЃЌВЂЭЈЙ§JPOSTКЯзїДцДЂПтдкProteomeXchangeСЊУЫЩЯДцЕЕЃЌЪ§ОнМЏБъЪЖЗћЮЊJPST002368ЁЃ
дкSevoзщжаЃЌга318ИіЕААзЯьгІЦпЗњУбЗЂЩњСЫСзЫсЛЏБфЛЏЃЌзмЙВга498ИіВювьСзЫсЛЏЪТМўЃЌАќРЈ257ИіГЌСзЫсЛЏКЭ241ИіЕЭСзЫсЛЏЪТМўЃЈЭМ6aЃЌFDR<0.2ЃЉЁЃдкSevo + H2зщжаЃЌга353ИіЕААзЯьгІЦпЗњУбЗЂЩњСЫСзЫсЛЏБфЛЏЃЌзмЙВга588ИіВювьСзЫсЛЏЪТМўЃЌАќРЈ301ИіГЌСзЫсЛЏКЭ287ИіЕЭСзЫсЛЏЪТМўЃЈЭМ6aЃЌFDR<0.2ЃЉЁЃДЫЭтЃЌЗжБ№га131КЭ166ИіЕААздкSevoзщКЭSevo + H2зщжаЬивьадЕиЗЂЩњВювьСзЫсЛЏЃЌЖј187ИіЕААздкСНзщжаЖМЗЂЩњСЫВювьСзЫсЛЏЃЈЭМ6bЃЉЁЃЪЙгУMetascapeЖдетаЉВювьСзЫсЛЏЕААзНјаазЂЪЭвдНјвЛВНЕїВщЫќУЧЕФЙІФмЃЈМћЭМS4ЃЌЪ§ОнS3ЃЉЁЃСНзщЙВгаЕФВювьСзЫсЛЏЕААззюЗсИЛЕФИЛМЏДиАќРЈЩёОЗЂг§ЁЂЭЛДЅаХКХКЭЯИАћМЋадЃЈЭМ6cЃЉЁЃетаЉНсЙћгыГЩФъДѓЪѓДѓФджавьЗњУбгеЕМТщзэЦкМфЕААзжЪСзЫсЛЏЕФБЈЕРЯрЫЦЁЃЖдНідкSevoзщКЭSevo + H2зщжаЕФВювьСзЫсЛЏЕААзНјвЛВНзЂЪЭЯдЪОЃЌзюЗсИЛЕФИЛМЏДиАќРЈЯИАћдЫЪфКЭжЪФЄЕїНкЃЈЭМ6d,eЃЉЁЃ
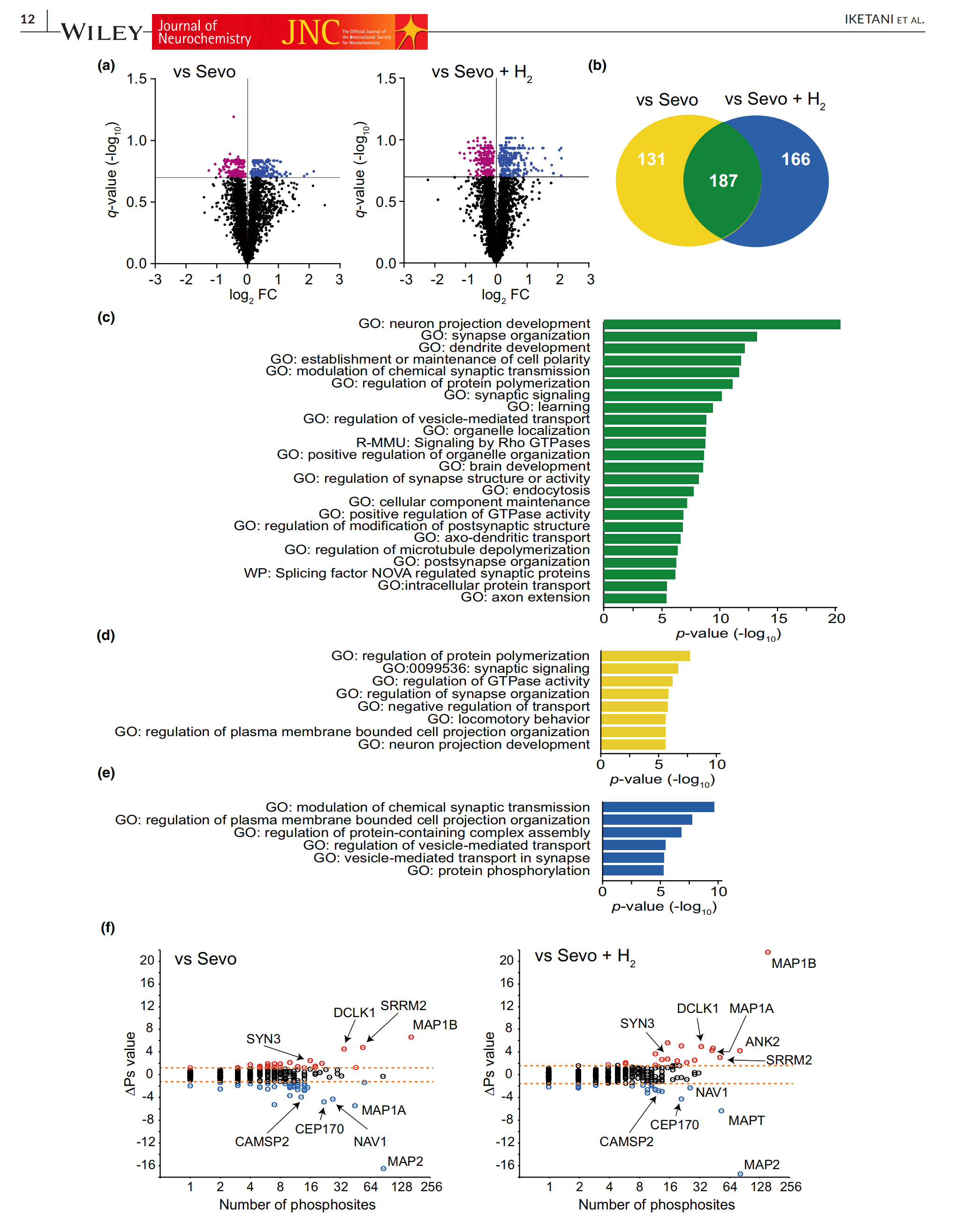
ЭМ6аЈЧАЦЄВуЯьгІЦпЗњУбЕФЖЈСПСзЫсЕААзжЪзщЃЌЮоТлЪЧЗёКЌгаЧтЦјЁЃгзЪѓЃЈn=24ЃЉЗжЮЊШ§зщЃКЖдеезщЃЈn=6ЃЉвдМАБЉТЖгкКЌ2%ЧтЦјЃЈn=9ЃЉЛђВЛКЌ2%ЧтЦјЃЈn=9ЃЉЕФЦпЗњУб3аЁЪБЁЃаЈЧАЦЄВуБЛНтЦЪВЂдкУПзщжаЗжЮЊШ§ИібљБОЁЃДгбљБОжаЬсШЁЕААзжЪВЂДІРэгУгкФЩУзвКЯрЩЋЦз-жЪЦзСЊгУЃЈnano-LC-MS/MSЃЉвдЖЈСПСзЫсЛЏыФЖЮЁЃЃЈaЃЉзмЙВЛцжЦСЫ5789ИіСзЫсЛЏаоЪЮЮЛЕуЁЃЛ№ЩНЭМЯдЪОСЫЖдеезщгыВЛКЌ2%ЧтЦјЕФЦпЗњУбзщЃЈvs. SevoЃЉЕФСзЫсЕААзжЪзщЃЌЯдЪОСЫ257ИіГЌСзЫсЛЏЃЈРЖЩЋЃЉКЭ241ИіЕЭСзЫсЛЏЃЈзЯЩЋЃЉЪТМўЃЌвдМАЖдеезщгыКЌ2%ЧтЦјЕФЦпЗњУбзщЃЈvs. Sevo + H2ЃЉЕФСзЫсЕААзжЪзщЃЌЯдЪОСЫ301ИіГЌСзЫсЛЏЃЈРЖЩЋЃЉКЭ287ИіЕЭСзЫсЛЏЃЈзЯЩЋЃЉЪТМўЃЈFDR < 0.2ЃЉЁЃЃЈbЃЉVennЭМЯдЪОЃЌдкSevoКЭSevo + H2зщжаЃЌга187ИіЕААзжЪдкСзЫсЕААзжЪзщЫЎЦНЩЯЗЂЩњСЫБфЛЏЃЌЖј131КЭ166ИіЕААзжЪЗжБ№жЛдкSevoКЭSevo + H2зщжаЗЂЩњСЫВювьСзЫсЛЏЁЃЃЈcЈCeЃЉЪЙгУMetascapeЖдВювьСзЫсЛЏЕААзНјааЛљвђзЂЪЭЃЈpжЕЃЈ−log10ЃЉ> 5ЃЉЁЃЃЈcЃЉдкSevoКЭSevo + H2зщжаЖМЗЂЩњСзЫсЛЏЕФбЁЖЈОЕфЭЈТЗЁЃЃЈd, eЃЉНідкSevoЃЈdЃЉКЭSevo + H2ЃЈeЃЉзщжаВювьСзЫсЛЏЕФбЁЖЈОЕфЭЈТЗЁЃЃЈfЃЉЖдеезщгыSevoзщжЎМфЃЈvs. SevoЃЛащЯпЃЌІЄPs = 1.23ЃЉвдМАЖдеезщгыSevo + H2зщжЎМфЃЈvs. Sevo + H2ЃЛащЯпЃЌІЄPs = 1.62ЃЉСзЫсЕААзЕФећЬхСзЫсЛЏзДЬЌБфЛЏЃЈІЄPsЃЉЗжЮіЁЃ
ДЫЭтЃЌЮвУЧШЋУцСПЛЏСЫСзЫсЛЏЕФБфЛЏЃЌетЪЙЕУФмЙЛМЦЫуЕЅИіЕААзжЪећЬхСзЫсЛЏзДЬЌБфЛЏЕФЕЅвЛжЕЃЈІЄPsЃЛМћЪ§ОнS4ЃЉЁЃдкSevoКЭSevo + H2зщжаЃЌЗжБ№га26КЭ18ИіЕААзжЪЯджјГЌСзЫсЛЏЃЌвдМА37КЭ23ИіЕААзжЪЯджјЕЭСзЫсЛЏЃЈ|ІЄPs| < 1.23КЭ1.62ЃЉЃЈЭМ6fЃЛЭМS5ЃЉЁЃСНзщжаГЌСзЫсЛЏзюЖрЕФЕААзжЪАќРЈЮЂЙмЯрЙиЕААзЃЈMAPЃЉ1BЁЂЫПАБЫс/ОЋАБЫсжиИДЛљжЪ2ЁЂЫЋЦЄжЪЫибљМЄУИ1КЭЭЛДЅЕААзIIIЁЃЕЭСзЫсЛЏЕААзАќРЈMAP2ЁЂжааФЬхЕААз170ЁЂЩёОдЊЕМКНЕААз1КЭИЦЕїЕААзЕїНкЕФЙтЦзЕААзМвзхГЩдБ2ЁЃЦфжавЛаЉЕААзВЮгыЯИАћЙЧМмЕААзНсКЯКЭЭЛДЅЪТМўЁЃгаШЄЕФЪЧЃЌMAP1AдкSevoзщжаЕЭСзЫсЛЏЃЌдкSevo + H2зщжаГЌСзЫсЛЏЁЃДЫЭтЃЌMAP1BдкSevo + H2зщжаЕФСзЫсЛЏГЬЖШдЖИпгкSevoзщЁЃЭЈЙ§Phos-tagММЪѕНјвЛВНШЗШЯСЫMAP1AКЭMAP1BдкH2зїгУЯТЕФдіЧПСзЫсЛЏЁЃдкЪЙгУPhos-tagЧэжЌЬЧбЁдёадИЛМЏаЈЧАЦЄВужаЕФСзЫсЕААзКѓЃЌЭЈЙ§ЮїЗНгЁМЃЗЈШЗЖЈСЫСзЫсЛЏЕФMAP1AКЭMAP1BЕААзЃЈЭМS6ЃЉЁЃЯрЗДЃЌMAP TauЕААзжЛдкSevo + H2зщжаЬивьадЕЭСзЫсЛЏЃЈЭМ6fЃЛЭМS5ЃЉЁЃ
4 ЬжТл
аэЖрбаОПБэУїЃЌЮќШыЧтЦјЖджЮСЦФдбЊЙмМВВЁЁЂЩёОЭЫааадМВВЁвдМААќРЈТщзэЦјЬхв§Ц№ЕФВЁРэБфЛЏдкФкЕФаТЩњЖљФдВПМВВЁгааЇЁЃЧтЦјЕФзїгУПЩФмжБНгвжжЦШЮКЮФдЫ№ЩЫЃЌвђЮЊжЮСЦаЇЙћдкЪмЩЫКѓМИаЁЪБФкОЭвбОЯдЯжЁЃШЛЖјЃЌЧтЦјЕФПЙбзКЭЦфЫћжЮСЦаЇЙћВЛФмЭЈЙ§жБНгЧхГ§ЛюадбѕЮяжжЃЈROSЃЉЕФФмСІРДНтЪЭЃЌвђЮЊЧтЦјЯдЪОГіЗЧГЃШѕЕФЗДгІадЃЈBuxtonЕШЃЌ1988ФъЃЉЁЃ
ЮвУЧШЗШЯЃЌдкаТЩњаЁЪѓДѓФджаЃЌОРњЦпЗњУбгеЕМЕђЭіЕФЯИАћЪЧаЈЧАЦЄВужаЕФГВЕААзбєадЩёОЧАЬхЯИАћЃЌетдкДѓЪѓФЃаЭжаБЛБЈЕРЬиБ№ЪмЕНШБбѕШБбЊЕФгАЯьЁЃаТЩњЦкЕФЦпЗњУбБЉТЖЛсЕМжТГЄЦкЗЂеЙЕФЩёОаФРэЙІФмеЯАЃЌАќРЈМЧвфеЯАЁЃЮќШыЧтЦјзшжЙСЫетаЉЮДГЩЪьЩёОдЊЕФЯИАћЫРЭіЁЃвдЧАЕФбаОПЗЂЯжЃЌАќРЈЦпЗњУбдкФкЕФШЋЩэТщзэвЉПЩФмЛсдкМИжжЯИАћРраЭжаЃЌЬиБ№ЪЧдкЗЂг§жаЕФДѓФджав§Ц№гРОУадЫ№ЩЫЁЃHofacerЕШШЫБЈИцЫЕЃЌЭЈЙ§НЋгзФъКЭФъЧсГЩФъаЁЪѓБЉТЖгквьЗњУбЃЌгеЕМСЫбЧПХСЃЧјАќРЈЭэЦкЧАЬхЯИАћКЭЮДГЩЪьПХСЃЯИАћдкФкЕФЮДГЩЪьЩёОдЊЕФбЁдёадЕђЭіЁЃНЋХрбјЕФаЁЪѓаТЦЄВуЩёОдЊКЭЧаЦЌБЉТЖгквьЗњУбЃЌвжжЦСЫгЩnetrinгеЕМЕФЩњГЄзЖЫњЯнЃЌВЂЦЦЛЕСЫжсЭЛЕМЯђЁЃNetrinМАЦфЪмЬхDCCКЭUnc5ВЮгыЩёОдЊЕМЯђКЭЕђЭіЁЃТщзэвЉПЩФмЭЈЙ§ИЩШХЩёОдЊГЩЪьЕФЙиМќЛЗНкРДгеЕМЩёОдЊЕђЭіЁЃЮвУЧМйЩшЧтЦјИФЩЦСЫетаЉаЇгІВЂБЃЛЄСЫЩёОЧАЬхЯИАћЁЃ
ЧтЦјИјвЉЯдЪОСЫЧтЦјХЈЖШгыФдЯИАћЕђЭіЪ§СПжЎМфЕФЗЧЯпадUаЭЙиСЊЃЛзюгааЇЕФЧтЦјХЈЖШЮЊ1-8%ЁЃжЎЧАгаБЈЕРГЦЃЌЧтЦјЖдЦпЗњУбгеЕМЕФЩёОдЊЕђЭіЕФБЃЛЄзїгУЪЧХЈЖШвРРЕадЕФЃЌДг0ЕН1.3%ЁЃЯрБШжЎЯТЃЌЧтЦјЖдДѓФдКЭаФдрШБбЊдйЙрзЂЫ№ЩЫЕФБЃЛЄзїгУЯдЪОГіUаЭЙиСЊЃЌзюгааЇЕФЧтЦјХЈЖШдМЮЊ2%ЃЌгыЮвУЧФПЧАЕФНсЙћЯрЫЦЁЃШЛЖјЃЌИќИпХЈЖШЕФЧтЦјВЂУЛгазшжЙЕђЭіЃЛЮЊЪВУДУЛгаЙлВьЕНБЃЛЄаЇЙћЩаВЛЧхГўЁЃвђДЫЃЌгІИУНјвЛВНбаОПЧтЦјЩёОБЃЛЄзїгУЕФЧБдкЛњжЦЁЃгаШЄЕФЪЧЃЌгаБЈЕРГЦЃЌУПЬьЮќШыСНДЮЧтЦјЃЌГжај4жмЃЌдіМгСЫХСН№ЩВЁЛМепФђвКжа8-OHdGЕФЫЎЦНЁЃ
ЧтЦјЃЈH2ЃЉЃЌвЛжжИпЩјЭИадЦјЬхЃЌПЩвдЭЈЙ§ЬхБэЮќЪеЁЃШЛЖјЃЌЮвУЧЕБЧАЕФбаОПБэУїЃЌЭЈЙ§ЮќШыЧтЦјЪЙЬхФкH2ХЈЖШдіМгЕФЫйТЪЃЌБШЭЈЙ§ЬхБэЮќЪевЊПьЕУЖрЁЃЕБЮќШыЦјЬхжаЕФH2ХЈЖШНЕжССуКѓЃЌЬхФкЕФH2ХЈЖШвВбИЫйЯТНЕЁЃетаЉНсЙћЬсЪОЃЌдкЪЕбщЯфФкЕФH2ЦјЬхжївЊЪЧгыЦпЗњУбвЛЦ№БЛЮќШыЃЌВЂгЩЗЮВПЮќЪеЁЃ
дкетЯюЪЕбщжаЃЌЮЊСЫАяжњШЗШЯЮќШыH2ЦјЬхВЛгАЯьЬхЮТЃЌЮвУЧНЋЪЕбщЯфФкВПБЃГждк37ЁуCЁЃгаБЈЕРГЦЃЌдкДѓЪѓаФЗЮжшЭЃФЃаЭжаЃЌЮќШыH2ЦјЬхВњЩњЕФФдБЃЛЄаЇЙћгыЕЭЬхЮТЯрЕБЁЃжЎЧАЕФвЛЯюбаОПЦРЙРСЫТщзэЧАКѓЖЏТібЊЕФЧщПіЃЌВЂжЄУїгыЦпЗњУбЭЌЪБИјгш1.3% H2ЖдpHжЕЁЂбѕЦјЗжбЙЛђЖўбѕЛЏЬМЗжбЙУЛгаЯджјВЛСМгАЯьЁЃДЫЭтЃЌH2ЕФЭЌЪБИјвЉдкТщзэКѓЮДв§Ц№ШЮКЮааЮЊвьГЃЁЃЮвУЧЛЙЙлВьЕНЃЌгыНігУЦпЗњУбТщзэЕФаЁЪѓЯрБШЃЌгУ2% H2КЭЦпЗњУбТщзэЕФаЁЪѓдкбЊСїЫйТЪЛђбЊбѕЫЎЦНЩЯУЛгаУїЯдБфЛЏЁЃетаЉНсЙћБэУїЃЌЮќШыH2ВЛгАЯьЦпЗњУбЕФТщзэаЇЙћЃЌЧвЩЯЪіЩњРэВЮЪ§ВЛЩцМАH2геЕМЕФЩёОБЃЛЄЛњжЦЁЃ
ЮвУЧеЙЪОСЫЦпЗњУбТщзэгеЕМСЫЦЄВужаc-JunПижЦЯТЕФЛљвђБэДяЃЌЦфжавЛаЉЛљвђЭЈЙ§ЭЌЪБЮќШыH2ЖјБЛЯТЕїЁЃдкаТЩњДѓЪѓКЃТэжаЃЌJNK/c-Jun/AP-1ЭЈТЗПЩвдБЛЦпЗњУбМЄЛюЃЌЫцКѓгеЕМСЌНгЕААз43ЕФБэДяЁЃвђДЫЃЌЮвУЧМьВщСЫc-JunЕФСзЫсЛЏЧщПіЃЌЗЂЯжH2МѕЩйСЫЦпЗњУбгеЕМЕФСзЫсЛЏдіМгЁЃc-JunЕФМЄЛюгеЕМСЫc-JunБОЩэЕФБэДяЁЃДЫЭтЃЌЦпЗњУбдіМгСЫc-Jun mRNAКЭЕААзжЪЫЎЦНЃЌЖјетжждіМгБЛH2ЯджјНЕЕЭЁЃетаЉНсЙћБэУїЃЌвжжЦгыc-JunМЄЛюЯрЙиЕФаХКХЭЈТЗПЩФмВЮгыСЫH2ЕФПЙЕђЭіЛюадЁЃгІНјвЛВНЕїВщвдВћУїАќРЈJNKКЭAP-1дкФкЕФc-JunЩЯЯТгЮЪЧЗёЪмЕНH2ЕФгАЯьЁЃ
JNK/c-Jun/AP-1ЭЈТЗЕФМЄЛюЖдгкгЩЖржжЩЫКІв§Ц№ЕФЩёОдЊЕђЭіжСЙиживЊЃЌАќРЈбѕЛЏгІМЄЁЃШЛЖјЃЌЙигкЮќШыадТщзэвЉв§Ц№ЕФбѕЛЏгІМЄЕФбаОПНсЙћЪЧгаељвщЕФЁЃвЛЯюбаОПБЈИцГЦЃЌЦпЗњУбдіМгСЫГЩФъДѓЪѓЗЮВПЕФБћЖўШЉЃЌЕЋМѕЩйСЫЩідрКЭДѓФджаЕФБћЖўШЉЁЃЯрЗДЃЌСэвЛЯюбаОПБЈИцГЦЃЌЦпЗњУбдіМгСЫаТЩњДѓЪѓКЃТэжаЕФROSЫЎЦНКЭбѕЛЏDNAЫ№ЩЫЁЃДЫЭтЃЌвЛЯюЖдНгЪмФђЕРЯТСбаоИДЪжЪѕЕФгзЖљЃЈ12-36ИідТДѓЃЉЕФбаОПЗЂЯжЃЌЪжЪѕ2аЁЪБКѓЦпЗњУбдіМгСЫПЙбѕЛЏУИЕФmRNAЫЎЦНЃЌБэУїЦпЗњУбднЪБдіМгСЫЖљЭЏЕФбѕЛЏгІМЄЁЃЮвУЧЕФЪ§ОнЯдЪОЃЌБЉТЖгкЦпЗњУб3аЁЪБдіМгСЫаТЩњДѓФджаЕФжЌжЪЙ§бѕЛЏКЭбѕЛЏDNAЫ№ЩЫЁЃгаШЫЬсГіЃЌгЩгкЦпЗњУбЕМжТЕФЩёОдЊЗХЕчБЌЗЂдіМгСЫДњаЛашЧѓЃЌетПЩвдбИЫйКФОЁОжВППЩгУЕФбѕЦјЃЌвђЮЊШЋЩэТщзэИЩШХСЫе§ГЃЕФЩёОбЊЙмёюКЯВЂгеЗЂСЫбѕЛЏгІМЄЁЃШчКѓЫљЪіЃЌТщзэв§Ц№ЕФЮЂЙмЕїНкЕМжТЕђЭіЃЌвВПЩФмдіЧПСЫДЅЗЂJNKКЭЕђЭіаХКХЗДгІЕФROSВњЩњЁЃжЕЕУзЂвтЕФЪЧЃЌбѕЛЏгІМЄЕФдіМгБЛH2ЮќШыЛКНтСЫЁЃШЛЖјЃЌH2ЖдROSЕФЛЙдЛюадЬЋЕЭЃЌЮоЗЈжБНгвжжЦбѕЛЏгІМЄЁЃ
ЕААзжЪСзЫсЛЏЪЧвЛжжживЊЁЂПьЫйЧвЫВЪБЕФЗвыКѓаоЪЮЃЌЫќЕїНкзХаэЖрЩњЮябЇЙ§ГЬЃЌЖјЖЈСПСзЫсЕААззщбЇе§ГЩЮЊбаОПШЋОжаХКХЫЎЦНБфЛЏЕФЗЧГЃгааЇЕФЙЄОпЁЃгабаОПБэУїЃЌЕААзжЪСзЫсЛЏПЩФмЪЧТщзэМСМАЦфЫцКѓдкДѓФджав§Ц№ЕФЩёОдЊЫ№ЩЫЕФПьЫйЛњжЦЕФЛљДЁЁЃЖдБЉТЖгквьЗњУбЕФГЩФъаЁЪѓДѓФдНјааЕФСзЫсЕААззщбЇбаОПЃЌШЗЖЈСЫТщзэМСЕФжївЊвЉРэбЇАаЕувдМАгыЩёОдЊаЫЗмадЁЂДѓФдФмСПЮШЬЌЁЂЭЛДЅПЩЫмадКЭДЋЕнМАЮЂЙмЙІФмЯрЙиЕФЦфЫћМИжжЕААзжЪзїЮЊТщзэМСЕФДЮвЊМйЖЈАаЕуЁЃЯИАћЙЧМмжиЫмКЭЭЛДЅаХКХЪТМўжаЩцМАЕФЕААзжЪЕФПьЫйЗвыКѓаоЪЮПЩФмНщЕМШЋЩэТщзэМСЕФжааФЛњжЦЁЃ
дкЮвУЧЕФбаОПжаЃЌБЉТЖгкЦпЗњУбЕФаТЩњаЁЪѓДѓФджаВювьСзЫсЛЏЕААзжЪЕФзюЗсИЛМЏШКАќРЈЩёОЗЂеЙКЭЭЛДЅаХКХДЋЕМЃЌетБэУїТщзэМСЭЈЙ§ИФБфЩёОдЊНсЙЙКЭЙІФмЗЂЛгзїгУЁЃЭЛДЅаЮГЩЪЧЮДГЩЪьЩёОдЊДцЛюЕФЙиМќВНжшЃЌвђЮЊРДздФПБъЕФвђзгДгЭЛДЅдЫЪфЕНАћЬхЃЌгАЯьЕђЭіаХКХЁЃдкSevoЛђSevo + H2зщжаЕФВювьСзЫсЛЏЕААзжЪЯдЪОЃЌзюЗсИЛЕФМЏШКАќРЈЯИАћдЫЪфКЭЯИАћЭЛГізщжЏЁЃЪЕМЪЩЯЃЌдкЬхЭтБЉТЖгкH2ЕФЩёОФИЯИАћСіжаЃЌФкЬхдЫЪфЕНЛиЪеФкЬхЕФбгГйвбБЛЙлВьЕНЁЃМгЫйЕФЯИАћФкдЫЪфЖдгкЕђЭіжСЙиживЊЃЌЕБЯИАћФкдЫЪфБЛЕїНкЛие§ГЃЫЎЦНЪБЃЌЕђЭіЛсБЛбгГйЁЃвђДЫЃЌЮвУЧЭЦВтH2ЭЈЙ§ЕїНкЯИАћдЫЪфРДМѕЧсЦпЗњУбгеЕМЕФЕђЭіЁЃ
ЯИАћФкдЫЪфвРРЕгквЛИіИпЖШзщжЏЛЏЕФЯЕЭГЃЌАќРЈЮЂЙмКЭЮЂЙмЯрЙиЗжзгТэДяЕААзЁЃаэЖрЗЂЯжБэУїЃЌЮЂЙмЭјТчдкЕђЭіЙ§ГЬжаЛсжиаТаЮГЩЃЌВЂажњКЫКЭЯИАћЫщЦЌЕФЗжЩЂЁЃЮЌГжЕђЭіЮЂЙмЭјТчвРРЕгкИпATPЫЎЦНКЭЛюдОЕФЯпСЃЬхЃЌЯпСЃЬхФЄЕчЮЛЕФЯћЩЂЕМжТГЌбѕЛЏЮявѕРызгЕФЙ§ЖШВњЩњКЭROSЫЎЦНЕФдіМгЁЃдкЮвУЧЕФЪЕбщжаЃЌШЋУцЖЈСПЕФСзЫсЛЏБфЛЏНвЪОЃЌМИИіЦпЗњУбгеЕМЕФЙ§ЖШКЭЕЭСзЫсЛЏЕААзгыЮЂЙмЭјТчКЭЯИАћЙЧМмжиЫмгаЙиЃЌетПЩФмЪЧТщзэМСКЭЮДГЩЪьЩёОдЊЕђЭіаЇгІЕФЛљДЁЁЃЮвУЧЗЂЯжЃЌH2ЮќШыНјвЛВНдіМгСЫMAP1AКЭMAP1BЕФСзЫсЛЏЁЃMAP1МвзхГЩдБИљОнЦфСзЫсЛЏзДЬЌКЭдкЩёОЗЂг§МАећЬхЮЌЛЄжаЕФЙІФмЃЌПЩФмдкЕїНкЮЂЙмЯИАћЙЧМмжаЗЂЛгЖржизїгУЁЃЫЋЬивьадРвАБЫсСзЫсЛЏЕїНкМЄУИ-1aЃЈDYRK1aЃЉПЩвдСзЫсЛЏMAP1AКЭMAP1BЃЌРвЕААзМЄУИ2ПЩвдСзЫсЛЏMAP1AЁЂMAP1BКЭУЊЕААз-2ЁЃгаШЄЕФЪЧЃЌDYRK1aвбБЛжЄУїдкЗЂг§жаЕФЪгЭјФЄКЭМИжжШЫРрАЉЯИАћЯЕжавжжЦФкдкЕђЭіЭООЖЁЃРвЕААзМЄУИ2ЕФЙ§БэДяДйНјаЧаЮНКжЪЯИАћдіжГЃЌвжжЦЩёОдЊЕђЭіЃЌВЂМѕЩйДѓЪѓДѓФджаЕФаЧаЮНКжЪЯИАћНщЕМЕФбзжЂЁЃетаЉМЄУИЕФМЄЛюКЭMAP1МвзхГЩдБЕФСзЫсЛЏПЩФмВЮгыСЫТщзэЦкМфH2ЕФПЙЕђЭіЛюадЁЃ
5 НсТл
ЮвУЧЕФЪ§ОнБэУїЃЌдкТщзэЦкМфИјгшH2ПЩЭЈЙ§ЮЂЙмЕФЙ§ЖШСзЫсЛЏРДЕїНкЯИАћдЫЪфЁЃетПЩФмвжжЦROSЕФВњЩњВЂМѕЧсЩёОЧАЬхЯИАћЕФЕђЭіЁЃЮвУЧБиаыПМТЧЕНЃЌНЯИпХЈЖШЃЈ> 4%ЃЉЕФH2ЪЧвзШМЕФЃЌЦфжївЊАаЗжзгШдШЛЮДжЊЁЃШЛЖјЃЌИјгш1-4%ЕФH2ЦјЬхПЩвдШнвзЧвАВШЋЕиМѕЧсДѓФджаТщзэгеЕМЕФЕђЭіЁЃДЫЭтЃЌдкЖљПЦТщзэжаЪЙгУH2ПЩФмЛсЬсИпЕБЧАШЋЩэТщзэЗНАИЕФАВШЋадЁЃ
https://blog.sciencenet.cn/blog-41174-1437488.html
ЩЯвЛЦЊЃК2аЭЬЧФђВЁЛМепФкдДадВњЧтЦјЯТНЕ
ЯТвЛЦЊЃКФёблЗТЩњШЫЙЄЪгОѕЯЕЭГ
