博文
胆汁酸决定你是否更容易发胖
 精选
精选
||
胆汁酸决定你是否更容易发胖
肥胖,被人们戏称为“一种工伤”,是我们现代社会高强度工作状态下的一个普遍现象。“工伤”型肥胖通常指向心性肥胖,即四肢纤细,腰腹脂肪堆积,源于饮食不当、工作压力及睡眠不足,荷尔蒙分泌紊乱。肥胖除了会增加许多疾病包括心血管疾病、2型糖尿病、肝脏疾病、睡眠呼吸中止症、癌症、退行性关节炎、抑郁等的发病风险外,还会导致疾病的症状加重,预后更差。最近几个月肥胖患者们被新冠(COVID-19)虐了一把,成为一批重点受害人群。因此,这样的“工伤”一旦发生,后果很严重。前一阵子有一封辞职信在网上刷屏,辞职理由是:长胖了24公斤,要回家减肥。
但是,阻碍着我们在法律上把肥胖定义为“工伤”的一个重要因素,是我们的工作场所里还有那么一些怎么吃都胖不起来的人。他们跟我们坐一个办公室,干一样的活,却能随意吃喝甚至胡吃海喝而身材不受任何影响。这种让人羡慕无比(当然有时候也恨得牙痒痒)的人类,我们在科研中称之为肥胖抵抗(obesity-resistant)型;而与之相对应的,所谓“喝口水都能胖起来”的容易受伤的我们,则称为肥胖易感(obesity-prone)型。当然这样来区分也是相对的。我们做科学研究的,不会相信真有绝对吃不胖的人,也不相信只喝点白水就真的会让人胖起来。
有趣的是动物中其实也有同样的现象。很早以前科研人员就发现,同一种属的啮齿动物在接受相同的饮食喂养后,动物之间会出现两种体重表型,即部分动物产生明显的肥胖,而另一些动物的体重增长相对较慢。2006年的时候,我们组的博士研究生李后开对相同遗传背景以及同性别年龄的实验鼠研究发现,同一组动物中肥胖易感型的与肥胖抵抗型的动物的代谢组有差异,而差异代谢物的大多数来自于菌群的代谢产物,提示两种代谢表型存在“细微的”肠道菌群上的差异[1]。由于当时的菌群测序技术和生物信息学尚处于早期发展阶段,尚不成熟,这个工作没有继续往下做。
近两年我们组的魏美林博士研究生对这个现象进行了更深入的研究,他的研究发现,由肠道菌介导的胆汁酸代谢中的一些细微变化,会促使哺乳动物在相同饮食条件下体重增加,发生“肥胖易感”的表型。研究结果在2020年5月在线发表于EbioMedicine [2]。
下面我们来讨论一点生化机制,这里涉及一些专业知识,不同知识背景的读者们请咬紧牙关往下看,如果觉得实在晦涩难懂,就直接跳过,看最后结论。
胆汁酸是在肝脏中合成的胆固醇衍生物,参与了机体中很多重要的生理和代谢反应,除了促进营养物质的肠道吸收外,近年来被发现作为重要的信号调控分子,在糖脂代谢、能量消耗甚至是调节机体免疫功能中均具有重要的作用。胆汁酸的合成有两条通路[3],第一条称为经典途径(classical pathway),由胆固醇7α-羟化酶(CYP7A1)起始,再经由胆固醇12α-羟化酶(CYP8B1)控制。另一条称为替代途径(alternative pathway),由胆固醇 27-羟化酶 (CYP27A1)起始,然后由氧固醇 7α-羟化酶(CYP7B1)控制 ,主要负责合成非12位羟基( non-12-OH)胆汁酸 ,包括 CDCA及起源于 CDCA的胆酸如 UDCA和 LCA及它们的结合型胆汁酸,在大鼠、小鼠中还包含 MCA等胆汁酸 。经典途径中CYP8B1是合成CA的关键酶,控制着CA/CDCA的比例[4]。机体内12位羟基(12-OH)胆汁酸主要是CA以及肠道菌利用CA产生的DCA及它们的甘氨酸、牛磺酸结合形式,因此CYP8B1对体内12-OH胆汁酸的合成具有重要的调控作用。小鼠中CDCA合成后可以通过CYP2C70迅速转变为MCA类胆汁酸[5]。无菌小鼠能够抵抗高脂饮食诱导的肥胖,有意思的是无菌小鼠体内(主要来自于替代合成途径的)non-12-OH胆汁酸显著高于常规饲养的小鼠[6]。而cyp8b1敲除的小鼠亦能抵抗高脂饮食诱导的肥胖[7]。综上研究结果提示,胆汁酸谱组成成分的变化可能在肥胖的代谢表型中起重要的作用,且可能和肠道菌密切相关。目前对于不同肠道菌介导的胆汁酸谱的变化以及与肥胖易感及肥胖抵抗表型的关系尚不清楚。
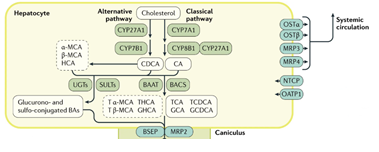
图1. 胆汁酸的合成途径[2]
我们在临床上比较了两组肥胖人群,一组是代谢方面比较健康的,另一组是患有2型糖尿病等代谢疾病的肥胖者。我们研究发现,non-12-OH胆汁酸比例在代谢健康的肥胖人群中显著高于有代谢疾病的肥胖患者,提示non-12-OH 胆汁酸与肥胖的代谢状态密切相关。我们进一步研究了高脂饮食肥胖易感及高脂饮食肥胖抵抗的两组小鼠(即相同喂养条件下体重增加较快的和较慢的两部分小鼠),分析其胆汁酸差异,发现了类似的结果:相比肥胖易感小鼠,体重增长缓慢的小鼠体内拥有更高的non-12-OH胆汁酸水平,且这些小鼠的代谢紊乱程度较轻。体重增长较快的小鼠体内的non-12-OH胆汁酸如UDCA,LCA和CDCA水平显著减少。胆汁酸谱的这些变化与小鼠肝脏中CYP8B1及CYP7B1的表达调控及及其调控因子如肠道及肝脏中FXR信号有关。其次,我们通过小鼠的肠道菌宏基因组数据分析,发现这些小鼠的肠道菌存在着显著差异,定殖其中差异最显著的一个菌株后可明显增加其体内non-12-OH胆汁酸水平。小鼠胆汁酸谱的改变伴随着回肠组织内胰高血糖素样肽-1(GLP-1)表达及棕色脂肪组织产热基因表达改变。最后,利用这两组小鼠中差异最显著的胆汁酸-熊去氧胆酸(UDCA)干预肥胖易感型小鼠,发现UDCA摄入可以明显增强non-12-OH胆汁酸的组成并减轻高脂饮食诱导的体重增加及其代谢紊乱,提示由肠道菌群介导的胆酸成分的改变通过影响胆汁酸信号途径改变机体代谢及对肥胖的易感性。胆汁酸合成途径(在经典途径和替代途径之间)的平衡可能是调控肥胖代谢的重要靶点,修饰胆汁酸组分可能是调控肥胖代谢的重要干预手段,可以成为一种肥胖预防和治疗的新策略。
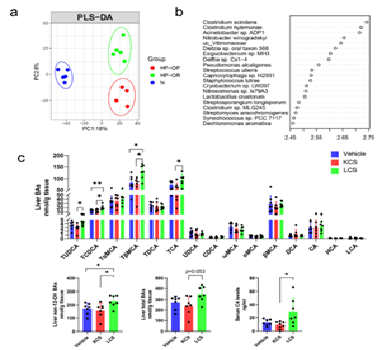
图2. 利用差异的菌株干预后小鼠体内胆汁酸变化
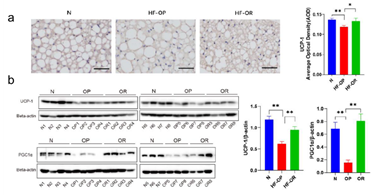
图3. 小鼠棕色脂肪组织UCP1及PGC1α表达水平
该研究中non-12-OH胆酸比例的变化受肝脏中CYP8B1及CYP7B1的表达调控。近年来一些研究表明CYP8B1表达减少或者下调可能对宿主代谢产生有益的影响。例如,cyp8b1-/-小鼠可通过减少脂质吸收来抵抗高脂饮食诱导的肥胖及脂肪肝和胰岛素抵抗的发生[7]。cyp8b1基因缺失会通过增加GLP-1的分泌来改善小鼠的葡萄糖耐量[8]。既往有研究提示肝脏的一个长链非编码RNA可以显著下调CYP8B1的表达,从而增加结合性的MCA/CA比率,进一步增强apoC2的表达并进一步改善脂质代谢[9]。CYP8B1的关键产物CA,作为一种主要的12-OH胆汁酸成分,是一种能够高效形成乳糜微粒的胆汁酸,在促进胆固醇在肠道的吸收中具有重要的作用。基于这个原因,CA被添加在高胆固醇饲料中(致结石饮食,含0.2% 胆固醇和0.5% CA),用于诱导产生高胆固醇血症及结石发生的小鼠模型。而改变胆汁酸组分(增加MCA和UDCA的比例)能够使胆汁酸池亲水性增加,从而导致脂肪吸收减少[10]。另外,胆汁酸替代合成途径中CYP7B1也被证实在代谢的调控中发挥重要的作用。有研究报道,小鼠在寒冷环境暴露试验中会触发新的代谢机制并增加能量消耗,CYP7B1在该机制中起重要的作用[11]。在肥胖的二型糖尿病患者中肝脏CYP7B1表达水平降低,并且在小鼠中敲除cyp7b1会显著减少棕色脂肪组织中的UCP1表达。相反,使用腺病毒过表达肝脏中的cyp7b1能增加棕色脂肪中的UCP1表达及能量消耗。
胆汁酸调控机体的代谢主要是通过胆汁酸受体实现的,目前研究的比较多胆汁酸受体是TGR5及FXR。当能量摄入超过能量消耗时,导致能量代谢发生失衡时,肥胖就容易发生。因此增加能量消耗是控制肥胖的有效策略。已有研究表明UCP1活性对小鼠肥胖的发展具有决定性的作用,而胆汁酸可以调控UCP1的表达。例如,CDCA,LCA和其它TGR5激动会增加棕色脂肪细胞中UCP1的表达,表明胆汁酸受体TGR5对介导这一过程起着重要的作用[12]。给予TGR5的特异性激动剂INT-777只能增加tgr5+/+小鼠棕色脂肪的线粒体数量,而在tgr5敲除小鼠中则无此效果,说明此过程是依赖于TGR5实现的[13]。另外,LCA作为CDCA肠道菌的代谢产物,是TGR5最有效的天然激动剂[14],LCA等胆汁酸可以通过刺激肠道内分泌L细胞TGR5受体增强GLP-1的释放进一步调控机体代谢[15]。
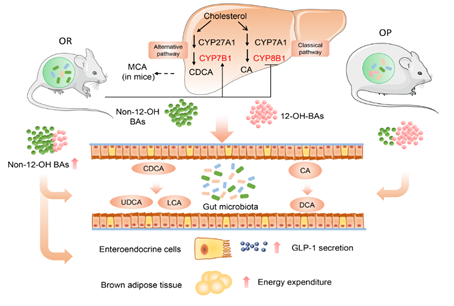
图4:胆汁酸-肠道菌代谢影响肥胖易感表型的大致机制
最后总结一下,肥胖易感和肥胖抵抗小鼠肠道菌群存在着差异,这个差异的“净结果”使得两种小鼠体内的胆汁酸种类有显著差异,肥胖易感者体内12-OH类的胆汁酸比例较高,而肥胖抵抗者体内非12-OH类的胆汁酸高。这样的胆汁酸差异对肥胖表型的影响有两个方面。一方面,12-OH类的胆汁酸(尤其是结合型的12-OH如TCA)在肠道内能够很好地形成乳糜微粒,更有效地吸收脂肪,帮我们增肥;而相反,非12-OH类的胆汁酸能更有效地消耗脂肪和能量,帮我们减肥。这么一来一去,体重差异就显现出来了,就出现了“吃不胖”和“一吃就胖”的两种“体质”。
看到这里,你也许要问,除此以外,还有没有别的生化机制决定着我们肥胖“抵抗能力”上的差异呢?应该还有,这个研究领域目前是一块“热土”,很多新机制正不断被发现!最后我们免费帮各家茶商打个广告,我们组去年发表的普洱茶降脂研究中发现,自愿者喝了普洱茶,非12-OH类的胆汁酸升高,而喝普洱茶的小鼠实验结果也证实了这个发现。看来喝茶是一个很不错的“改良”肥胖易感的养身方法哦!
[1]Li H, Ni Y, Su M, et al. Pharmacometabonomic phenotyping reveals different responses to xenobiotic intervention in rats. J Proteome Res 2007;6(4):1364-70.
[2]Wei M, Huang F, Zhao L, et al. A dysregulated bile acid-gut microbiota axis contributes to obesity susceptibility. EBioMedicine 2020;55:102766.
[3]Jia W, Xie G, Jia W. Bile acid–microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nature reviews Gastroenterology & hepatology 2018;15(2):111.
[4]Li-Hawkins J, Gåfvels M, Olin M, et al. Cholic acid mediates negative feedback regulation of bile acid synthesis in mice. The Journal of clinical investigation 2002;110(8):1191-200.
[5]Takahashi S, Fukami T, Masuo Y, et al. Cyp2c70 is responsible for the species difference in bile acid metabolism between mice and humans. J Lipid Res 2016;57(12):2130-37.
[6]Sayin SI, Wahlström A, Felin J, et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring fxr antagonist. Cell Metab 2013;17(2):225-35.
[7]Bertaggia E, Jensen KK, Castro-Perez J, et al. Cyp8b1 ablation prevents western diet-induced weight gain and hepatic steatosis because of impaired fat absorption. American Journal of Physiology-Endocrinology and Metabolism 2017;313(2):E121-E33.
[8]Kaur A, Patankar JV, de Haan W, et al. Loss of cyp8b1 improves glucose homeostasis by increasing glp-1. Diabetes 2015;64(4):1168-79.
[9]Li P, Ruan X, Yang L, et al. A liver-enriched long non-coding rna, lnclstr, regulates systemic lipid metabolism in mice. Cell Metab 2015;21(3):455-67.
[10]Wang DQ-H, Tazuma S, Cohen DE, et al. Feeding natural hydrophilic bile acids inhibits intestinal cholesterol absorption: Studies in the gallstone-susceptible mouse. American Journal of Physiology-Gastrointestinal and Liver Physiology 2003;285(3):G494-G502.
[11]Worthmann A, John C, Ruhlemann MC, et al. Cold-induced conversion of cholesterol to bile acids in mice shapes the gut microbiome and promotes adaptive thermogenesis. Nat Med 2017;23(7):839-49.
[12]Broeders EP, Nascimento EB, Havekes B, et al. The bile acid chenodeoxycholic acid increases human brown adipose tissue activity. Cell Metab 2015;22(3):418-26.
[13]Velazquez-Villegas LA, Perino A, Lemos V, et al. Tgr5 signalling promotes mitochondrial fission and beige remodelling of white adipose tissue. Nature communications 2018;9(1):1-13.
[14]Kawamata Y, Fujii R, Hosoya M, et al. Ag protein-coupled receptor responsive to bile acids. J Biol Chem 2003;278(11):9435-40.
[15]Pols TW, Noriega LG, Nomura M, et al. The bile acid membrane receptor tgr5: A valuable metabolic target. Dig Dis 2011;29(1):37-44.
https://blog.sciencenet.cn/blog-265898-1238369.html
上一篇:如果人生有四季
下一篇:学会吃饭
全部作者的其他最新博文
- • 学会吃饭
- • 如果人生有四季
- • 普洱茶通过影响肠道菌群和胆汁酸代谢降低胆固醇
- • 城里的月光
- • 华山论剑-2 朋友圈