博文
[转载]RNA m6A修饰问题汇总(第二期)
||
上期推出了第一期m6A修饰的相关问题汇总,解答了有关m6A修饰的一些入门级问题,本期继续推出相关问题汇总,深入探讨m6A机制研究的难点与挑战。m6A的研究故事怎么讲?之前的研究思路在自己所关注的疾病过程中是否适用?一直在讨论稳定性、翻译效率、降解等关键词,但m6A这些作用是普适性的功能还是具有时空特异性?随着研究成果的不断积累和相关讨论的不断深入,是时候系统性的总结归纳现有研究对我们的启发与思考了。在第二期问题汇总中,一起来看看有哪些有关机制讨论的相关问题被研究者关注,文中相关观点和文献支持结合2021年发表的文献综述进行讨论,感兴趣的老师可以后台回复“文献”进行索取。
参考文章封面: 文章原文:m6A RNA methylation: from mechanisms to therapeutic potential
文章原文:m6A RNA methylation: from mechanisms to therapeutic potential
有关m6A修饰的序列偏好性、区域偏好性、位置选择、METTL3/METTL14、m6A发生的内外调控系统、时空特异性问题请见上一期文章。本期将重点围绕Reader蛋白和Eraser蛋白讨论研究者经常关心的其他问题:
一.Reader蛋白可以识别并结合m6A位点,这种结合有什么特征?对结合的RNA稳定性有什么影响?
在综述文章中,作者描述依赖于m6A降解和翻译的过程是“context-dependent”,这里的context指的是被m6A修饰mRNA分子具体的相关特征,也就是结合带来的影响要看修饰位点的位置、区域、Reader蛋白的类型,以及最重要的被修饰RNA分子的特征。
目前多个相关研究中达成共识的是m6A可以通过YTHDF2分子结合降低mRNA稳定性,与之相反的是IGF2BP1、IGF2BP2、IGF2BP3识别m6A位点增强mRNA稳定性(图1)。RIP-seq可以看到YTHDF2蛋白的结合位点信息与m6A的位点分布近似,但是IGF2BP1-3分子的结合位点的分布则与这两者不同。不同之处在于IGFBP1-3结合位点没有富集于终止密码子,而是均衡分布于3’UTR区,IGF2BP1-3和YTHDF2的结合位点重合度也比较低。尽管IGF2BP1-3与m6A RNA结合的模式还没有完全解析,但是可以推测他们是通过KH功能域同m6A RNA结合。因此,被m6A修饰的RNA片段是和IGF2BP1-3结合变得更加稳定,还是和YTHDF2结合变得不稳定,主要还是看m6A RNA片段的周围序列特征和相应结合的蛋白活性。
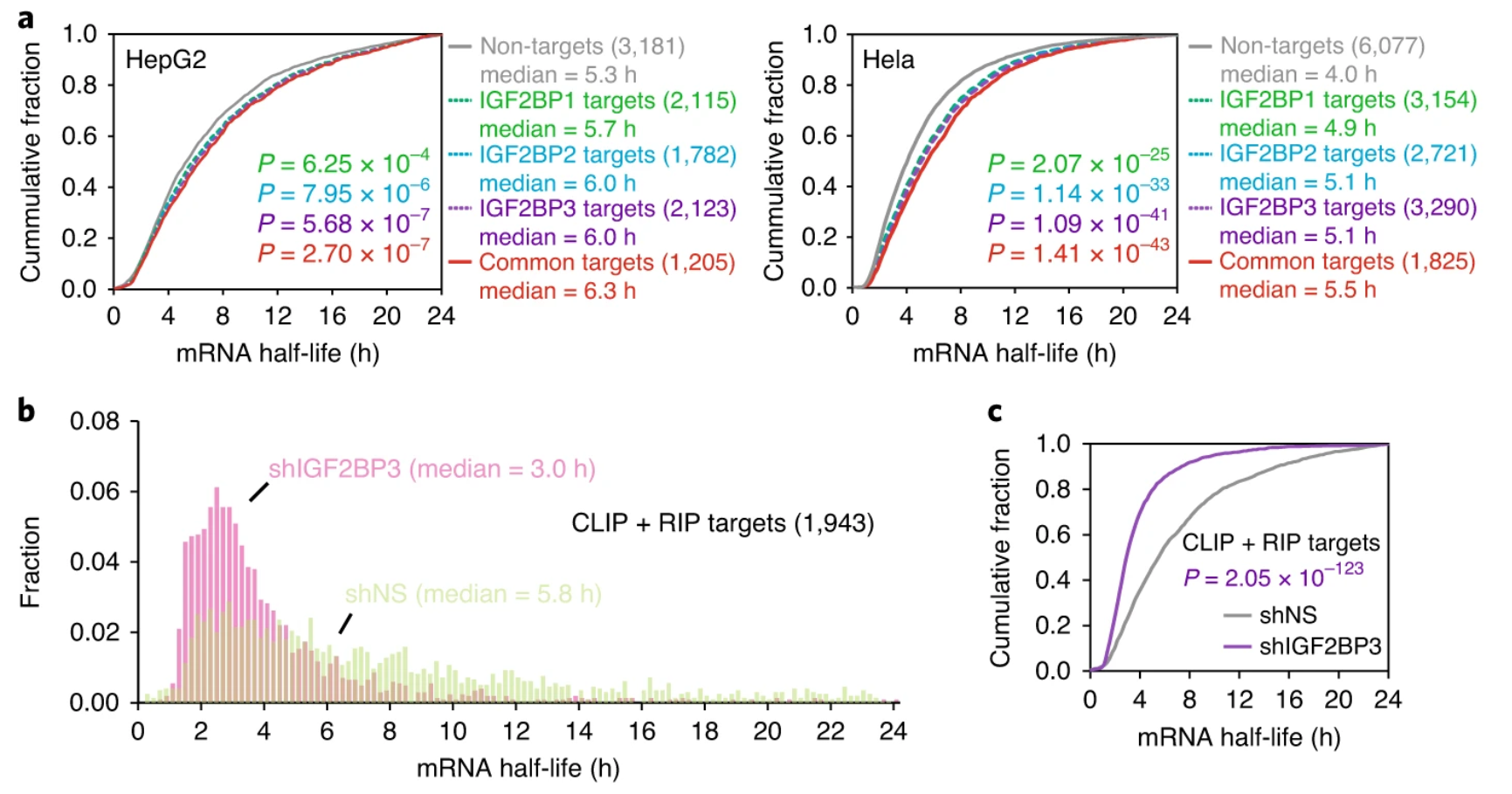
图1:敲低IGF2BPs降低mRNA稳定性
(Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation)
二.Reader蛋白对结合的RNA翻译过程有什么影响?
m6A影响mRNA翻译也是有类似的特征,相关研究显示在HeLa细胞和子宫内膜癌细胞中,YTHDF1选择性的结合到m6A修饰转录本并增强翻译效率,但是这种YTHDF1对翻译的上调作用是否明显主要还是看具体的细胞环境,在HEK293T细胞系和乳腺癌细胞系中人工构建的m6A报告基因的上调效果就并不明显。最近围绕学习和记忆缺陷的小鼠模型的研究中发现,这种情况下YTHDF1介导的翻译上调作用有可能由外界的刺激因素而诱导引发,海马特异的敲除YTHDF1可以引起小鼠的空间记忆缺陷和情景恐惧记忆(contextual fear memory)。在培养的海马神经元的稳态环境中,YTHDF1对翻译的影响相对温和,但是在使用氯化钾去极化处理后,这种影响会变得更加明显,同时在围绕背根神经节(dorsal root ganglion)的研究发现,虽然正常情况下YTHDF1对背根神经节的影响比较微小,但是在损伤诱导轴突再生过程中,YTHDF1促进蛋白合成是必不可少的环节。

图2: YTHDF1增强m6A修饰基因的翻译对神经刺激产生应答
(m6A facilitates hippocampus-dependent learning and memory through YTHDF1)
m6A修饰的具体位置也会改变YTHDF1对于翻译的影响,5’UTR m6A能够增强非帽结构依赖的翻译起始,在CDS区域的m6A位点可以调控翻译延伸,终止密码子附近和3’UTR的m6A位点可以被YTHDF1和YTHDF3识别,增强翻译起始,总而言之,不同的位置修饰位点,对RNA带来的影响也会有所区别。
三.除了mRNA本身以外,m6A对非编码的影响如何?又是怎样发挥功能的?
除了mRNA发生m6A修饰以外,非编码RNA同样广泛存在m6A修饰是被广泛认同的事实,只是之前大部分研究聚焦在mRNA m6A。在另外一类非编码RNA分子中(综述文章称为非编码染色体相关调控RNA,non-coding chromosome-associated regulatory RNA,carRNA)m6A修饰同样重要,m6A本身可以造成这些carRNA的降解从而影响这些非编码RNA分子的临近基因。举个例子,在自我更新和分化的ESCs中,早期的研究关注的是细胞质m6A阅读蛋白对整个细胞表型的影响,细胞质中,编码多能性维持因子蛋白的转录本容易发生m6A修饰进而被YTHDF2结合促进其降解,细胞也在特定时间点从胚胎干细胞开始分化,Mettl3敲除导致早期胚胎致死,Ythdf2敲除鼠可以维持到胚胎发育晚期,所起作用类似于在核中敲除Ythdc1(核m6A Reader蛋白),说明m6A可能在细胞核中也发挥重要作用。
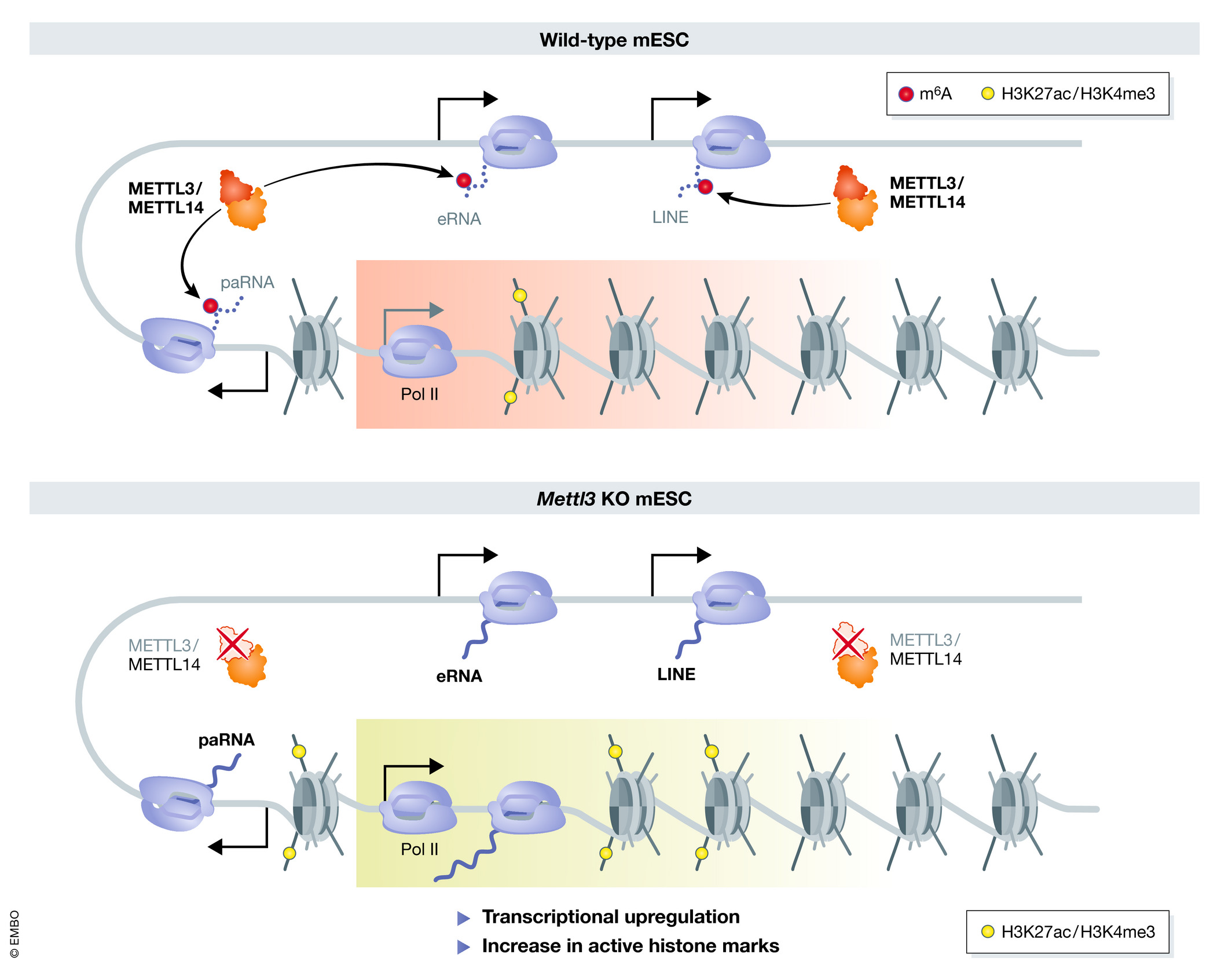
图3:carRNA的m6A位点调控染色质开放状态以及转录活性
(m6A RNA methylation: from mechanisms to therapeutic potential)
高通量测序的结果表明,METTL3不仅可以催化带有polyA结果的mRNA和lincRNA发生m6A修饰,同时,对非polyA结构的线性转录本、启动子相关转录本(promoter-associated RNA)、增强子RNA(enhancer RNA)、转座元件来源RNA(repeats RNA)也发生m6A修饰,这些非编码RNA分子被统称为carRNA,其中15%-30%均含有m6A修饰,METTL3敲除可以广泛的上调这些RNA丰度。METTL3的敲除可以增强染色质的开放性,广泛的上调转录活性(图3),由于paRNA和eRNA是部份基因转录的上游调控因子,因此这些m6A修饰的paRNA或者eRNA在METTL3敲除后均能上调靶分子,m6A水平下降可以延长carRNAs的半衰期,最终上调下游基因的转录,提高局部H3K4me3和H3K27ac的水平,表明m6A修饰carRNA可以间接实现调控转录的过程。
四.去甲基化的调控过程是怎样的?
m6A过程动态可逆的特点是非常重要的,ALKBH5和FTO是两个目前认为最重要的去甲基化酶,在癌症进展、免疫和精子形成过程中都发挥重要作用。FTO的特点是功能非常强大,强大到在很多病理、生理过程中都能发挥作用,对于分子来讲,从pre-mRNA、可变剪切、多聚APA到翻译,FTO每时每刻都在发挥作用;FTO的底物非常广泛,以至于m6A、m6Am、tRNA m1A都能被FTO调控发生去甲基化,有关于FTO活性和底物的争论还有很多。研究表明,对于FTO的底物选择更多的依赖于FTO的亚细胞定位,对m6A去甲基化作用更多的是在细胞核中被报道,对m6Am的去甲基化作用更多的研究是围绕细胞质进行。由于FTO的作用十分强大,且底物分布十分广泛,在AML细胞甚至FTO介导了40% mRNA m6A的去甲基化过程,有关FTO作用主导的调控细节仍有不少争论点,下面的例子说明,想要研究清楚FTO到底通过哪条途径影响表型并不是一件容易的事。
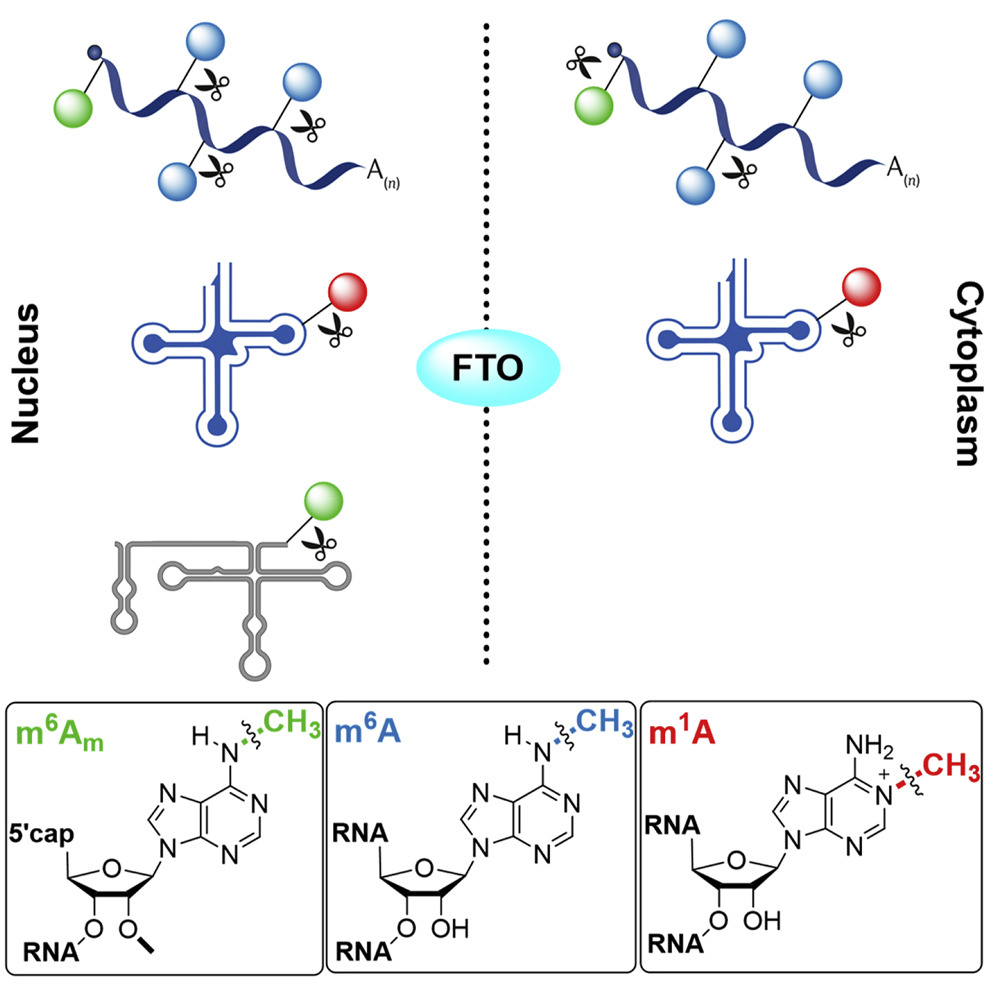
图4:FTO可以作用m6A,m6Am,m1A三种甲基化修饰类型
(Differential m6A, m6Am, and m1A Demethylation Mediated by FTO in the Cell Nucleus and Cytoplasm)
由于FTO的影响涉及多种底物,因此对特定过程中FTO对其生物功能影响是比较复杂的,例如FTO的敲除大部分情况会引起小鼠模型的胚胎致死,即便是幸存的模型鼠发育明显缺陷,体重较对照更轻,那这种表型变化到底是由于FTO使哪种底物的m6A修饰发生了去甲基化作用而产生?
分别做实验来看,首先敲除介导mRNA帽子结构和U2 snRNA m6Am修饰的Pcif1或者Mettl4酶,表型不明显,也不影响小鼠的生存情况,但是敲除Mettl13、Mettl14(催化mRNA m6A)就会明显引起胚胎致死,需要注意的是,相比于Fto敲除,Mettl14\Mettl3 敲除鼠表现出更严重的发育问题,而Fto敲除鼠的发育问题相比Pcif1或者Mettl4敲除鼠更严重,因此cap mRNA、U2 snRNA m6Am甲基化对于胚胎发育过程并没有那么广泛。这些比较实验说明,mRNA 和U2 snRNA m6Am修饰的去甲基化并不是造成Fto 敲除导致发育缺陷表型的主要原因。
五.Open Question:当前m6A研究的难点和挑战在哪里?
m6A对mRNA的功能影响是多变且多面的,举两个例子来看:在Hela细胞中,YTHDF蛋白均促进RNA降解,结合位点、蛋白相互结合以及亚细胞定位pattern均呈现相似的结果,研究结果甚至表明YTDHF1在Hela细胞中并不能促进翻译;第二项研究中,Lasman等人发现在小鼠的早期发育过程中,mESCs中三个阅读蛋白之间存在代偿关系,相比于同时敲除三个蛋白,单独敲除某一个Ythdf蛋白不会造成mESC分化和mRNA的降解。由此看出,针对包括Writer、Reader、Eraser在内的各项研究都具有自己的特点和特性,并不能以一盖全,整个调控过程极其复杂,甲基化去甲基化的动态过程受多方面调控,因此针对特定的生物学过程的相关表型,结合之前的类似研究,进行综合性的评价和讨论,才能完善m6A的调控机制和作用关系。
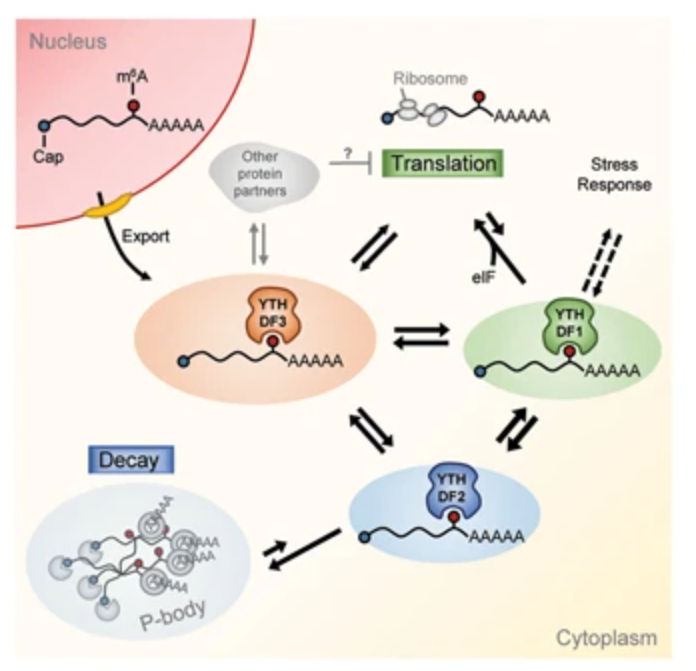
图5:YTHDF蛋白功能网络
(YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA)
第二期的总结当中,讨论了一些有关m6A RNA的重要机制问题,整体来看RNA m6A的功能多样,分布广泛,在不同的生物学过程中既有共性也有特性,下一期的RNA m6A问题总结中,我们将整理一些有关m6A分析的常用生信工具和技术手段,方便各位研究者查询有效信息,辅助m6A相关研究,敬请期待!
https://blog.sciencenet.cn/blog-707141-1337861.html
上一篇:[转载]RNA m6A修饰问题汇总(第一期)
下一篇:[转载]RNA m6A修饰工具大全