ВЉЮФ
ШЫРрЮЊЪВУДУЛгаЮВАЭЃП
|
ЮВАЭЪЧЖЏЮяЭѕЙњЕФЙВЭЌЬиеїЃЌЫљгаВИШщЖЏЮядкХпЬЅЗЂг§Й§ГЬжаЕФФГИіЪБКђЖМгаЮВАЭ 1 .дкШЫРржаЃЌЮВАЭдкХпЬЅЦкНсЪјЪБЯћЪЇЁЊЁЊДѓдМдкзгЙЌФкАЫжм 2 ЁЊ ОЁЙмФкВПСуМўШдвдЮВЙЧЕФаЮЪНДцдкЁЃЮВАЭЕФЯћЪЇБЛШЯЮЊЪЧРрШЫдГЕФвЛИіЯдзХЬиеїЃЌПЩФмгАЯьСЫЮвУЧздМКЕФжБСЂаазпЗНЪНЁЃдк ЁЖздШЛЁЗдгжОЩЯЕФвЛЦЊТлЮФжаЃЌXia et al . 3 БЈЕРГЦЃЌдкНјЛЏЙ§ГЬжаЃЌвЛжждкЛљвђзщжмЮЇвЦЖЏЕФвЦЖЏЛљвђађСаЕФВхШыЃЌГЦЮЊзЊзљдЊМўЃЌПЩФмгыЮВАЭЕФЖЊЪЇгаЙиЁЃ
A mobile DNA sequence could explain tail loss in humans and apes (yyttgd.top)
ДѓЖрЪ§КязгЖМгаЮВАЭЃЌЮВАЭдк6500ЖрЭђФъЧАОЭДцдкгкСщГЄРрЖЏЮяЦзЯЕЕФЦ№дДжа 4 .ЪТЪЕЩЯЃЌУЛгаЮВАЭЪЧЧјЗждГКяКЭКязгЕФвЛжжЗНЪНЁЃетжжЬиеїЛђБэаЭдкЫљгаРрШЫдГжаЖМЪЧЙВгаЕФЃЌетБэУїЮВАЭЕФЩЅЪЇгыдГРрЕФсШЦ№ЯрЮЧКЯЃЌЛђепЗЂЩњдкдГРрдкДѓдМ2500ЭђФъЧАгыКязгЕФзюКѓвЛИіЙВЭЌзцЯШЗжЕРбяя№КѓВЛОУЁЃ
гаСЫетаЉжЊЪЖЃЌXiaМАЦфЭЌЪТБШНЯСЫМИжжСщГЄРрЖЏЮяЛљвђзщжаЮВАЭЖЊЪЇЕФжївЊКђбЁЛљвђЃЌзюГѕЙизЂЕФЪЧЭтЯдзгЃЈБрТыЕААзжЪЕФDNAЧјгђЃЉЁЃЕБетжжЗНЗЈУЛгаНсЙћЪБЃЌзїепНЋЫћУЧЕФбаОПРЉеЙЕНЛљвђЩЯгЮЛђЯТгЮЛђЛљвђБОЩэЕФЗЧЕААзжЪБрТыЧјгђЁЃКѓвЛИіЧјгђБЛГЦЮЊФкКЌзгЃЌЫќУЧЭЈГЃжаЖЯЕААзжЪБрТыађСаЁЃXiaМАЦфЭЌЪТЗЂЯжЃЌвЛжжСщГЄРрЖЏЮяЬивьадзЊзљдЊМўГЦЮЊAluдЊМў 5 ШыTBXTЛљвђЕФФкКЌзгжаЃЌЕЋНіЯогкРрШЫдГЃЌЖјВЛДцдкгкЦфЫћСщГЄРрЖЏЮяЦзЯЕжаЁЃ TBXT вВБЛГЦЮЊ Brachyury ЃЈвтЮЊЁАЖЬЮВЁБЃЉЃЌвђЮЊИУЛљвђЕФЭЛБфгыЖрИіЮяжжЕФЖЬЮВгаЙиЃЌАќРЈАЂЖћМАРћбЧаЁЪѓЃЈ Mus spretus ) 6 КЭТэЖїЕКУЈЃЈ Felis catusЃЉ) 7 .
ЕЋЪЧЃЌНЋвЛИіЖЬЕФЁЂДѓдМ300ИіМюЛљЖдЕФAluдЊМўВхШыЕНЛљвђЕФЗЧБрТыађСажаЃЌШчКЮгажњгкЮоЮВБэаЭФиЃПЮЊСЫЛиД№етИіЮЪЬтЃЌXiaМАЦфЭЌЪТНјвЛВНзаЯИбаОПСЫећИіTBXTЛљвђЃЌВЂдкФкКЌзг5жаШЗЖЈСЫСэвЛИіЗНЯђЯрЗДЃЈЕЙжУЃЉЕФAluдЊМўЁЃгЩгкНќДІЕЙжУЕФ Alu ађСаПЩвдХфЖдВЂВњЩњЫЋСД RNA НсЙЙЃЌвђДЫзїепЬсГіЃЌЮЛгкСНИі Alu ађСажЎМфЕФЭтЯдзг 6 ПЩФмдкзЊТМКѓжБНгДг RNA зЊТМБОжаШЅГ§ЃЌетИіЙ§ГЬГЦЮЊМєНгЃЈЭМ 1ЃЉЁЃ
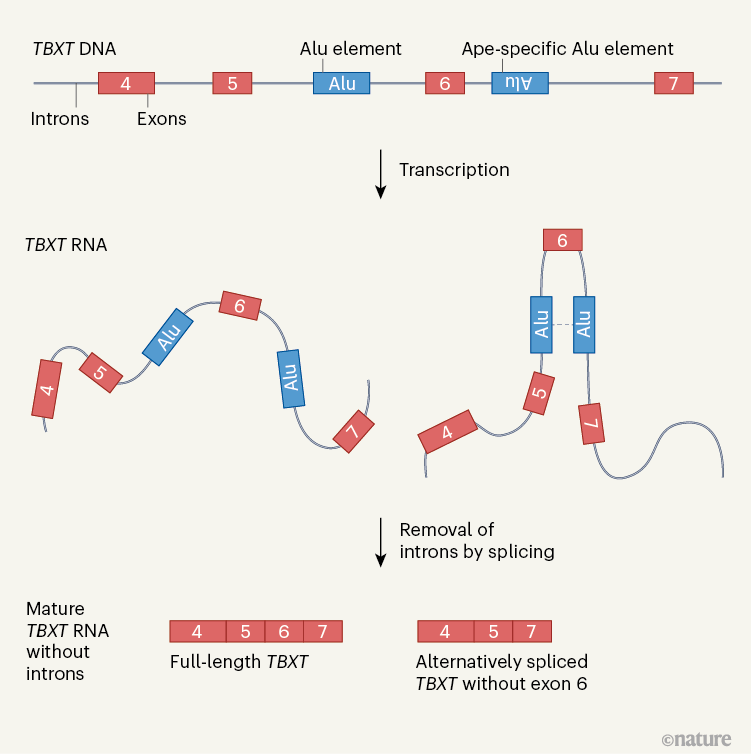
ЭМ 1 |ЮВВПЗЂг§ЛљвђЕФРрШЫдГЬивьадЬцДњ RNA зЊТМБОЁЃ TBXTЛљвђВЮгыЮВВПЗЂг§ЁЃЯФЕШШЫ . 3 БШНЯСЫСщГЄРрЖЏЮяЕФTBXTађСаЃЌВЂМјЖЈГівЛИіЖЬЕФвЦЖЏDNAађСаЃЌГЦЮЊAluдЊМўЃЌИУађСадкРрШЫдГжаВхШыTBXTЕФЗЧЕААзжЪБрТыЧјЃЈФкКЌзгЃЉЃЌЕЋдкЦфЫћСщГЄРрЖЏЮяжадђВЛВхШыЁЃЕБ DNA БЛзЊТМГЩ RNA ЪБЃЌетИі Alu дЊМўгыСэвЛИіИННќЕФ Alu дЊМўЕФЯрЛЅзїгУЃЌЫќВЛЪЧРрШЫдГЬивьадЕФЃЌЖјЪЧЯрЗДЕФЗНЯђЃЌПЩвдЕМжТдкМєНгЙ§ГЬжаШЅГ§ЕААзжЪБрТыЧјЃЈЭтЯдзгЃЉЃЌДгЖјВњЩњГЩЪь RNA ЕФСНжжПЩФмАцБОЁЊЁЊвЛжжЪЧШЋГЄЕФЃЌСэвЛжжЪЧШБЪЇЭтЯдзг 6ЁЃетжжЭтЯдзгЬјЙ§ЕФTBXTЕФБэДяПЩФмЕМжТдчЦкРрШЫдГНјЛЏЪБЮВАЭЕФЩЅЪЇЁЃ
ЮЊСЫШЗЖЈетСНИіAluађСаЪЧЗёВњЩњСЫTBXT RNAзЊТМБОЕФПЩБфМєНгАцБОЃЌXiaМАЦфЭЌЪТНЋШЫРрКЭаЁЪѓХпЬЅИЩЯИАћНјааЗжЛЏЃЌвдБуЫќУЧФЃФтTBXTБэДяЕФЗЂг§зДЬЌВЂВЮгыЮВВПЗЂг§ЁЃаЁЪѓЯИАћЕФЛљвђзщВЛАќКЌСщГЄРрЖЏЮяЬивьад Alu ВхШыЃЌНіБэДяШЋГЄ Tbxt зЊТМБОЃЌЕЋШЫРрЯИАћЭЌЪББэДяШЋГЄзЊТМБОКЭВЛАќРЈЭтЯдзг 6 ЕФНЯЖЬзЊТМБОЁЃЪЙгУЛљвђБрМЙЄОп CRISPR-Cas9 ШЅГ§ШЮвЛ Alu дЊМўЕМжТШБЗІЭтЯдзг 6 ЕФзЊТМБОМИКѕЭъШЋЖЊЪЇЁЃ
ЮЊСЫНЋИФБфЕФзЊТМБОгыдкРрШЫдГжаЙлВьЕНЕФЮоЮВБэаЭСЊЯЕЦ№РДЃЌXia ЕШ ДДНЈСЫМИЬѕЛљвђаЁЪѓЁЃЦфжавЛИіаЁЪѓФЃаЭжЛЪЧЩОГ§СЫ Tbxt ЕФЭтЯдзг 6ЁЃЮЊСЫШЗШЯЗНЯђЯрЗДЕФИпЖШЯрЫЦЕФађСаЃЈЗДЯђЛЅВЙађСаЃЉПЩФмЕМжТбЁдёадМєНгЃЌДгЖјЕМжТЭтЯдзг6ЕФЬјдОЃЌЫћУЧЛЙЭЈЙ§аоИФTbxtЭтЯдзг6ВрвэЕФФкКЌзгађСаДДНЈСЫЁАШЫдДЛЏЁБаЁЪѓФЃаЭЁЃЫћУЧЭЈЙ§ећКЯвЛЖдЗДЯђЛЅВЙЕФAluађСаЛђвЛЖдЗДЯђЛЅВЙЕФаЁЪѓЬивьадађСаРДзіЕНетвЛЕуЁЃ
зїепжЄЪЕЃЌШБЗІЙІФмадШЋГЄTbxt RNAзЊТМБОЕФаЁЪѓЮоЗЈДцЛюЕНГіЩњ 6 , 7 ЃЌВЂЗЂЯжОпгавЛИіЭъећЛљвђПНБДКЭвЛИіИФБфПНБДЕФаЁЪѓОпгаПЩБфЕФБэаЭЃЌДгЮоЮВЕНОпгаШЋГЄЮВАЭЁЃШЛЖјЃЌОпгаЗДЯђЛЅВЙ Alu ађСаВрвэЭтЯдзг 6 ЕФШЫдДЛЏаЁЪѓФЃаЭУЛгаЮоЮВЛђЖЬЮВБэаЭЁЃетОЭЬсГіСЫвЛИіЮЪЬтЃЌМДЮВАЭЕФЫ№ЪЇЪЧЗёШЗЪЕЭъШЋЪЧгЩРрШЫдГЬигаЕФAluВхШыЧ§ЖЏЕФЃЌЛђепЪЧЗёашвЊЦфЫћДйГЩвђЫиЁЃгаШЄЕФЪЧЃЌВхШыаЁЪѓЬивьадЕФЗДЯђЛЅВЙађСаШЗЪЕЕМжТСЫаЁЪѓЕФЮВАЭЫѕЖЬБэаЭЁЃЯФЕШШЫ .гаСЫСэвЛИіСюШЫаЫЗмЕФЗЂЯжЃКЫљгаОпгавЛИіTbxtПНБДЕФаЁЪѓЃЌЦфжаЭтЯдзг6БЛЩОГ§ЃЌвЛИіПНБДВхШыСЫЗДЯђЛЅВЙаЁЪѓЬивьадађСаЃЌЖМУЛгаЮВАЭЁЃ
змжЎЃЌетаЉЪ§ОнжЇГждГРрЬивьадAluВхШыдкДйНјдГРрЮоЮВБэаЭжаЕФзїгУЁЃДЫЭтЃЌзїепЙлВьЕНЃЌБэДяИпЫЎЦНЕФЭтЯдзгЬјдОЛљвђзЊТМЮяЕФаЁЪѓдкХпЬЅНсЙЙжаГіЯжШБЯнЕФЗчЯедіМгЃЌХпЬЅНсЙЙКѓРДаЮГЩДѓФдКЭМЙЫшЃЌГЦЮЊЩёОЙмЁЃвђДЫЃЌзїепЬсГіСЫвЛжжПЩФмадЃЌМДЮвУЧРрШЫдГзцЯШЕФЮВАЭЖЊЪЇПЩФмАщЫцзХЩёОЙмШБЯнЗчЯеЕФдіМгЁЃ
ФЧУДЃЌЮЊЪВУДдчЦкЕФРрШЫдГЛсЪЇШЅЮВАЭФиЃПвЛаЉбаОПШЫдБНЋетжжЫ№ЪЇНтЪЭЮЊЪЪгІадЫ№ЪЇЃЌетвтЮЖзХЫќНЋЬсЙЉНјЛЏгХЪЦЁЃетЪЧЯФЕШШЫЕФЯыЗЈЁЃвВВЮгыЃЌКєгІвдЧАЕФЯыЗЈЃЌМДЮВАЭЕФЩЅЪЇгажњгкИФЩЦСНЬѕЭШЃЈЫЋзуЃЉдЫЖЏЁЃЖдШЫРрЙ§ЖЩЮяжжЕФбаОПБэУїЃЌСНзуЖЏЮязюГѕЪЧдкЮвУЧЕФЪїЦмзцЯШжаНјЛЏЖјРДЕФЃЌКѓРДБЛгУгкЕиУцЦмЯЂЕФЩњЛюЗНЪН 8 .ПЦбЇМвУЧЧуЯђгкЙизЂЮВАЭЖЊЪЇЕФЪЪгІадНтЪЭвдМАЫќШчКЮЪЙШЫРрФмЙЛвЦЖЏЃЌЕЋгаМИЬѕжЄОнБэУїЃЌгЕгаЮВАЭВЂВЛЗСАСНзуЖЏЮяЃЌЪЕМЪЩЯПЩвджЇГжЫќЁЃР§ШчЃЌЮВАЭЫЦКѕгажњгкБЃГжОэЮВКяЃЈ Sapajus libidinosusЃЉдкдЫЪфЪЏЦїКЭЫЋзуаазпЪБЕФзЫЪЦ 9 .ОЁЙмШЫРрОГЃЫЋзудЫЪфЛѕЮяЃЌЕЋЛњЦїШЫбаОПБэУїЃЌбќВПАВзАЕФЁАЮВАЭЁБПЩвддіМгЮШЖЈад 10 ЃЌетвтЮЖзХЮВАЭЩѕжСПЩвдЪЪгІЯжДњШЫРрЁЃ
СщГЄРрЖЏЮяжжШКЕФЮяРэИєРыЬсЙЉСЫСэвЛжжНтЪЭЁЃДѓдМ2500ЭђФъЧАдкЖЋЗЧПЊЪМЕФЙЙдьЛюЖЏЃЌАщЫцзХЛ№ЩНЛюЖЏЁЂКўВДПЊЗЂКЭКгСїЭјТчЕФжизщЃЌЕМжТСЫЦјКђКЭзЪдДПЩгУадЕФЯдзХБфЛЏЁЃетаЉЕиРэБфЛЏПЩФмгыдчЦкРрШЫдГПЊЪМНјЛЏЕФЭЌЪБЗЂЩњ 11 .дчЦкЕФРрШЫдГзцЯШПЩФмвђЮЊЦјКђОчБфЖјБфЕУЙТСЂЁЃдкжжШКЙцФЃНЯаЁЕФЧщПіЯТЃЌЫцЛњвХДЋЦЏвЦ - Р§ШчXia ЕШШЫБЈЕРЕФAluВхШыЕФЙЬЖЈЁЃЁЊЁЊБОРДПЩвдЗЂЛгБШбЁдёИќДѓЕФзїгУ ЮВАЭЫ№ЪЇ 12 .вђДЫЃЌдчЦкРрШЫдГжаTBXTЛљвђЙІФмЕФИФБфПЩФмЪЧгЩгквЛИіаЁЕФЁЂЩњжГИєРыЕФжжШКжаЕФвХДЋЦЏвЦЃЌзїЮЊвЛжжЪЪгІадЗДгІЃЌЛђСНепМцЖјгажЎЁЃ
ОЁЙмзюжеЕФвђЙћЙиЯЕПЩФмШдШЛВЛПЩжЊЃЌЕЋXiaМАЦфЭЌЪТЕФбаОПНсЙћЮЊЮвУЧЕФЮВАЭЙЪЪТЬсЙЉСЫСюШЫаХЗўЕФаТЦЊеТЃЌВЂШЗЖЈСЫзЊзљдЊМўШчКЮгажњгкШЫРрЛљвђБэДяПтЕФЖрбљЛЏЃЌВЂзюжеДйНјЕфаЭЕФШЫРрЬиеїЁЃ
https://blog.sciencenet.cn/blog-41174-1423518.html
ЩЯвЛЦЊЃКАБЛљЫсзѓа§ЪжадЕФЦ№дД
ЯТвЛЦЊЃКаЧаЮНКжЪЯИАћЖдааЮЊЕФЕїНкзїгУ
