博文
生物群落之波动及其演替
||
一. 生物群落的波动

1. 生物群落的波动(Fluctuation)
群落短期的可逆的变化。

2. 波动的原因
群落的波动多数是由群落所在地区气候条件的不规则变动引起的,生物本身的活动周期和人为活动也会引起波动。群落发生波动时,群落区系成分(Floristic component)的相对稳定性,群落数量特征变化的不定性以及变化的可逆性。

3. 波动的类型
3.1 不明显(不显露)波动(Obsolete fluctuation):
种群间的数量变化较小,群落的外貌、结构特征基本保持不变。这种波动可能出现在不同年份的气象、水文状况差不多一致的情况下。

3.2 摆动性波动(Oscillating fluctuation):
种群个体数量(Number of individuals)、密度(Density)、盖度(Coverage)、生产力(Productivity)等数量特征发生短期(1-5 a)的波动。这种波动往往与群落中优势种(Dominant species)的逐年更替有关。

3.3 偏途性波动(Plagiosere fluctuation):
由于气候、水文条件较长期的偏离均态而引起优势种群消长置换形成的一种较长周期(5-11 a)的植被动态现象,波动幅度的大小受气候、水文中长周期波动因素制约也与太阳黑子(Macula)活动周期有一定联系。

4. 波动的特征
木本植物占优势的群落较草本植物稳定一些。
常绿木本群落要比夏绿木本群落稳定一些。

在一个群落内部,许多定性特征(如种类组成、种间关系、分层现象等)较定量特征(如密度、盖度、生物量等)稳定一些。
成熟的群落较发育中的群落稳定。

不同的气候带(Climate zone)内,群落的波动性不同,环境条件越是严酷,群落的波动性越大。

二. 生物群落的演替

1. 生物群落的演替(Succession)
指在群落发展变化过程中,由低级到高级、由简单到复杂、一个阶段接着一个阶段,一个群落代替另一个群落的自然演变现象。
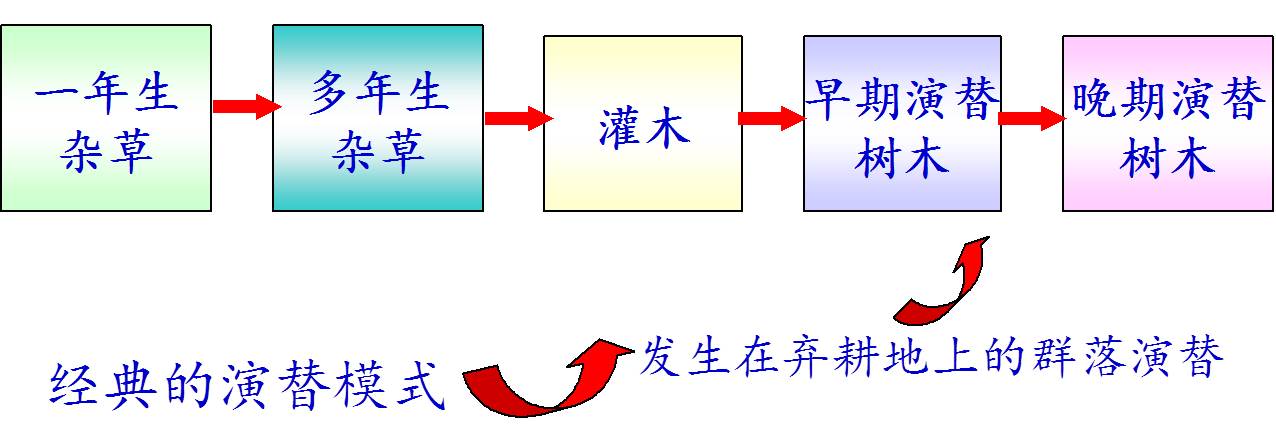
2. 演替的类型
2.1 按群落起始条件划分:
a. 原生演替(Primary succession):
从未有过任何生物的裸地(Primary bare area)上开始的演替,如在裸露的岩石、在河流的三角洲、冰川、火山喷发区等所开始的演替。
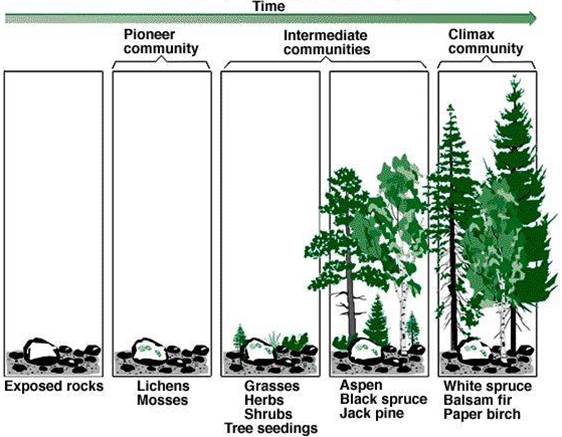

b. 次生演替(Secondary succession):
在原有生物群落被破坏后的次生裸地(Secondary bare area,如森林砍伐迹地、弃耕地)上开始的演替。在这种情况下,演替过程不是从一无所有开始的,原来群落中的一些生物和有机质仍被保留下来,附近的有机体也易侵入。
因此,次生演替比原生演替更为迅速。




2.2 按演替发生的时间进程划分:
a. 快速演替:
即在时间不长的几年内发生的演替。如地鼠类的洞穴、草原撂荒地上的演替,在这种情况下很快可以恢复成原有的植被。但是要以撂荒地面积不大和种子传播来源就近为条件,否则草原撂荒地的恢复过程就可能延续达几十年。



b. 长期演替:
延续的时间较长,几十年或有时几百年,如云杉林等被采伐后的恢复演替。

c. 世纪演替:
延续时间相当长久,一般以地质年代计算。常伴随气候的历史变迁或地貌的大规模改造而发生。

2.3 按引起演替的主导因素划分:
a. 内因生态演替或内因动态演替(Endogenic succession):
由于群落中生物的生命活动结果导致的演替。群落中生物的生命活动结果首先使它的生境发生改变,然后被改变了的生境又反过来作用于群落本身,如此相互促进,使演替不断向前发展。一切源于外因的演替最终均通过内因性演替来实现。所以,内因性演替是群落演替的最基本和最普遍的形式。
b. 外因生态演替或外因动态演替(Exogenic succession):
由于外界环境因素的作用所引起的演替,如气候发生演替、地貌发生演替、土壤发生演替、火成演替、人为发生演替等。

2.4 按基质的性质划分:
a. 水生基质演替(Hydrosere):
开始于水生环境中,但一般都发展到陆地群落,如淡水或池塘中水生群落向中生群落的转变过程。

b. 旱生基质演替(Xerosere succession):
从干旱缺水的基质上开始,如裸露的岩石表面或沙丘上生物群落的形成过程。

2.5 按群落代谢特征划分:
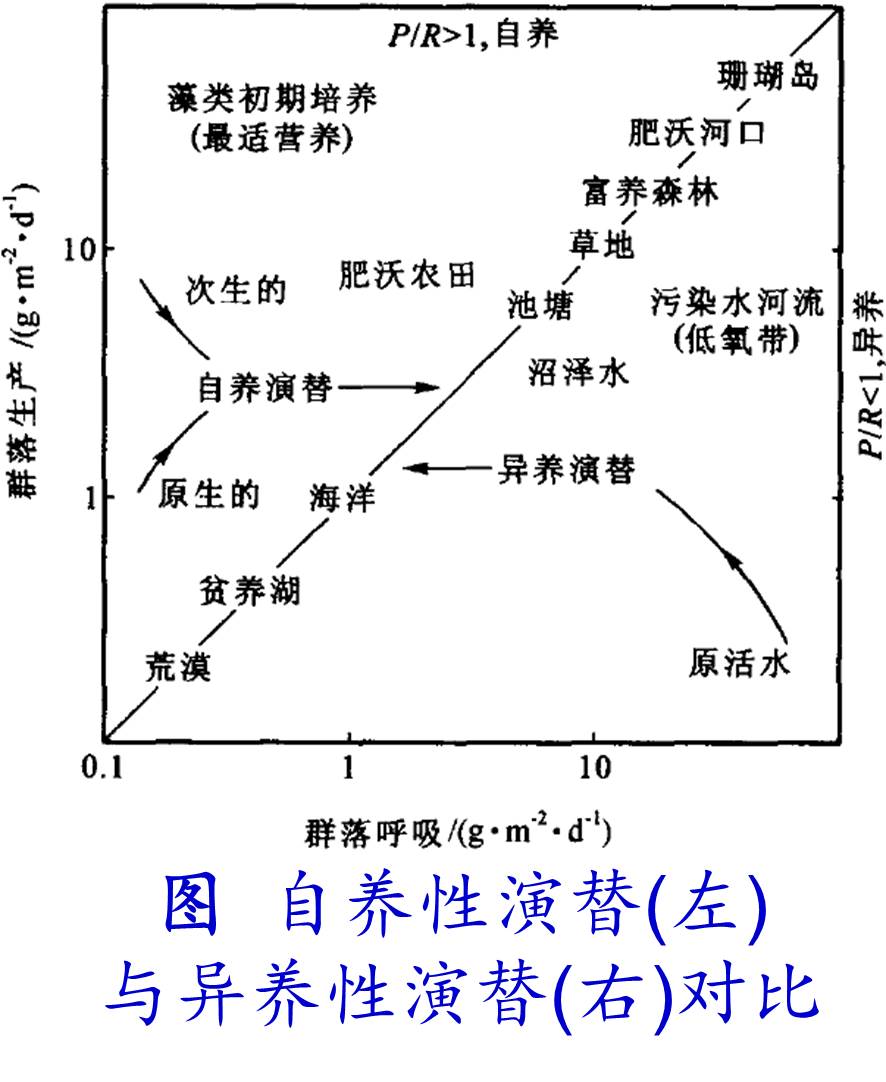
a. 自养性演替(Autotrophic succession):
在演替过程中,群落的初级生产量(P)超过群落的总呼吸量(R),即P/R>1,群落中的能量和有机物逐渐增加。例如陆地从裸地→地衣、苔藓→草本→灌木→乔木的演替过程中,光合作用所固定的生物量越来越多。
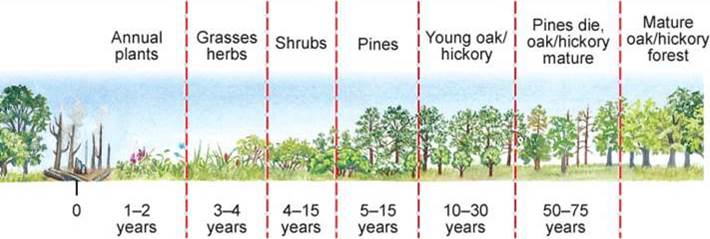
b. 异养性演替(Heterotrophic succession):
在演替过程中群落的生产量少于呼吸量,即P/R<1,表明群落中能量或有机物在减少。异养型演替多见于受污染的水体,如海湾、湖泊和河流受污染后,由于微生物的强烈分解作用,有机物质随演替而减少。

3. 演替系列
3.1 演替系列(Successionsere):
生物群落的演替过程,从植物的定居开始,到形成稳定的植物群落为止的过程。

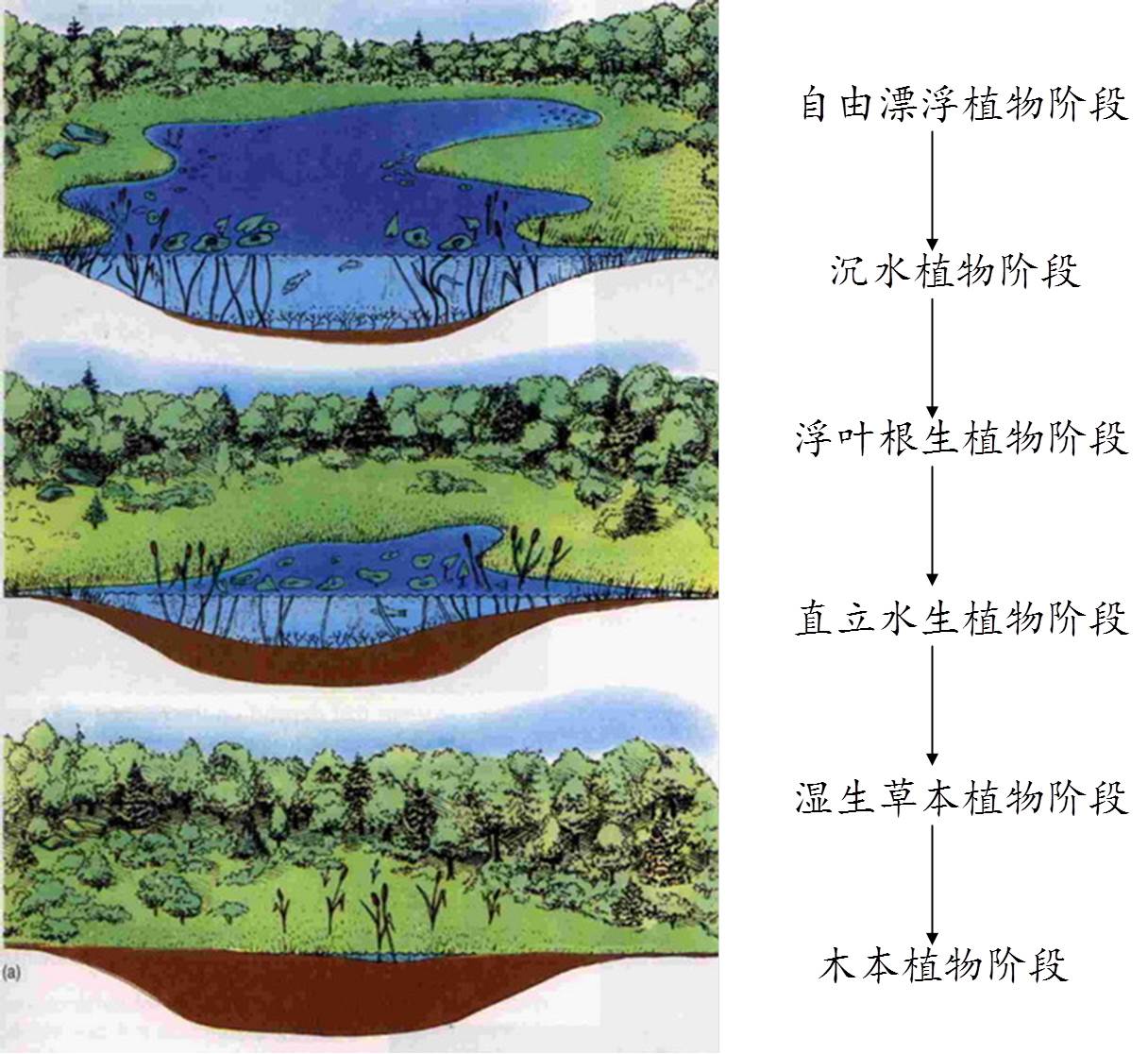
3.2 水生演替系列(Hydrosere):
a. 自由飘浮植物阶段:有机质的聚集靠浮游有机体的死亡残体,以及湖岸雨水冲刷所带来的矿质微粒,导致湖底增高。
b. 沉水植物阶段:5-7 m水深,首先出现轮藻属植物,由于它的生长,湖底有机质积累较快而多,由于湖底嫌气条件轮藻的残体分解不完全,湖底进一步抬高;水深2-4 m左右,有金鱼藻、狸藻等出现,繁殖强,垫高湖底。
c. 浮叶根生植物阶段:水深1 m左右,睡莲等植物飘浮水面,导致水下的沉水植物得不到光照而被排挤,飘浮植物的茎部的阻碍,更多泥沙沉积下来,同时植物残体量更大,湖底抬高,有利于下一阶段植物入侵。
d. 直立植物阶段:水变浅,芦苇、香蒲等个体更大,突出水面,枝叶茂密,根常纠缠绞结,拦截泥沙能力更强,残体也更多,水更浅,使湖底迅速升高。
e. 湿生草本植物阶段:水变成季节性积水,根茎发达的湿生的沼泽植物开始生长。如莎草科、禾本科等一些湿生种类。排水能力更强和垫高能力更强。
疏林阶段:耐水湿的灌木、乔木出现,如柳、赤杨等。
f. 中生森林:随树木的侵入,形成森林。地下水位降低,大量地被物改变了土壤条件。
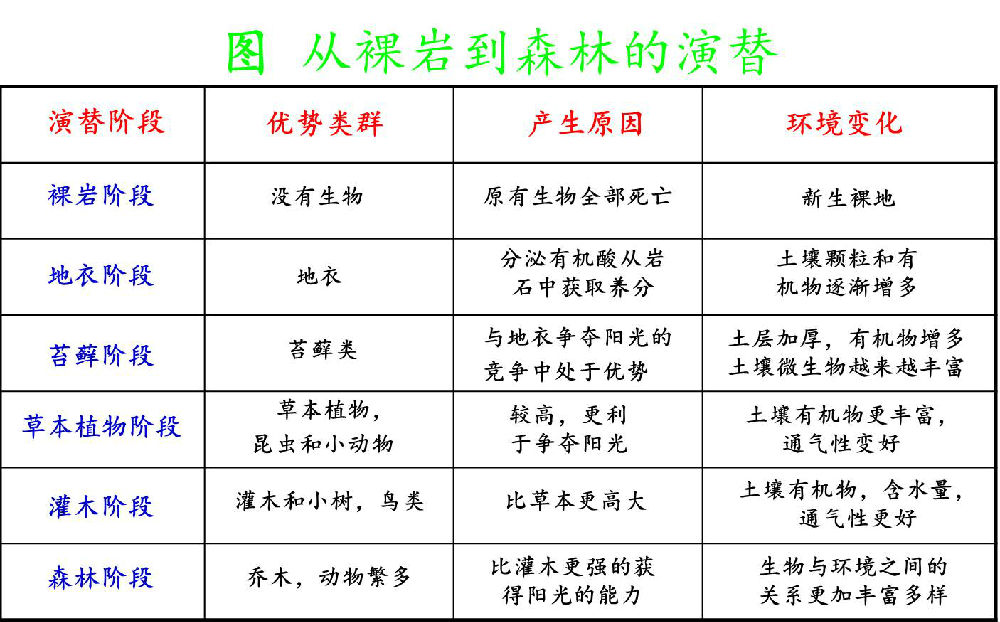
3.3 旱生演替系列(Xerosere):
a. 地衣植物阶段:壳状地衣分泌有机酸腐蚀岩石表面,加上岩石风化作用,壳状地衣的一些残体,逐渐形成一些极少量的土壤叶状地衣:可含蓄较多的水分,积聚更多的残体,使土壤增加的更快些。叶状地衣把岩石表面遮盖部分,生长枝状地衣,生长能力强,全部代替叶状地衣。
b. 苔藓植物阶段:在干旱时进入休眠,待到温和多雨时大量生长。积累的土壤更多,为以后生长的植物创造条件。
c. 草本植物阶段:蕨类、一年生、二年生植物,低小耐旱种,取代苔藓植物,土壤增加、小气候形成、多年生草本出现,使土壤增厚、遮荫、减少蒸发,土壤中真菌、细菌和小动物增多。
d. 灌木群落阶段:喜光的阳性灌木出现,与高草混生形成“高草灌木群落”,以后灌木大量增加,形成优势灌木群落。
e. 乔木群落阶段:阳性乔木树种生长,逐渐形成森林,林下形成荫蔽环境使耐荫树种定居,随着耐荫种的增加,阳性树种在林内不能更新而逐渐从群落消失。林下生长耐荫的灌木和草本植物复合的森林群落形成。







4. 演替方向
4.1 进展演替(Progressive succession):
指随着演替的进行,生物群落的结构和种类成分由简单到复杂;群落对环境的利用由不充分到充分利用;群落生产力由低到逐步增高;群落逐渐发展为中生化;生物群落对外界环境的改造逐渐强烈。

4.2 逆行演替(Regressive succession):
在不利的自然因素和人为因素(如污染和过牧)干扰下,生物群落的演替也可向反方向进行,使群落逐渐退化,即导致生物群落结构简单化;不能充分利用环境;生产力逐渐下降;不能充分利用地面;群落旱生化;对外界环境的改造轻微。


多数群落的演替有一定的方向性,但也有一些群落有周期性的变化,即由一个类型转变为另一个类型,然后又回到原有的类型,称周期性演替(Periodic succession),如石楠→石蕊→熊果→石楠。

5. 演替机制
5.1 经典的演替观
每一演替阶段的群落明显不同于下一阶段的群落。

前一阶段群落中的物种活动促进了下一阶段物种的建立。

5.2 个体论演替观
Egler (1952)提出初始物种组成决定群落演替系列中后来优势种的学说,
Connell & Slatyer (1977)提出3种可能的物种取代机制。

5.3 演替机制
a. 促进模型(Facilitation model)
物种替代是由于先来物种的活动改变了环境条件,使它不利于自身生存,而促进了后来物种的繁荣。因此,物种的替代有顺序性、可预测和具方向性。
多出现在环境条件严酷的原生演替中。
b. 抑制模型(Inhibition model)
先来物种抑制后来物种,使后者难以入侵和发育,故物种替代无固定次序,各种可能均有,其结果很大程度上取决于哪一种先到。
演替在更大程度上决定于个体的生活史对策,因而难以预测。在该模型中没有一个物种可被认为是竞争的优胜者,而是决定于先到该地,故演替往往是从短命种到长命种,而不是由规律、可预测的物种替代。

c. 耐受模型(Tolerance model)
介于上述二者之间,认为物种替代决定于物种的竞争能力。先来的机会种在决定演替途径上并不重要,任何物种均可能开始演替,但有一些物种竞争能力优于其它种,因而它最后能在顶极群落中成为优势种。

3类模型的共同点:演替中的先锋物种最先出现,它们具有生长快、种子产量大、有较高的扩散能力等特点。这类易扩散和移植的物种一般对相互遮荫和根间竟争的环境是不易适应的,故在3种模型中,早期进入物种均是较易被挤掉的。

三种模型的区别:重要的是演替的机制,即物种替代的机制,是促进或抑制,亦或是现存物种对替代影响不大,而演替机制取决于物种间的竞争能力。

d. 适应对策演替理论(Adapting strategy theory,Grime 1989):
R对策种适应于临时性资源丰富的环境。C对策种生存于资源一直处于丰富状态下的生境中,竞争力强,称为竞争种。S对策种适用于资源贫瘠的生境,忍耐恶劣环境的能力强,为耐胁迫种。Grime (1988)提出R-C-S对策模型反映了某一地点某一时刻存在的植被是胁迫强度、干扰和竞争之间平衡的结果。该学说认为,次生演替过程中的物种对策格局是有规律的、可预测的。一般情况下,先锋种为R对策,演替中期的多为C对策,而顶级群落中的种则多为S对策种。
e. 资源比率理论(Resource ratio hypothesis,Tilman 1985):
一个种在限制性资源比率为某一值时表现为强竞争者,而当限制性资源比率改变时,因为种的竞争能力不同,组成群落的植物种也随之改变。因此,演替是通过资源的变化而引起竞争关系变化而实现的。

f. 等级演替理论(Hierarchical succession theory,Piclett 1987):
演替的一般性原因:即裸地的可利用性、物种对裸地利用能力的差异、物种对不同裸地的适应能力。
将以上的基本原因分解为不同的生态过程,如裸地可利用性决定于干扰的频率和程度,种对裸地的利用能力决定于种的繁殖体生产力、传播能力、萌发和生长能力等。
演替的机制水平,包括立地-种的因素和行为及其相互作用,这些相互作用是演替的本质。
6. 演替顶级学说(Climaxtheory)
6.1 单元顶极学说(Monoclimax hypothesis,F.E.Clements 1916)
演替顶极(Climax)和顶极群落:任何一类演替均经过迁移、定居、群聚、竞争、反应、稳定6个阶段,当群落达到与周围环境取得平衡时(物种组合稳定),群落演替渐渐变得缓慢,最后的演替系列阶段称演替顶极(Successionclimax),演替最后阶段的群落称顶极群落(Climax community)。
a. 气候顶极群落:Clements认为,在任何一个地区内,一般的演替系列的终点决定于该地区的气候性质,主要表现在顶极群落的优势种,能很好地适应于地区的气候条件,这样的群落称之为气候顶极群落(Climatic climax)。
在同一气候区内,无论演替初期的条件多么不同,植被总是趋向于减轻极端情况而朝向顶极方向发展,从而使得生境适合于更多的植物生长。
在一个气候区内,除了气候顶级之外,还会出现一些由于地形,土壤或人为等因素所决定的稳定群落阶段,即前顶级(Pre-Climax):
亚顶级:气候顶级以前的一个相当稳定的阶段,如内蒙古大针茅草原顶级前的羊草草原。
偏途顶级:由于强烈而频繁的干扰因素所引起的相对稳定的群落阶段,如内蒙古草原由于强烈干扰而长期停留在冷蒿阶段。
先顶级:由于局部条件较好而产生的稳定群落阶段,如草原中局部出现的草甸或森林。
超(后)顶级:由于局部条件较差而产生的稳定群落阶段,如草原中局部出现的荒漠植被阶段。
但这些顶级最终将发展成为气候顶级阶段。
6.2 多元顶极学说(Polyclimax theory,A.G. Tansley1954)
若一个群落在某种生境中基本稳定,能自行繁殖并结束其演替过程,即为顶极群落。

在一个气候区域内,群落演替的最终结果不一定均汇集于一个共同的气候顶极终点。

任何一个区域的顶极群落均是多个的,均是由一定环境条件所控制和决定的,如土壤湿度、土壤营养特性、地形和动物活动等,这些群落即为地形顶极(Topographic climax)、土壤顶极(Edaphic climax)和动物顶极(Zootic climax)。同时还存在一些复合型顶极,如地形-土壤顶极(Topoedaphic climax)和火烧-动物顶极(Fire-zootic climax)等。

单元顶极学说和多元顶极学说的差异:
相同点:①均承认顶极群落是经过单向变化而达到稳定状态的群落。②顶极群落在时间上的变化和空间上的分布均是和生境相适应的。

不同点:①单元顶极论认为只有气候才是演替的决定因素,其他因素均是第二位的,但可以阻止群落向气候顶极发展。多元顶极论则认为,除气候以外的其他因素,也可以决定顶极的形成。②单元顶极论认为,在一个气候区域内,所有群落均有趋同性的发展,最终形成气候顶级。而多元顶极论不认为所有群落最后都会趋于一个顶级。

6.3 顶极格局学说(Climax-pattern hypothesis,R.H. Whittaker 1953)
在任何一个区域内,环境因子均是连续不断地变化的。随着环境梯度的变化,各种类型的顶极群落(如气候顶极、土壤顶极、地形顶极等),不是截然呈离散状态,而是连续变化的,形成连续的顶极类型(Continuous climax type),构成一个顶极群落连续变化的格局。
在这个格局中,分布最广泛且通常位于格局中心的顶极群落,叫做优势顶极(Prevailing climax),它是最能反映该地区气候特征的顶极群落,相当于单元顶极论的气候顶极。

6.4 顶级演替学说(Climax succession hypothesis,R.H. Whittaker 1974)
一个顶极群落具有如下特征:
群落中的种群处于稳定状态。
达到演替趋向的最大值,即群落总呼吸量与总第一性生产量的比值接近。
与生境的协同性高,相似的顶极群落分布在相似的生境中。
不同干扰形式和不同干扰时间所致的不同演替系列都向类似的顶极群落会聚。
在同一区域内具最大的中生性。
占有发育最成熟的土壤。
在一个气候区内最占优势。
7. 群落顶级演替的功能过程变化
7.1 顶极群落的能量学(Energetics)特征

7.2顶极群落的的结构和生活史(Lifecycle)特征

7.3 顶极群落的物质循环(Material cycle)和内稳定性(Homeostasis)特征

注:囿于个人水平之限,错误在所难免。敬请批评指正!此外,本博文图片和部分文字介绍均源于网络,版权归原作者和(或)出版商,仅用于学术传播等非商业行为。特在此致谢!
https://blog.sciencenet.cn/blog-565899-1192867.html
上一篇:百颗毳毬谁织就 几枝琼萼露和圆
下一篇:南通濠河之风光