博文
2020年值得关注的科学技术
|||
| 思想领袖们对可能在2020年产生重大影响的科技进展做出预测。
更高品质的冷冻电镜样品
王宏伟
清华大学结构生物学家
我认为透射冷冻电子显微镜(cryo-EM)将在两到三年内成为破译大分子结构的最有力工具。大分子结构对于理解生化机制和药物开发具有至关重要的作用,而更加有效的解析方法可以加快这些工作。
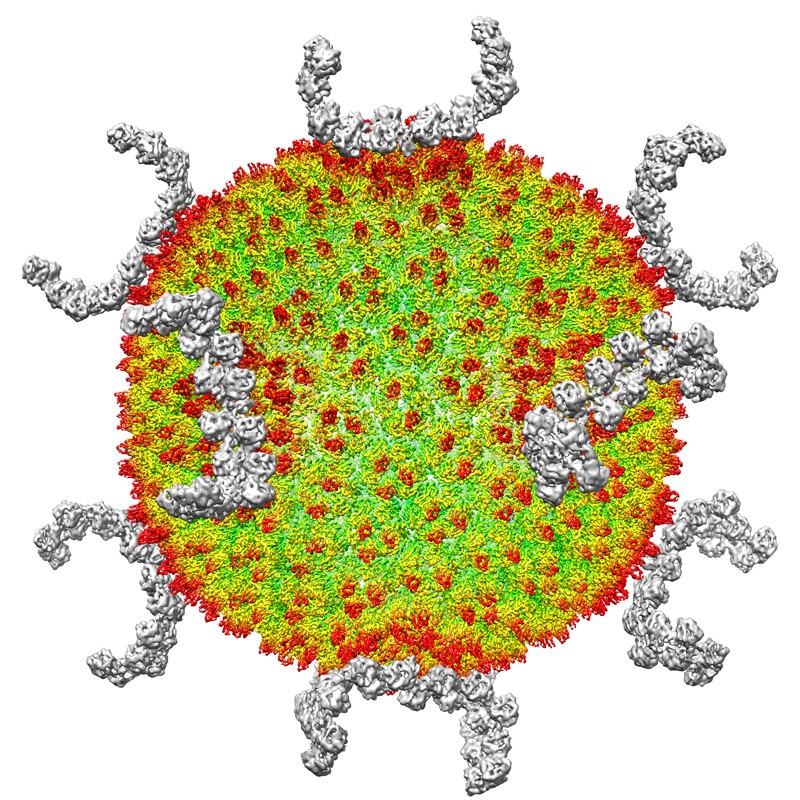
根据冷冻电镜图像重建的SH1病毒。图片来源:Luigi De Colibus/University of Oxford
对冷冻电镜而言,在液氮中快速冷冻生物样本有助于保存分子的含水量,并减少用于成像的高能电子所造成的损害。但是样品制备是一个主要的瓶颈:如果没有好的样品,就更谈不上成像。生物样品通常含有蛋白质,会在冷冻过程中所用到的薄液体层表面解体。
为了防止蛋白质解体,研究人员正在开发这样一类方法:加入液滴之前,将蛋白质固定在二维材料上——比如碳晶格石墨烯。通过这种方式,他们可以使液滴更小,同时可以避免蛋白质接触空气-水界面[1]。
一些实验室将纳升级样品直接放在某种表面上[2],而不是使用繁琐的旧方法从较大的液滴中吸走多余的液体。另一类方法是用聚焦离子束将冷冻细胞切成小于100纳米的薄层,这样研究人员就可以研究细胞内的分子结构[3]。
用冷冻电镜解析分子结构通常需要收集和分析多达10,000张图像,这往往需要几周到一个月的工作量。许多图像是有缺陷的,无法使用。但从理论上讲,几十张照片就足够了,而且收集和分析这些照片需要不到一天的时间。处理能力的提升,可以帮助我们了解疾病机制并且更有效地开发药物。
优化RNA分析
Sarah Woodson
约翰斯·霍普金斯大学生物物理学家
我一直在关注长读段RNA测序,以及基于发光RNA链适配子的活细胞成像。这些技术仍在不断完善,但我预计未来一两年将发生重大变化。
短读段测序改变了RNA生物学领域,比如它可以鉴定哪些RNA序列包含化学修饰位点。然而,长读段测序(例如,Oxford Nanopore和Pacific Biosciences公司提供的测序技术)目前能够用于确定特定修饰在细胞中的普遍性,以及RNA分子某一部分的变化是否与其他部分的变化相关。
发光适配子是实验室合成的单链DNA或RNA分子,能够与荧光染料结合。这些适配子与某些海洋动物产生的绿色荧光蛋白类似,与染料结合时,其荧光强度会增强。这使得研究人员能够追踪某些过程,例如,驱动神经退行性疾病的细胞内RNA簇的形成。
早期的发光适配子是不可靠的,只能产生微弱的信号,甚至有时根本不起作用,因为与目标RNA融合时序列会发生错误折叠。但一些研究团队已经开发出了新型荧光RNA,我在论文和汇报中已经看到了为提高现有适配子亮度和创造不同颜色荧光变体所做的巨大努力。
我的实验室使用化学足迹法来研究细胞内的RNA折叠。许多疾病都与RNA结构的改变有关,但这种改变难以确定。现在我们转而利用长读段测序和发光适配子来研究癌症、代谢综合征和阿尔茨海默病等疾病中的RNA-蛋白聚集。利用这些技术,我们可以更好地将细胞死亡和其他疾病特征与细胞内RNA分子的变化联系起来。
解析微生物组
Elhanan Borenstein
以色列特拉维夫大学计算系统生物学家
在过去的十年中,微生物群落的基因测序方法被用于探索人类微生物组的组成。最近,科学家们试图通过整合基因、转录、蛋白质和代谢物相关信息,来了解微生物组的活动,其中代谢物备受关注:它们有望让我们最直接了当地理解微生物如何影响机体健康,因为多种宿主-微生物互作是通过细菌产生和消耗代谢物发生的。
微生物组-代谢组研究呈爆炸式增长,以一系列粪便样本为例,这些研究通过宏基因组测序确定每个样本中存在的物种及其丰度,并使用质谱分析和其他技术来测定不同代谢物的浓度。结合这两组数据,我们希望了解微生物群落中的各种组分在做什么,进而判断某特定微生物是否决定了某些代谢物的水平。
但这些数据是复杂且多维的,可能存在一个完整的相互作用网络,涉及多个物种和途径,最终产生了一组代谢物。科学家们发表了一些用于关联微生物组和代谢组数据的计算方法,以了解其中的联系和模式。这些方法包括简单的相关性分析以及复杂的机器学习方法——利用现有的微生物组-代谢组数据集来预测新的微生物群落的代谢组,或重现微生物与代谢物之间的关系。
我们实验室采取了不同的策略。我们没有使用统计方法来发现微生物-代谢物的关联,而是针对我们所认为的特定微生物组成如何影响代谢组的机制建立了模型,并使用这些模型作为分析本身的一部分。实际上,我们的问题是:根据基因组和代谢信息,我们对每种微生物产生或吸收特定代谢物的能力了解多少?然后,我们可以预测特定微生物群体产生或降解特定代谢物的潜力,并将这些预测与实际的代谢组学数据进行比较。我们证明这种方法避免了简单的相关性分析的缺陷[4],并将在接下来的几个月里发布这一分析方法的新版本。
这类研究可以通过识别特定的微生物来改进基于微生物组的治疗方法,例如,识别哪些微生物产生的有害代谢物过多或产生的有益代谢物过少。
计算癌症
Christina Curtis
斯坦福大学计算和系统生物学家
对于癌症而言,我们不能看到疾病形成的过程,只能看到最终的结果:当临床上能够检测到肿瘤时,我们才能对它进行取样。然而此时肿瘤已经发生了许多突变,我们能做的只有去搞清楚发生了什么。

通过代码来建立模拟肿瘤发展的模型。图片来源:Getty
我们的团队建立了一个计算模型,在考虑组织空间结构的同时,探索肿瘤发展的动力学。该模型可以模拟一系列场景并生成“虚拟肿瘤”,其突变模式模拟了患者数据。通过比较模拟数据和实际的基因组数据,可以推断出哪些参数可能引发了病人的肿瘤。
使用新兴的标记和记录方法,将这些推断方法与直接测量肿瘤谱系和表型相结合令我感到兴奋。过去两年所取得的进展包括开发出基于CRISPR的标记,它可以记录哺乳动物发育过程中细胞的命运[5,6]。其他技术通过RNA的原位表达,利用基于图像的DNA标记检测方法,捕捉细胞谱系、空间邻近性和表型信息[7]。
在一项模拟结肠癌肿瘤生长的研究中[8],我们使用肿瘤测序数据和模拟来研究原发性和转移性肿瘤之间的关系。这些推断分析表明,绝大多数的癌症在原发肿瘤只有10万个细胞时已经开始扩散,此时肿瘤还太小而无法用结肠镜等标准诊断方法检测到。
模型和测量方法相结合有着更好的灵敏性和可扩展性,可以跟踪肿瘤形成过程中的谱系和空间关系,促进深入了解癌症的起源,包括特定的突变如何影响细胞适应性并加速疾病进程。
改进基因治疗
Alex Nord
加州大学戴维斯分校遗传学家
我们目前已经进行了15年的大规模实验,绘制参与控制细胞和器官表达基因的增强子和其他调控DNA序列图谱。虽然还需要进行更多的工作来完成这些图谱,但此刻我们已经可以利用已掌握的信息来更精确地控制基因组。
去年10月,在芝加哥举行的神经科学学会(the Society for Neuroscience)年会上,我参与主持了会议的一个专题。该专题聚焦于识别增强子序列,并利用它们来控制大脑中特定细胞类型的基因表达。一种方法是将工程病毒注入大脑,测试数千种相关增强子的基因表达谱。2019年,西雅图艾伦脑科学研究所(Allen Institute for Brain Science)的研究人员利用这一策略在人类大脑皮层的特定位置寻找增强子[9]。此外,哈佛大学的一个研究小组使用基于RNA测序的方法找到了只在特定中间神经元(一种产生神经回路的神经细胞)中起作用的增强子[10]。
增强子序列被识别后,科学家就可以利用它们来驱动特定细胞类型种的基因表达,从而用于基因治疗。在单拷贝基因失活或缺失引发的疾病中,CRISPR-Cas9基因编辑工具可以将转录激活因子定位到基因的增强子上,从而提高基因的表达量。小鼠研究表明,这些方法可以纠正与肥胖和X染色体综合征、Rett综合征和Dravet综合征等疾病相关的基因表达缺陷[11],Dravet综合征是我的实验室正在研究的一种严重的癫痫症。在未来的一年里,我认为我们仍将继续医治小鼠,但这项技术已获得大量产业投资。希望我们可以利用这些方法来改变人类基因治疗的方式。
单细胞测序
J. Christopher Love
麻省理工学院科赫综合癌症研究所化学工程师
我感兴趣的是如何更快、更便捷地为病人提供药物。这需要多层面的技术。一方面,需要有新发现,例如单细胞测序方法。另一方面,则是如何将技术应用到病人身上,即生产过程。这与治疗罕见疾病或小众药物尤其相关,甚至也适用于将我们已有药物推广到全球的过程。
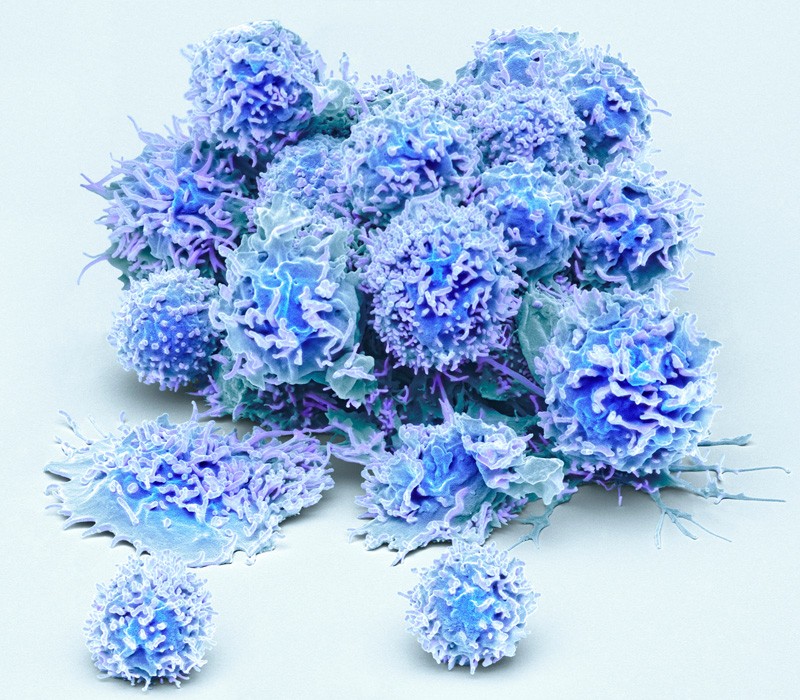
人血液中激活的T细胞。图片来源:Steve Gschmeissner/SPL
在药物发现方面,我们与麻省理工学院的同事合作,为实现高通量的单细胞RNA测序开发了一个便携且便宜的平台[12]。但是,要拥有足够的分辨率来区分免疫细胞亚型,例如具有不同作用和抗原特异性的免疫细胞亚型,仍然是一个挑战。在过去一年左右的时间里,我们以多种方式改进了单细胞基因组测序。首先,我们提出了一种更有效的检测低表达转录本的方法[13]。针对T淋巴细胞,我们特别设计了一种方案,将每个细胞的基因表达谱与其独特的抗原受体序列联系起来[14]。
与此同时,波士顿丹娜-法伯癌症研究所(Dana Farber Cancer Institute)的一个研究团队发表了一项巧妙的文库筛选策略,以解决另一个问题——找出某种特定T细胞受体所识别的抗原[15]。
我与麻省理工学院的Alex Shalek以及其他人合作,创办了蜂巢生物技术公司(Honeycomb Biotechnologies),尝试将我们的单细胞RNA测序平台商业化。不需要在离心机中将细胞离心,只需将其放在试管中,利用液氮冷冻,然后从非洲寄来,也就是说,你只需要寄来一排单细胞大小的孔,约为一个U盘的大小。这使得世界上任何地方的任何样本都可以进行单细胞存储和基因组分析。
将基因组结构和功能联系起来
Jennifer Phillips-Cremins
宾夕法尼亚大学表观遗传学家和生物工程师
倘若把一个单细胞的DNA从头到尾拉直,大约得有2米长,但它却能放在一个直径比大头针针尖还小的细胞核中。DNA的折叠模式不可能是随机的;染色体所形成的三维结构在生物体整个生命周期中都必须受到时空上的调控。
随着过去十年基因组学和成像技术的进步,我们现在可以创建有关基因组折叠的超高分辨率图谱。现在最大的问题是,每种折叠模式的功能是什么?它们如何控制基因表达、DNA复制和DNA修复等基本过程?
有几种合成生物学方法可以让我们在一定长度和时间尺度上折叠和探测基因组。一种方法是CRISPR-GO,它可以将DNA片段携带到细胞核上或细胞核内的特定区域[16]。这将使科学家们可以研究DNA序列在细胞核中的位置如何支配基因功能。
另一种方法是我们实验室的光激活动态循环(LADL)工具,它利用光和CRISPR-Cas9将特定的DNA片段连接在一起,以满足长距离的需要[17]。这可以使增强子直接接触到数千甚至数百万个碱基之外的目标基因,因此我们可以直接评估该调控序列的功能:其目标基因的表达是上调还是下调,以及到什么程度?该技术允许对基因表达进行精确的时空控制,而基因表达在许多疾病中都会被严重扰乱。
第三个系统是CasDrop,它使用另一种光激活的CRISPR-Cas9系统,将特定的DNA片段牵引入亚核的无膜“凝聚体”[18]。自几年前被发现以来,它们在细胞中的功能一直备受争议。
我对未来的设想是,我们可以将这些3D基因组工程工具与基于CRISPR的活细胞成像方法结合起来,以便在细胞中实时地设计和观察基因组。
功能可以驱动结构,或者结构可以驱动功能。这是一个巨大的谜题,未来将由这些工程工具来帮助我们解答。
参考文献:
1.Liu, N. et al. J. Am. Chem. Soc. 141, 4016–4025 (2019).
2.Wei, H. et al. J. Struct. Biol. 202, 170–174 (2018).
3.Schaffer, M. et al. Nature Meth.16, 757–762 (2019).
4.Noecker, C. et al. mSystems https://doi.org/10.1128/msystems.00579-19 (2019).
5.Chan, M. M. et al. Nature 570, 77–82 (2019).
6.Kalhor, R. et al. Science 361, eaat9804 (2018).
7.Askary, A. et al. Nature Biotechnol. 38, 66–75 (2020).
8.Hu, Z. et al. Nature Genet. 51, 1113–1122 (2019).
9.Mich, J. et al. Preprint at bioRxiv https://doi.org/10.1101/555318 (2019).
10.Hrvatin, S. et al. eLife 8, e48089 (2019).
11.Colasante, G. et al. Mol. Ther. https://doi.org/10.1016/j.ymthe.2019.08.018 (2019).
12.Gierahn, T. M. et al. Nature Meth. 14, 395–398 (2017).
13.Hughes, T. K. et al. Preprint at bioRxiv https://doi.org/10.1101/689273 (2019).
14.Tu, A. A. et al. Nature Immunol. 20, 1692–1699 (2019).
15.Kula, T. et al. Cell 178, 1016–1028 (2019).
16.Wang, H. et al. Cell 175, 1405–1417 (2018).
17.Kim, J. H. et al. Nature Meth. 16, 633–639 (2019).
18.Chin, Y. et al. Cell 175, 1481–1491 (2018).
原文以Technologies to watch in 2020为标题发表在2020年1月21日的《自然》技术特写版块
© nature
Nature|doi:10.1038/d41586-020-00114-4
想看更多精彩文章?
扫描下方二维码关注“Nature Research科研服务”服务号

版权声明:
本文由施普林格·自然上海办公室负责翻译。中文内容仅供参考,一切内容以英文原版为准。欢迎转发至朋友圈,如需转载,请邮件China@nature.com。未经授权的翻译是侵权行为,版权方将保留追究法律责任的权利。
© 2020 Springer Nature Limited. All Rights Reserved
https://blog.sciencenet.cn/blog-3432244-1240225.html
上一篇:青年科学家代写同行评议损失甚多
下一篇:快来参加《自然》首次博士后全球调研吧!