博文
氢气增强短跑间隔训练的表现:代谢组学洞察其潜在机制
||
氢气被认为可以通过减少高强度运动引起的氧化应激来缓解疲劳并减轻运动性能的下降。然而,用于评估氢气抗氧化能力的血液标志物的多样性限制了对其机理效应的全面理解。本研究使用代谢组学方法评估了富含氢气的气体(HRG)对短跑间隔训练后代谢物的影响,旨在阐明其作用的潜在机制。结果:与安慰剂(空气)相比,吸入HRG显著提高了第四次冲刺的平均功率、疲劳指数和达到峰值的时间,并显著减少了第一次和第四次之间峰值功率、平均功率和达到峰值时间的衰减值。代谢组学分析突出显示,在HRG吸入后,乙酰肉碱、丙酰-L-肉碱、次黄嘌呤和黄嘌呤显著上调,富集途径分析表明HRG可能通过增强辅酶A合成、促进甘油磷脂代谢和抑制胰岛素水平来促进脂肪动员。结论:在短跑间隔测试前吸入HRG可增强最终阶段的无氧冲刺能力和缓解疲劳。代谢组学分析表明HRG可能在休息期间加速脂肪氧化,并为后续冲刺提供更大的ATP补充,通过减轻线粒体氧化损伤、提高有氧效率和刺激脂肪动员来实现。

高强度运动导致活性氧物种(ROS)大幅增加,超过身体的清除能力并打破氧化-抗氧化平衡,从而诱发氧化应激[1, 2]。超出的ROS还可能影响线粒体功能[3],诱导电子传递和ATP合成的减少,降低有氧途径的效率,增加无机磷酸盐和乳酸的水平[4],并破坏一个或多个参与兴奋-收缩偶联的蛋白质,导致肌肉力量产生减少[5]。这些氧化应激的生物生理变化是疲劳的主要原因之一[5],因此会影响运动表现[6]。因此,迫切需要减少过量的ROS从而提高运动表现。
氢具有选择性清除对身体有害的自由基的特性[7]。研究表明,使用富含氢的水和富含氢的气体(HRG)对缓解氧化应激和运动引起的疲劳有益[8–11]。然而,评估氢机制的现有研究在选择血液生物标志物方面各不相同,大多数标志物与氢效应无关[9, 10],缺乏全面的筛选以识别与氢作用机制相关的靶化合物。迄今为止,没有研究揭示了与HRG在运动期间及其与体能表现相关的完整代谢物变化,使得HRG可能改善运动表现的机制仍然广泛未被探索。
代谢组学技术能够获取整个生物体的代谢状态的高吞吐量“快照”[12–14],通过分析运动期间内源性代谢物的变化,为功能性能调节背后的机制提供全面的洞察[15]。代谢组学也被广泛用于测量补充剂和药物引起的代谢物变化[16, 17]。然而,氢如何通过代谢组学机制诱导运动表现的益处?这一方面尚未得到明确和全面的检查。
因此,我们在这里旨在通过使用代谢组学技术来探索HRG能够增强运动表现和减轻疲劳的可识别代谢物和代谢途径。我们使用短跑间隔训练来诱导氧化应激,这种协议可以引发与高强度间隔训练或连续中等强度训练相当或更大的全身氧化还原状态反应,尽管总工作量较低[18]。这项工作获得的知识可以增进对氢气改善运动表现和缓解疲劳益处机制的理解。
2 方法
2.1 参与者
本研究招募了10名健康、休闲活跃的成年男性(23.6 ± 2.2岁,179.4 ± 5.9厘米,76.5 ± 7.3公斤)。纳入标准是:参与者每周至少进行2小时的训练习惯化(包括抗阻训练),并有能力完成高强度的自行车测功仪练习(在初步筛查访问期间能够完成Wingate测试)。排除标准是:实验前六个月内没有下肢受伤史,没有心血管、呼吸和内分泌疾病。在详细解释了实验程序后,每位参与者都签署了知情同意书。研究方案符合《赫尔辛基宣言》,并得到了北京体育大学(BSU)运动科学实验伦理委员会的批准(批准号:2021107H)。
2.2 协议
在这项双盲、平衡、随机、交叉设计的研究中,参与者在北京体育大学的实验室完成了两次研究访问,每次访问他们使用Wingate功率计(Monark 894E, 瑞典)完成短跑间隔测试(SIT),在此之前他们吸入了富含氢的气体(HRG)(这次访问记录为HG)或安慰剂气体(标准空气)(这次访问记录为PG)。研究访问的顺序是随机的。具体来说,访问顺序是根据由基于网络的计算机程序(www.randomizer.org)生成的平衡排列随机分配的。这两次访问之间提供了至少7天的洗脱期。
在这两次访问中,为了进行代谢组学分析,从参与者那里在三个时间点采集了肘静脉血样:吸入气体前(pre-gas)和吸入气体后立即(post-gas),以及SIT后立即(post-SIT)(图1)。所有参与者都被指示在每次试验期的48小时内避免剧烈运动、酒精、咖啡、补充剂、药物和任何特定的恢复治疗。在每次研究访问的早晨,所有参与者都消费了研究人员提供的标准餐,包括牛奶、面包和火腿肠。此外,他们在访问前不摄入任何额外的食物或饮料。
2.3 富含氢气的气体(HRG)
HRG由中国福州智恒氢健康科技有限公司生产的氢气发生器Hydrogen-oxygen Convalescent machine 2.5制备。该发生器可以产生30ml/s的氢氧混合气(氢气和氧气的比例为2:1)。HRG通过连接在气体发生器上的鼻导管供应。虽然由于技术限制我们无法直接测量进入体内的氢气和氧气浓度,但根据数学估计,健康年轻男性在安静状态下的平均吸气流速约为500 ml/s,这远远超过了氢气发生器的流速,稀释了吸入氢气的浓度,使得体内吸入的氢气最大浓度约为4.08%。同样,氧气浓度最多为21.57%[10]。与空气中的氧气含量相比,增加的氧气极其微小。安慰剂气体(环境空气,0.00005%氢气,20.9%氧气)通过连接到未启动制氢程序的氢气发生器的鼻导管供应。
2.4 短跑间隔测试(SIT)
短跑间隔测试用于检查HRG对短跑间隔性能的影响(一种扰乱身体氧化还原平衡的运动形式)。参与者需要完成四次30秒的全力短跑,分别称为第一次冲刺(S1)、第二次冲刺(S2)、第三次冲刺(S3)和第四次冲刺(S4),每次之间间隔4.5分钟(图1)。参与者在正式测试前需要进行5分钟的热身。参与者以全力蹬车,阻力在3秒内增加到规定负荷(0.075 × 体重(kg))。测试期间,给参与者口头鼓励。此外,要求参与者在测试期间避免将臀部抬离自行车座椅。之前的研究已经证明这种方法有效地扰乱了全身的氧化还原状态[18]。平均功率(MP)是主要的结果指标,反映了冲刺骑行期间的整体表现。此外,还记录了每次冲刺骑行的峰值功率(PP)、疲劳指数(FI,(最大功率 - 最小功率) / 最大功率 * 100%)和达到峰值时间(TTP)。考虑到氢气的作用直到高强度运动后活性氧种类(ROS)大幅上升时才会显现,我们通过S4的结果(氢气作用后效果)和S1与S4之间的衰减值(ΔS4-S1)来评估其影响。S4结果反映了高强度运动后的无氧性能,而ΔS4-S1结果反映了短跑间隔训练期间的疲劳和恢复能力。
2.5 血液代谢组学
2.5.1 血液样本的采集和处理
从参与者的前臂静脉采集约5mL血液,立即在液氮中冷冻,然后储存在-80°C下等待分析检测。在测试前,将200µL血浆与800µL色谱纯甲醇在1.5mL离心管中涡旋混合60秒,然后在-20°C冷却1小时。接着在4°C下以15,000rpm离心10分钟。提取900µL上清液,在4°C下真空离心干燥。加入90µL含有氘代内标的色谱纯甲醇,涡旋1分钟,然后加入90µL超纯水,再涡旋1分钟。在4°C下以15,000rpm离心10分钟,将上清液转移到新管中进行测量。最后,取出5µL上清液放入1.5mL离心管中,涡旋60秒进行质量控制(QC)。
为了全面捕捉代谢物信息,将使用正负离子模式进行数据采集,主要采用飞行时间扫描(TOF-SCAN)作为主要的收集方法。此外,也将使用这些离子模式分析QC样本,并通过自动串联质谱(Auto-MS/MS)进行迭代收集。使用TOF-SCAN旨在监测整个样品处理过程中仪器的稳定性,而Auto-MS/MS则用于获得化合物的详细碎片离子信息,这对于数据分析时的准确定性确认至关重要。在正离子模式下,使用C18柱(2.1×150 mm, 3.5 µm),柱温设置为50°C。流动相A为水中的0.1%甲酸,流动相B为乙腈中的0.1%甲酸。流速保持在0.35 mL/min。对于负离子模式,使用T3柱(2.1×100 mm, 1.8 µm),柱温也为50°C。在这里,流动相A是水中的6.5 mmol/L碳酸氢铵,流动相B是95%甲醇-水的6.5 mmol/L碳酸氢铵混合物。该模式下的流速同样设置为0.35 mL/min。这些参数经过精心选择,以优化每种电离模式下代谢物的分离和检测。
2.5.2 非靶向代谢组学
在我们的代谢组学研究中,我们首先使用MassHunter进行分子特征提取(MFE)处理全扫描模式下检测到的原始代谢物数据,然后将数据以CEF格式导出。接着,我们将这些数据导入Mass Profiler Professional(MPP)软件。在MPP中,我们通过对加合离子类型和模式、同位素类型、化合物电荷状态、保留时间、质量范围、质量过滤参数、信噪比和最小峰高等参数进行设置来对齐数据。为了提高数据分析的质量,我们在特定的递归条件下重新识别了离子。
鉴于代谢组学数据的高维度和变量间的相关性,传统的单变量分析在提取潜在信息方面效率低下且不准确。因此,我们采用了如主成分分析(PCA)等多变量统计方法来降低维度并对多维数据进行回归分析,力求保留最大的原始信息。
对于所有血液代谢物,我们使用MPP软件根据化合物标记(Filter Flags)和出现频率(Filter By Frequency)依次筛选差异化合物。我们利用PCA来评估样本质量并探索血液样本中代谢物的差异。正离子模式的过滤条件为:频率>50%,响应值过滤>10000,p<0.05,倍数变化阈值(FC)≥1.5;负离子模式的筛选条件为:频率>50%,响应值过滤>7000,p<0.05,FC≥1.5。随后,使用包含超过80,000种代谢物信息和约11,000种二级质谱数据的Metlin数据库,我们鉴定并分析了差异化合物,以确定代谢物的身份和结构特征,为生物学解释奠定了基础。
2.5.3 靶向代谢组学
最初识别目标化合物包括回顾与高强度运动相关的代谢物的文献,然后进行一级质谱分析,以及由于一级响应不佳或二级质谱质量不足而无法纳入个人化合物数据库和库(PCDL)的化合物。最终成功纳入了15种化合物(如表1所示)。准备好的样本在机器上进行测试,测试数据与已建立的PCDL进行定性匹配,以分析样本中是否存在目标化合物以及检测到的化合物是否有响应值差异。这15种代谢物随后被用作浓度为1 µg/mL的混合标准溶液进行定量控制分析。这15种氨基酸代谢物最终使用MassHunter定量软件进行靶向。这些代谢物质谱碎片离子信息如表1所示。
3 结果
所有10名参与者完成了所有研究测试,他们的数据被纳入分析。所有受试者都无法区分氢气和安慰剂气体之间的差异。他们也没有报告吸入氢气有任何副作用。
3.1 吸入富含氢气的气体(HRG)改善短跑间隔测试
主要配对t检验模型显示,HG中的S4的MP显著高于PG(t=2.585,p=0.029),且HG中的ΔS4-S1的MP显著大于PG(t=3.579,p=0.006)。其次,HG中S4的FI和TTP显著低于PG(FI: t=-4.567,p=0.001;TTP: t=-3.643,p=0.005),但S4的PP在HG和PG中没有显著差异(t=2.114,p=0.064)。ΔS4-S1的PP在HG中显著大于PG(t=5.046,p=0.001),ΔS4-S1的TTP在HG中显著小于PG(t=-3.939,p=0.003),但ΔS4-S1的FI在HG和PG中没有显著差异(t=0.234,p=0.820)。这些结果表明,运动前吸入HRG有助于在高强度运动后的冲刺骑行中维持无氧性能并减轻疲劳。
3.2 非靶向代谢组学测试结果
MPP软件从正离子模式获取的数据中总共识别出5,717种代谢物。通过筛选出现频率超过50%的代谢物,得到1,639种代谢物。进一步基于响应值优化后,得到474种代谢物。在排除源于溶剂的化合物后,最终剩下300种代谢物。在HG和PG中,每种代谢物在吸入前后均无显著差异(p>0.05),但有18种代谢物在SIT前后显示出显著差异(p<0.05)。与METLIN数据库比较后,鉴定出4种具有显著差异的代谢物。进一步分析这4种代谢物在HG和PG中的ΔSIT显示,乙酰肉碱、PS(O-18:0/0:0)和丙酰基-L-肉碱在HG中比PG更显著上调,而4-甲基苯基辛酸酯在HG中比PG更显著下调(p<0.05)。
MPP软件从负离子模式获取的数据中总共识别出11,683种代谢物。通过筛选出现频率超过50%的代谢物,得到3,903种代谢物。进一步基于响应值优化后,得到373种代谢物,其中没有来自溶剂的化合物。在HG和PG中,吸入前后每种代谢物均无显著差异(p>0.05),但有14种代谢物在SIT前后显示出显著差异(p<0.05)。与METLIN数据库比较后,鉴定出4种具有显著差异的代谢物。Coriose、Methoxyacetic acid在SIT后显著上调(p<0.05),但其ΔSIT在HG和PG之间无显著差异(p>0.05)。(±)-6-methyl-caprylic acid和2-(Ethylsulfinylmethyl) phenyl methylcarbamate在SIT后显著下调(p<0.05),且2-(Ethylsulfinylmethyl) phenyl methylcarbamate在HG中的下调显著大于PG(p<0.05)。然而,(±)-6-methyl-caprylic的ΔSIT在HG和PG之间无显著差异(p>0.05)。
从样本中迭代收集的二次数据使用MassHunter定性软件进行二次质谱确认,并且在上述非靶向筛选的代谢物中,Acetylcarnitine和Propionyl-L-carnitine除了其分子离子峰外,还具有至少一个碎片离子,从而便于其定性鉴定。
3.3 靶向代谢组学测试结果
所有样本中的15种氨基酸代谢物使用MassHunter定量软件进行靶向。在HG和PG中,吸入前后每种代谢物均无显著差异(p>0.05),但有3种代谢物(Hypoxanthine、Xanthine、D-Pantothenic acid)在SIT前后显示出显著差异(p<0.05)。收集的靶向分析数据通过MPP软件进行分析。Hypoxanthine在正离子模式下SIT后上调,且HG的上调显著大于PG(p<0.05)。Hypoxanthine和Xanthine在负离子模式下SIT后上调,且HG的上调显著大于PG(p<0.05)。
3.4 与差异代谢物相关的多条代谢途径
对HG中的差异代谢物进行代谢途径分析,KEGG数据库匹配分析显示了前20个显著富集的相关途径。其中,某些分子特征与氢气吸收代谢有关,如甘油磷脂代谢、泛酸和辅酶A(CoA)生物合成以及胰岛素抵抗(图2)。
4 讨论
据我们所知,这是首次研究通过代谢组学分析探索运动前吸入富含氢气的气体(HRG)改善短跑间隔训练表现的潜在机制。发现了四类化合物是HRG益处的潜在关键机制代谢物,包括来自非靶向分析的Acetylcarnitine和Propionyl-L-carnitine,以及靶向分析中的Hypoxanthine和Xanthine。
Hypoxanthine是一种天然存在的嘌呤衍生物,作为tRNA反密码子中的肌苷,是核酸成分[19]。血浆次黄嘌呤浓度随运动强度而变化。高强度运动后,次黄嘌呤水平呈指数级上升[20, 21]。次黄嘌呤水平与细胞ATP消耗直接相关[22]。高强度运动引起ATP水解增加,ADP和缺氧增多,从而增强ADP的代谢产生次黄嘌呤。次黄嘌呤随后被黄嘌呤氧化酶转化为黄嘌呤[23]。在我们的研究中,吸入HRG的SIT表现更好,同时伴有更高浓度的次黄嘌呤和黄嘌呤,这表明吸入氢气后,冲刺过程中不断有更多ATP被分解以提供能量。Acetylcarnitine和Propionyl-L-carnitine是由短链脂肪酸(SCFA)与肉碱结合形成的酯,其主要生物学功能是将脂肪酸从胞浆运输到线粒体基质中进行氧化[24]。脂肪动员增加与SCFA水平升高有关。在我们的研究结果中,吸入HRG后Acetylcarnitine和Propionyl-L-carnitine的浓度更高,这表明氢气可能增强了线粒体脂质代谢并加速了脂肪酸的β-氧化代谢[25]。以前的研究也表明氢气增加了脂肪氧化[26, 27]。
在SIT期间,参与者在高强度冲刺运动后体内产生大量ROS[28],这会导致线粒体氧化损伤并降低后续的有氧利用[4]。这导致ATP补充不足,降低了后续的无氧冲刺表现。吸入HRG可以减轻这种效应。通常,电子传递链会创建质子梯度,从而促进ATP的产生[29]。但ATP也可以通过建立氢梯度独立产生[30]。Dohi发现氢分子可以创建梯度,独立于电子传递链促进线粒体ATP的产生[31]。然而,我们的研究发现,氢可能通过增加脂肪氧化和形成更多ATP来增强SIT表现。在SIT中,30秒的冲刺阶段依赖于ATP-CP和无氧糖酵解,而4.5分钟的间歇期涉及有氧氧化以补充磷酸肌酸储备,为下一次无氧冲刺提供所需能量。氢清除了高强度运动期间产生的大量ROS,减少了它们对线粒体的氧化损伤,从而提高了有氧途径的利用[4, 32]。减少氧化损伤使得线粒体更有效地将底物转化为ATP,提供更多的细胞能量。线粒体效率的增加不会直接影响ATP-CP系统的功能,但有助于运动间歇期间更快的恢复,从而改善ATP的补充,为下一次运动提供能量。这导致S4表现提高和疲劳减少(ΔS4-S1)。
泛酸是CoA的前体,对于依赖CoA的反应至关重要,尤其是在脂质代谢和脂肪酸β-氧化中[33]。途径富集分析显示,泛酸和CoA的生物合成途径得到加强,这表明氢可能增强CoA的合成,从而促进酰基肉碱的产生并增强脂质氧化。在甘油磷脂代谢过程中,甘油磷脂分解成游离脂肪酸有助于脂质氧化过程。观察到甘油磷脂代谢以及脂肪消化和吸收途径的富集,强调了氢在促进脂质代谢中的作用。此外,酶的激素调节在脂肪动员中的作用,特别是胰岛素对激素敏感脂肪酶活性的影响,展示了脂肪动员与胰岛素敏感性之间的复杂相互作用,已知这会随着体育锻炼而改善[34]。然而,氢抑制胰岛素水平升高的能力可能激活激素敏感脂肪酶,从而增强脂解作用[35, 36]。这与胰岛素抵抗代谢途径中的富集相一致,提供了氢如何影响代谢过程的潜在机制的见解。
5 结论
总之,吸入HRG后进行SIT可以提高最终的无氧冲刺表现并减轻疲劳。代谢组学分析表明,HRG的作用机制可能是消除初始冲刺期间产生的大量ROS并减少对线粒体的氧化损伤,从而提高有氧利用的效率,并可能通过增加辅酶A合成、促进甘油磷脂代谢和抑制胰岛素水平来促进脂肪动员,帮助加速间隔阶段的脂肪氧化,恢复更多ATP的补充,为后续冲刺提供更好的无氧输出。
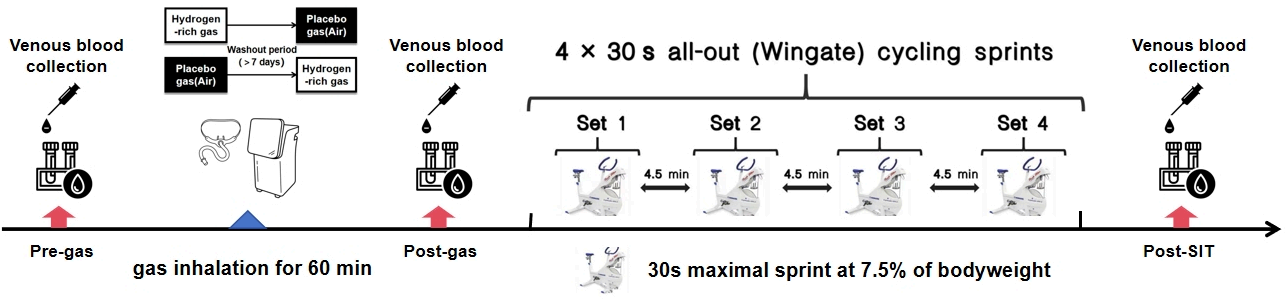
图 1实验试验顺序和程序。气前:吸入前、气后:吸入后立即;安装后:冲刺间隔试验后立即。
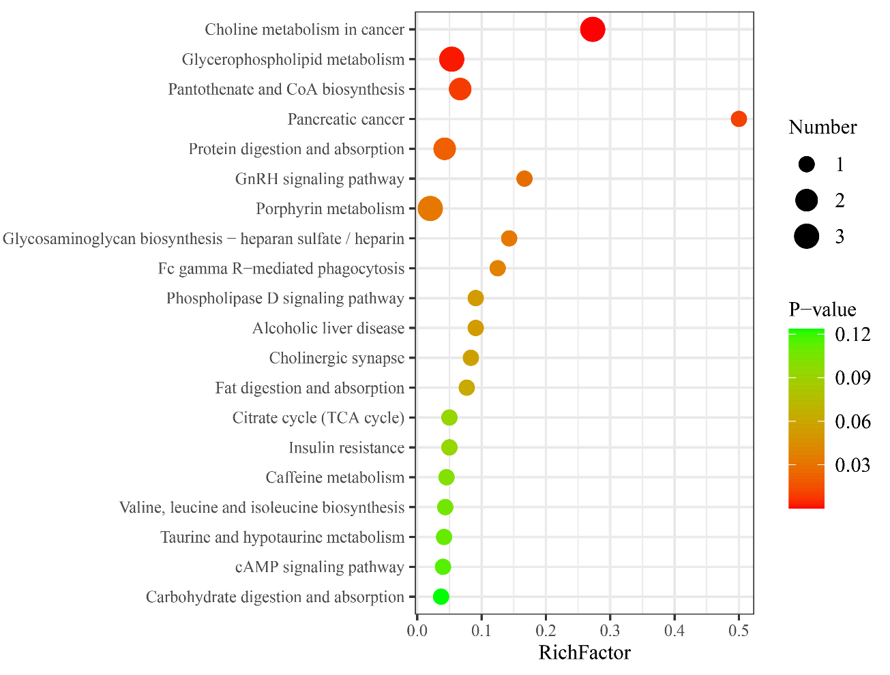
图2 氢组中不同代谢物的前20个差异代谢途径。
https://blog.sciencenet.cn/blog-41174-1437393.html
上一篇:哺乳动物产热进化
下一篇:氢气减少七氟烷诱导新生儿皮层神经元凋亡
