博文
改写教科书的新现象:心率相关脑电震荡  精选
精选
|
与心跳相关的脑点图频率。做研究要细心,有时候不能过于聪明过于主观,要较真要耿直。例如本研究所发现的和心跳相关的脑电频率,如果是我做实验,肯定会视而不见。因为会把这现象当成是心跳或心电导致的干扰。不会认为这是大脑自身迎合心跳产生的共振。
1942 年,医学诺贝尔奖(32年)获得者英国电生理学家 E. D. Adrian 发表了刺猬嗅球 (OB) 的电信号记录,表明有三种基本类别电振荡:呼吸-鼻-气流相关振荡 (RRO)、感觉-气味诱导的振荡和局部神经网络固有的振荡 (1)。这些振荡很重要,因为通过将神经元静息膜电位移向和远离尖峰阈值,它们可以同步本地和远程神经网络中的尖峰活动。所有三类振荡都依赖于突触传递。在其他大脑区域也有记录,尤其是在人类大脑皮层和海马体中。并被认为是大脑正常处理信息的基础 (2)。在本期《科学》第494页,Jammal Salameh等(3) 报告了第四类振荡——由动脉压脉动诱发并由中枢压力感受器转导的心跳相关振荡 (HRO)。

Jammal Salameh等重点是了解在大鼠 OB 僧帽细胞 (MC) 层中记录的 ~4 Hz 的突出局部场电位 (LFP) 振荡的机制。LFP是在细胞周围的局部细胞外空间中产生的电信号,包括兴奋性和抑制性突触的综合活性以及任何刺突活性。与阿德里安的研究一样,大型啮齿动物OB提供了一个高度可及的大脑区域进行记录。但是,在 Jammal Salameh 等人的研究中,鼻脑制备 (NBP) 大鼠模型 (4) 的性质意味着呼吸和感觉诱发机制可以被排除在外,因为缺乏肺、鼻气流和上行呼吸通路。此外,该模型中没有心脏似乎排除了心跳诱导的压力搏动的作用(大鼠~4 Hz)。然而,有点巧合的是,用于向NBP灌注含氧溶液的蠕动泵由八个单独的滚轮组成,因此对于30 rpm(0.5 Hz)的典型泵旋转速率,会产生4 Hz的压力脉动。通过同时记录LFP振荡和测得的压力脉动,作者确定了慢速LFP振荡的基频与2~4 Hz脉动范围内的压力脉动频率之间的完美匹配。此外,多条证据表明,LFP既不是搏动伪影,也不是像其他三种振荡那样由轴突突触传递介导的。相反,他们研究了中枢压力感受器是否在起作用,其中压力脉动由压力激活通道转导,例如以 MC 表达的压电式机械敏感离子通道组分 2 (PIEZO2) (5,6)。值得注意的是,Jammal Salameh 等通过考虑 PIEZO2 通道的特定压力灵敏度和门控特性,能够精确模拟 LFP 波形和观察到的自发 MC 尖峰峰值增加。
为什么 Adrian 或在过去 80 年进行的许多脑振荡研究中都没有检测到 HRA?Jammal Salameh等发现心跳调节的尖峰仅发生在一小部分(~15%)的OB神经元中,仅涉及放电速率的适度增加(~10%),并且在清醒的头部固定小鼠的记录中,心跳后20毫秒达到峰值。相比之下,鼻呼吸诱导的 RRO 夹带了更高比例的 OB 神经元 (~94%),涉及更大的放电增加 (~30%),并在吸气后 150 毫秒达到峰值。如果没有特定的焦点,这些与心跳相关的微妙影响可能会被遗漏或误解为搏动伪影。值得注意的是,作者在小鼠海马体和额叶新皮层中记录了同样微妙的心跳相关反应,它们也表达 PIEZO2 通道蛋白 (6)。
HRO在大脑功能中可能扮演什么角色?HRO可能被大脑过滤掉为“生理噪音”,因为心跳始终存在 - 从最早的胚胎大脑阶段到脑死亡都感觉到 - 并且保留在所有大脑状态中,从高度唤醒到最深的睡眠状态。有趣的是,人类指尖中许多 (≤60%) 特化触觉受体和人体肌肉中拉伸感受器的尖峰要么是锁相的(即,每次心跳后都会出现一个尖峰),要么由周围组织中产生的心跳诱导的动脉压搏动调节 (7)。这与外周神经元中 PIEZO2 通道密度较高与中枢神经元密度一致 (6)。尽管大脑可能会忽略传入的尖峰作为噪声,但在狭窄的时间窗口(即 <100 毫秒)内到达的一连串传入尖峰可能会通过类似于随机共振的机制降低中央突触的触觉和拉伸阈值 (7)。HRO 可以通过加强到达 OB 和海马体的突触输入来执行类似的功能。
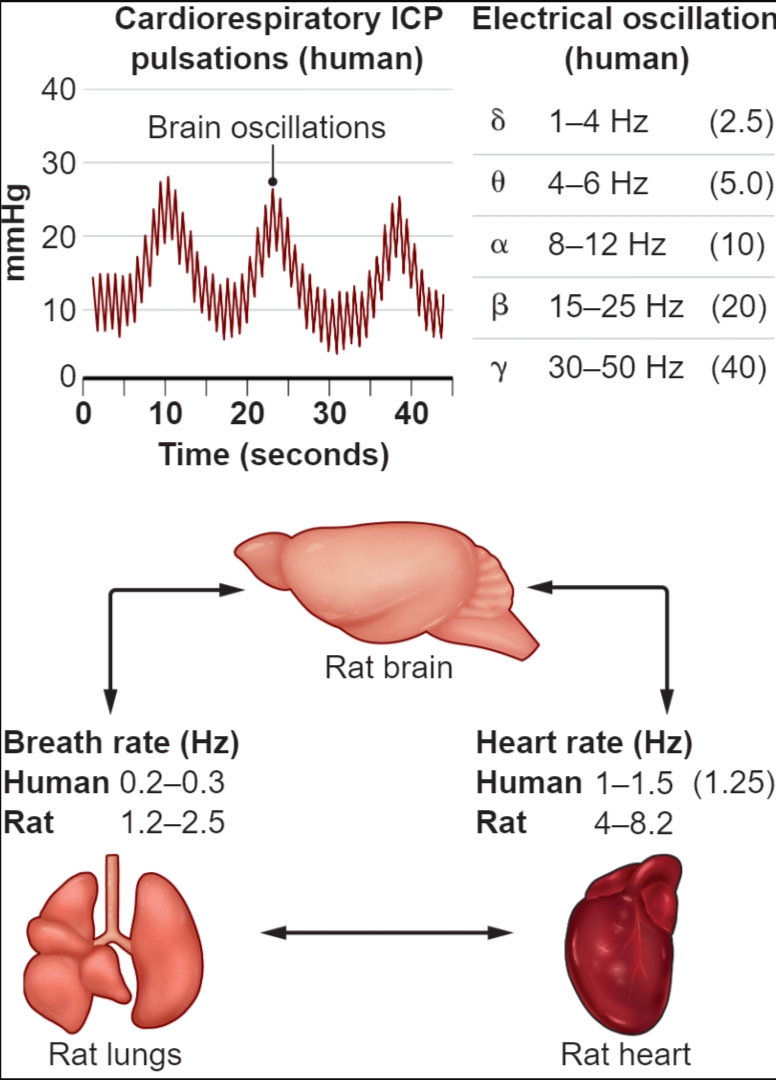
此外,HRO可能在大脑中发挥更重要的作用。与RRO(8,9)一样,HRO可能通过在睡眠期间重新点燃它们来稳定与新记忆相关的神经组件,从而抵消任何损失。 也有人提出,大脑和身体的振荡是相互联系的(10,11),甚至谐波相关,形成中心频率的二元层次结构,其中基频为1.25 Hz,对应于健康人类的心率为75 bpm(见图)。 在这些提案中,HRO现在可以提供心跳脉动与脑电振荡的机械电耦合机制。
心跳不表现为节拍器,而是高度可变的。除了对精力充沛和情绪需求的反应外,心率还通过正常呼吸来调节,这种现象称为呼吸窦性心律失常(即,吸气时心率增加,呼气时心率降低)(12)。这在共振呼吸中最为明显,当呼吸频率降低到 0.1 Hz (6 bpm) 时,压力感受器反射(涉及外周压力感受器和脑干调节核之间的反馈回路)、血压变化和呼吸同步(即共振)。共振呼吸,特别是涉及鼻吸气,用于冥想练习,它被认为可以减少焦虑和缓解惊恐发作以及其他精神疾病(12)。这些效应依赖于由 PIEZO 通道转导的心脏和动脉传入棘突活动,并且主要通过迷走神经传递到特定的大脑区域 (13)。预计与共振呼吸相关的心率的巨大正弦波动将增加颅内压搏动和 HRO,从而在脑功能中发挥额外的有益作用。

介绍
神经电振荡被认为是大脑处理信息的基础。各种振荡模式反映了局部或全脑网络中的处理,并且自发发生或与感觉和认知处理有关。越来越多的证据表明,这种神经振荡也可以通过身体节律的内感受来调节,例如呼吸或心跳。内感受是对身体内部信号的感知,而不是对外部世界的感官感知,从而告知大脑有机体的状态。
理由
为了研究受限网络内局部振荡的机制,我们开发了一种半完整的大鼠嗅球制剂 - 大脑中嗅觉处理的第一个站,以其强大的振荡活动而闻名。在这种类型的简化制剂中,没有心脏、肺或其他大脑区域的输入,并且球的脉管系统通过蠕动泵灌注人工血液。该泵在脑血管系统内产生压力脉动,在我们的设置中,恰好符合体内颅内压心跳诱导脉动的生理范围。值得注意的是,这些泵诱导的机械脉动之后是源自嗅球主要神经元僧帽细胞的局部电场振荡。根据最近关于机械敏感离子通道在大脑中主要神经元子集中表达的证据,我们假设这些神经元可能能够直接感知与心跳相关的血管血压脉动。
结果
泵诱导的压力脉动为嗅球僧帽细胞内的机械感觉转导通路提供了足够的刺激。这种转导是由快速兴奋性机械敏感离子通道介导的,这些离子通道存在于僧帽细胞的一个子集中。在许多其他身体组织中,Piezo2 已被证明有助于检测相似频率范围内的振动。它的门控特性很可能是将压力刺激的正弦波形转换为我们制备中观察到的更复杂的局部场振荡波形的基础。虽然这种快速转导途径不涉及突触传递,但血管压力脉动节律夹带了僧帽细胞的自发尖峰活动。因此,机械感觉转导对尖峰时间产生了直接的调节影响。这条通路可以让大脑在体内感知心跳吗?在清醒的小鼠中,我们发现神经元的尖峰活动实际上是由心跳调节的,~15%的嗅球神经元被这种节律夹带,主要是在20毫秒内。这种效应比已知的神经元活动与呼吸节律的耦合要弱得多,这解释了为什么直到现在还没有观察到它。我们在海马体和前额叶皮层中也观察到类似的心跳诱导的神经元活动调节。
结论
内感受在脑功能中的作用是当前神经科学的主要挑战之一。在人类中,最近的实验证据支持通过心动周期调节自主和有意识的感知和认知。尽管这种调节部分是由源自主动脉压力感受器的经典上行多突触通路介导的,但本研究结果表明,心跳诱导的脑血管搏动可以通过激活机械敏感通道直接影响中枢神经元活动。尽管目前这种直接通路的功能尚待推测,但我们提出,一个由“心跳哨兵神经元”组成的全脑网络介导认知、情绪和自主神经状态的内感受调节。例如,某些唤醒状态的发生可能与该网络的激活有关。我们的发现为内感受体脑轴增加了一条快速传输线,中枢神经元可以感觉到大脑内的脉搏。
https://blog.sciencenet.cn/blog-41174-1420266.html
上一篇:氧化应激在神经退行性疾病中的作用:活性氧和抗氧化剂的预防研究进展
下一篇:AI识别婴儿认识世界模式给我们的启发
