ВЉЮФ
ЩњЮяЕФбѕЛЏЛЙдЕїНкЁОздШЛЁП  ОЋбЁ
ОЋбЁ
|
Helmut SiesЪЧЕквЛЬсГібѕЛЏгІМЄИХФюЕФбЇепЃЌвВЪЧЕБНёздгЩЛљЩњЮябЇСьгђЕФСьОќШЫЮяЃЌЯждкЕТЙњЖХШћЖћЖрЗђДѓбЇЙЄзїЃЌОГЃаДвЛаЉздгЩЛљЩњЮябЇСьгђЕФзлЪіадЮФеТЃЌетЦЊЮФеТЪЧИеИеГіС§ЕФДѓзїЃЌжЕЕУДѓМвбЇЯАЁЃ
бѕЛЏЛЙдЗДгІЪЧжИЛЏбЇЮяжЪжЎМфЕФЕчзгзЊвЦЗДгІЁЃЫќУЧЪЧЩњУќЙ§ГЬЕФЛљДЁЃЌДгдкВњЩњбѕЦјЕФЙтКЯзїгУжаВЖЛёЬЋбєФмЕНЧ§ЖЏЯИАћДњаЛЁЃИїжжбѕЛЏЛЙдЗДгІдкЫљЮНЕФбѕЛЏЛЙдЕїНкжав§ЕМКЭПижЦЩњЮяЙ§ГЬЁЃбѕЛЏЛЙдЕїНкжаЕФКЫаФзїгУЪЧгЩЕААзжЪжаЕФКЌСђКЭКЌЮјАБЛљЫсВаЛљжДааЕФЃЌЦфБЛЩњРэбѕЛЏМСЃЈШчЙ§бѕЛЏЧтЃЈH2O2ЃЉЃЉПЩФцбѕЛЏЃЌзїЮЊПижЦКЭЕїНкУИЛюадКЭЕААзжЪЙІФмЕФПЊЙиЁЃетаЉбѕЛЏЛЙдПЊЙиЪЙЯИАћКЭЩњЮяЬхФмЙЛЖдИїжжВЛЭЌЕФДЬМЄзіГіЗДгІЃЌР§ШчЩњГЄвђзгЁЂЖОЫиКЭгЊбјЮяжЪПЩРћгУадЕФБфЛЏЁЃ
дкдчЦкПЊДДадЙЄзїЕФЛљДЁЩЯЃЌЗжзгЯИАћЩњЮябЇММЪѕЕФНјВНдкУшЪіЗДгІЮяжжзїЮЊбѕЛЏЛЙдЕїНкЙ§ГЬНщжЪЕФВЛЭЌЙІФмЗНУцШЁЕУСЫОЊШЫЕФНјеЙЁЃБОзлЪіЕФжиЕужївЊЪЧЛюадбѕЃЈROSЃЉАќРЈздгЩЛљКЭЗЧздгЩЛљбмЩњЮяЃЌАќРЈГЌбѕвѕРызгЃЈO2•−ЃЉКЭЙ§бѕЛЏЧтЁЃЮвУЧНЋЬжТлЯпСЃЬхЁЂФкжЪЭјЁЂЙ§бѕЛЏЮяУИЬхЁЂЯИАћКЫЁЂАћжЪШмНКЛђжЪФЄжаЕФбѕЛЏЛЙдЭООЖВњЩњЕФЙ§бѕЛЏЧтШчКЮзїЮЊЕкЖўаХЪЙЃЌЭЈЙ§ЕААзжЪАыызАБЫсВаЛљЕФПЩФцбѕЛЏДЅЗЂЯИАћаХКХЁЃдкЩњЮяЬхЫЎЦНЩЯЃЌХпЬЅЗЂЩњКЭзщжЏЗЂг§ЁЂЯИАћЧЈвЦЁЂЩёОЛюЖЏЁЂЩЫПкгњКЯЕШЙ§ГЬЖМашвЊЙ§бѕЛЏЧтаХКХЁЃжЕЕУзЂвтЕФЪЧЃЌЛюадЕЊЃЈRNSЃЌР§ШчЃЌвЛбѕЛЏЕЊ•NOЃЉЁЂЛюадСђЃЈRSSЃЛР§ШчЃЌСђЛЏЧтH2SЃЉЁЂЧзЕчЪдМСЃЈRESЃЛР§ШчЃЌ4-єЧЛљ-2-ШЩЯЉШЉЃЉЛђєЪЛљЕФЗДгІадЮяжЪвВЗЂЛгаХКХДЋЕМзїгУЃЌЦфжаЕФУПвЛжжЖМУшЪіСЫОпгаВЛЭЌаджЪКЭЯИАћЯрЛЅзїгУЕФЙуЗКЛЏбЇЮяжЪ8ЃЌЕЋНЋВЛНјааЯъЯИЬжТлЁЃ
БОзлЪіЕФЗЖЮЇНіЯогкВИШщЖЏЮяЯИАћДњаЛЃЌУЛгаЯъЯИАќРЈЩњЮябѕЛЏЛЙдПЦбЇЕФЦфЫћДѓСьгђЃЌШчжВЮяЁЃЮвУЧЭЈЙ§ЖдбѕЛЏМСЃЈзюживЊЕФЪЧЙ§бѕЛЏЧтЃЉЕФгаЛњЮяМфКЭЯИАћМфаХКХДЋЕМЬиадМАЦфдкПижЦЯИАћЙІФмжаЕФживЊадЬсГізлКЯЙлЕуЃЌЬжТлСЫбѕЛЏЛЙдЩњЮябЇСьгђЕФаТНјеЙЁЃЮвУЧЬжТлСЫбѕЛЏЛЙдЫГЪЦЖЏСІбЇЕФзюаТИХФюЃЌЦфЬиеїЪЧСЌајЕФбѕЛЏЛЙдДЋИаЁЂбѕЛЏЛЙдаХКХзЊЕМЕФМЄЛювдМАЫцКѓЕФаХКХзЊЛЏЮЊЖржжРраЭЕФЯИАћгІМЄЗДгІЕФМЄЛюЁЃетаЉЗДгІЩцМАбѕЛЏЛЙдЕїНкЕААзЃЈМДбѕЛЏЛЙдЕААзжЪзщЃЉЛюадЕФБфЛЏЃЌВЂзюжеЕМжТБэЙлвХДЋбЇОАЙлКЭЛљвђБэДяЕФБфЛЏЁЃЮвУЧНЋЬжТлhormesisЃЈРДздЯЃРАгяhormaeinЃКЁАЦєЖЏЁБЃЉЃЌЫќУшЪіСЫБЉТЖгкЕЭМССПгІМЄдДЃЈР§ШчбѕЛЏМСЃЉКѓЕФећЬхЩњЮяЪЪгІадЗДгІЃЌВЂЬНЫїЗЧБрТыRNAЃЈncRNAЃЉдкбѕЛЏЛЙдЕїНкжаЕФаТШЯЪЖзїгУЁЃЮвУЧНЋНщЩмгыЭтВПЛЗОГЕФЖрЗНУцЯрЛЅзїгУЕФаТжЊЪЖЃЌЭГГЦЮЊБЉТЖЃЌзюКѓПМТЧбѕЛЏЛЙдвНбЇЕФЗЂеЙСьгђЃЌВЂЖдЯжгаЕФжЊЪЖВюОрНјааеЙЭћЁЃ
вЛЁЂбѕЛЏЛЙдЮШЬЌЖЏСІбЇгыбѕЛЏЛЙдУмТы
бѕЛЏЛЙдЮШЬЌжИЕФЪЧЮЌГжбѕЛЏМСКЭПЙбѕЛЏМСжЎМфЦНКтЕФзДЬЌЁЃОЁЙмЙ§СПЕФбѕЛЏМСВњЩњЛсЕМжТЯИАћЩњЮяЗжзгЕФбѕЛЏЫ№ЩЫВЂв§Ц№бѕЛЏгІМЄЃЌЕЋШБЗІзуЙЛЕФбѕЛЏМСЫЎЦНЛсЫ№КІЙиМќЕФаХКХДЋЕМЙ§ГЬВЂЕМжТЛЙдгІМЄ20ЁЃбѕЛЏМСКЭПЙбѕЛЏМСжЎМфЕФЦНКтЭЈЙ§бѕЛЏЛЙдЕїПиЛёЕУЃЌАќРЈбѕЛЏЛЙдИагІЁЂбѕЛЏЛЙдаХКХДЋЕМЁЂбѕЛЏЛЙдЯьгІКЭЗДРЁПижЦЭООЖЃЈМћЭМ1ЃЉЁЃбѕЛЏаоЪЮЕФЛЙдЪЧгЩNADPHКЭNADHЕФЛЙдСІЧ§ЖЏЕФЃЌЫќУЧГфЕБЁАРэЯыЕФПЩРЉЩЂЕФСНЕчзгзЊвЦЛѕБвАќЁБЃЌвдЮЌГжЪ§АйжжВЛЭЌЕААзжЪКЭаЁЗжзгЕФбѕЛЏЛЙдзДЬЌЁЃNADPHгыЯИАћЕААзлЯЛљЃЈЯИАћбѕЛЏЛЙдаоЪЮЕФжївЊФПБъЃЉжЎМфЕФСЊЯЕЪЧЭЈЙ§СђбѕЛЙЕААзКЭЙШызИЪыФЃЈGSHЃЉЯЕЭГНщЕМЕФЃЌЗжБ№гЩСђбѕЛЙЕААзЛЙдУИКЭGSHЖўСђЛЏЮяЛЙдУИДпЛЏ21ЁЃ
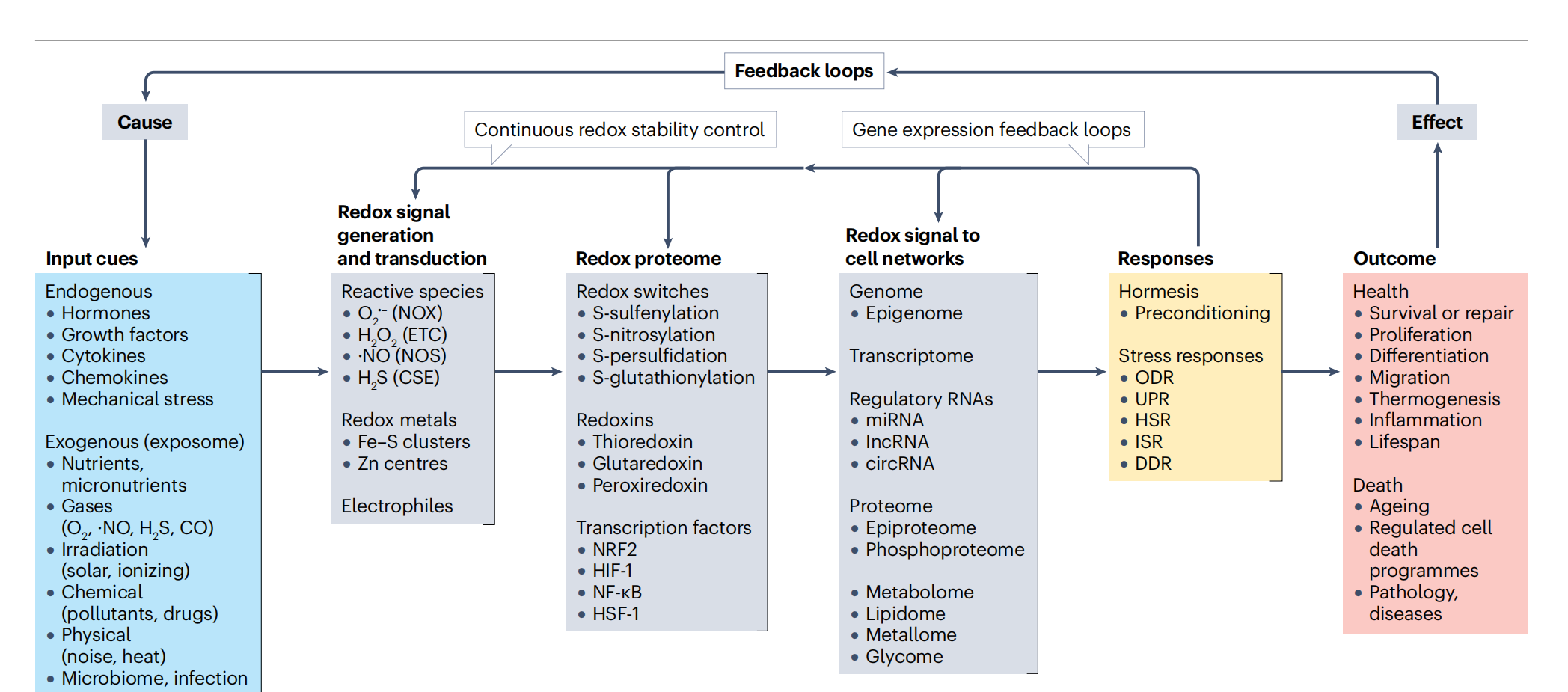
ЭМ1ЃКбѕЛЏЛЙдЕїПидРэЁЃ
бѕЛЏЛЙдЕїПиЕФЙ§ГЬЪМгкГѕЪМЕФвђЙћЪфШыЯпЫїЃЈШчЃЌФкдДадДЬМЄШчМЄЫиЛђЭтдДадЪфШыРДздгЊбјЫиЃЉЁЃФкдДадКЭЭтдДадбѕЛЏЛЙдаХКХЕФВњЩњЕМжТЛюадбѕЮяжжЃЈROSЃЉЁЂЕЊЮяжжЃЈRNSЃЉКЭСђЮяжжЃЈRSSЃЉЕФВњЩњЁЃетаЉбѕЛЏЛЙдаХКХПЩвдЭЈЙ§ГЌбѕвѕРызгЃЈO2•−ЃЉЁЂЙ§бѕЛЏЧтЃЈH2O2ЃЉЁЂвЛбѕЛЏЕЊЃЈ•NOЃЉКЭСђЛЏЧтЃЈH2SЃЉДЋВЅЃЌетаЉЮяжЪЗжБ№гЩNADPHбѕЛЏУИЃЈNOXЃЉЁЂЕчзгДЋЕнСДЃЈETCЃЉКЭЦфЫћFADвРРЕадЯпСЃЬхЭбЧтУИЁЂвЛбѕЛЏЕЊКЯУИЃЈNOSЃЉКЭАыызАБЫсІУ-СбНтУИЃЈCSEЃЉВњЩњЁЃбѕЛЏЛЙдаХКХвВПЩвдЭЈЙ§ЧзЕчЬхЁЂжЌжЪЙ§бѕЛЏЮяКЭЯѕЛљЯЉДЋВЅЁЃбѕЛЏЛЙдаХКХЕФзЊЕМЕМжТбѕЛЏЛЙдЕААзжЪзщКЭДњаЛзщЕФЕїНкЁЃбѕЛЏЛЙдаХКХЭЈЙ§АыызАБЫсПЊЙиЕФЮЛЕуЬивьадбѕЛЏКЭЛЙдДЋВЅЃЌвВГЦЮЊбѕЛЏЛЙдПЊЙиЁЃе§ШчдкКазг2жаЯъЯИЬжТлЕФЃЌбѕЛЏЛЙдПЊЙиЪмЕНПЩФцЕФбѕЛЏКЭЛЙдЃЌЭЈЙ§S-бЧЛЧѕЃЛЏЁЂS-ЙШызИЪыФЛЏЁЂS-бЧЯѕѕЃЛЏЛђЦфЫћбѕЛЏЛЙдУєИаЕФаоЪЮДЋЕнбѕЛЏЛЙдаХКХЁЃбѕЛЏЛЙдаХКХЕФДЋВЅЛЙЭЈЙ§redoxinМвзхЕФУИЃЈСђбѕЛЙЕААзЁЂЙШбѕЛЙЕААзКЭЙ§бѕЛЏЮяЛЙдЕААзЃЉНщЕМЃЌетаЉУИЭЈЙ§ДпЛЏлЯЛљЕФбѕЛЏКЭЛЙдБЛМЄЛюКЭ/ЛђЪЇЛюЁЃжюШчКьЯИАћКЫвђзг2ЯрЙивђзгЃЈNRF2ЃЉЁЂКЫвђзг-ІЪBЃЈNF-ІЪBЃЉЁЂШБбѕгеЕМвђзг1ЃЈHIF-1ЃЉЛђШШанПЫвђзг1ЃЈHSF-1ЃЉЕШгІМЄИагІзЊТМвђзгвВЪЧбѕЛЏЛЙдДЋИаЦїЃЌВЂЭЈЙ§АыызАБЫслЯЛљЕФбѕЛЏКЭЛЙдМЄЛюЛђЪЇЛюЃЌетгАЯьСЫЫќУЧМЄЛюБэДяЫљашвдв§ЗЂЯИАћгІМЄЗДгІЕФЛљвђЕФФмСІЁЃРлЛ§Ц№РДЃЌбѕЛЏЛЙдаХКХЕФДцдкгАЯьДгЛљвђзщКЭЕААзжЪзщЕНЦфЫћзщЕФЯИАћЭјТчЕФдЫзїЃЌЕМжТбѕЛЏЛЙдаХКХЕФећКЯЁЃСэвЛжжбѕЛЏЛЙдЕїПиФЃЪНЪЧгЩИїжжРраЭЕФЕїПиадЗЧБрТыRNAЃЈШчЮЂRNAЃЈmiRNAsЃЉЁЂГЄЗЧБрТыRNAЃЈlncRNAsЃЉЛђЛЗзДRNAЃЈcircRNAsЃЉЃЉЬсЙЉЕФЖрЗНУцЯрЛЅзїгУЁЃМЄЛюИїжжгІМЄЗДгІЃЈШчбѕЛЏгІМЄЗДгІЃЈODRЃЉЁЂЮДелЕўЕААзЗДгІЃЈUPRЃЉЁЂШШанПЫЗДгІЃЈHSRЃЉЁЂзлКЯгІМЄЗДгІЃЈISRЃЉКЭDNAЫ№ЩЫЗДгІЃЈDDRЃЉЃЉЕФМЄЛюаЇгІНсЙћЕМжТдкЪЪгІЁЂЮЌГжЁЂаоИДКЭНЁПЕЛђЯИАћЦЦЛЕЁЂвЦГ§ЛђЫРЭіжЎМфЕФОіВпЁЃГжајдЫзїЕФЗДРЁЛиТЗЭЈЙ§ЕќДњаЕїЕФВЈЖЏКЭеёЕДРДЮЌГжбѕЛЏЛЙдЮШЖЈадЃЌАќРЈЛљвђБэДяЗДРЁЛиТЗЛђзЊТМ-ЗвыЗДРЁЛиТЗЃЌШЛКѓМгЧПЛђМѕШѕбѕЛЏЛЙдаХКХЁЃ
Г§СЫИїжжЭбЧтУИЁЂбѕЛЏУИЁЂбѕЛЏЛЙдУИЁЂМгбѕУИКЭЕчзгзЊвЦИДКЯЮяВњЩњЕФбѕЛЏМСЭтЃЌИїжжРраЭЕФЯИАћДЬМЄЃЌР§ШчЩњГЄвђзгЁЂгЊбјЫиКЭЖОЫиЃЌвВЛсДЅЗЂЪмПиВњЩњвЛЯЕСаЕФбѕЛЏЛЙдЕкЖўаХЪЙЃЌгШЦфЪЧЕЭЩњРэХЈЖШЯТЕФH2O2ЁЂ•NOКЭH2SЃЈЦфЫћЕиЗНзлЪі3,9ЃЉЁЃШЛКѓЃЌЯИАћФкКЭЯИАћМфЦїЙйЕФЭјТчжаЪмбѕЛЏЛЙдЕїПиЕФЕААзжЪаЕїВЂећКЯаХКХвдЮЌГжЩњУќЙ§ГЬЁЃЙЙГЩбѕЛЏЛЙдЯЕЭГзщжЏЕФЛљДЁддђвбБЛжЦЖЈЮЊЁАбѕЛЏЛЙдУмТыЁБ22ЁЃКазг1МђвЊНщЩмСЫетЬззщжЏддђвдМАЕБЧАЛюдОбаОПСьгђЕФвЛаЉаТЪгНЧЁЃЮШЬЌЗЖЪНгыГжајНјааЕФДњаЛСїЕФЮЌГжгаЙиЃКгЩгкЩњУќЙ§ГЬвдЗЧЦНКтШШСІбЇЮЊЬиеї23ЃЌЮЌГжЮШЬЌашвЊВЛЖЯМрВтКЭжиаТБрГЬбѕЛЏЛЙдВЈЖЏЃЌМДЃЌбѕЛЏМСЫЎЦНЕФПеМфКЭЪБМфБфЛЏ24ЁЃетвЛЕуЩѕжСдкИќДѓГпЖШЕФНсЙЙВуУцЩЯСюШЫгЁЯѓЩюПЬЕиЕУЕНСЫР§жЄЃЌМДЯпСЃЬхсеЕФСЌајжиЫмЁЊЁЊгЩФкЯпСЃЬхФЄелЕўаЮГЩЕФёожхЃЌетаЉёожхИЛКЌВњЩњбѕЛЏМСЕФЭбЧтУИКЭЕчзгДЋЕнСДЃЈETCЃЉИДКЯЮяЁЊЁЊдкУыЕФЪБМфГпЖШЩЯЗЂЩњ25ЁЃгЩИУСьгђЕФЯШЧ§епХСРШћЖћЫеЫЙЁЂПЫТхЕТЁЄВЎФЩЕТЁЂЮжЖћЬиЁЄПВХЉКЭТЗЕТЮЌЯЃЁЄЗыЁЄБДЫўРЪЗЦЕШШЫВћЪіЕФОЕфЮШЬЌИХФюЃЌвдФкВПЛЗОГЕФКуЖЈЮЊжааФ26ЁЃШЛЖјЃЌгЩгкЦфЖЏЬЌздЮвзщжЏЕФФмСІЃЌЩњЮяЯЕЭГдкВЛЖЯНјааЕФДњаЛСїжаЪЧЁАздЖЏЬЌЕФЁБ27ЁЃвђДЫЃЌгыЦфЪЙгУбѕЛЏЛЙдЮШЬЌЃЌВЛШчЪЙгУбѕЛЏЛЙдздЖЏЬЌРДУшЪіетвЛЙ§ГЬИќЮЊЧЁЕБЁЃЭМ1УшЪіСЫЯИАћжабѕЛЏЛЙдаХЯЂЕФСїЖЏЃЌЬиБ№ЪЧВЛЭЌЕФЩњРэКЭЛЗОГЯпЫїЭЈЙ§бѕЛЏЛЙдПЊЙиКЭЭјТчЛуМЏЕНЯИАћЩЯЃЌгеЕМЗДгІКЭЙІФмНсЙћЃЌАќРЈЕќДњЗДРЁЛиТЗЃЌШчЛљвђБэДяЗДРЁЛиТЗ28ЁЃРэНтЯИАћШчКЮЯьгІетаЉДЬМЄЪЧвЛИіОпгаЬєеНадЕФШЮЮё29ЃЌетНЋашвЊЖдЩцМАЕФЙ§ГЬгаИќЯъЯИЕФЗжзгВуУцЕФМћНтЁЃШЛЖјЃЌе§ШчЫцКѓЬжТлЕФЃЌзюНќдкбѕЛЏЛЙдЩњЮябЇЗНЗЈЗНУцЕФНјВННвЪОСЫЯИАћШчКЮећКЯИїжжбѕЛЏЛЙдЕїПиЭООЖНјааЯИАћФкКЭЯИАћМфЭЈаХЕФЛњжЦЁЃ
Казг1 ЩњЮябѕЛЏЛЙдЕїПиЕФЛљБОддђЃКбѕЛЏЛЙдУмТы
2015ФъжЦЖЈСЫвЛЬзЩњЮябѕЛЏЛЙдЗДгІдкЛюЯИАћКЭЩњЮяЬхжазщжЏЕФЛљБОдзМдђЃЌГЦЮЊбѕЛЏЛЙдУмТыЃЈВЮПМЮФЯз22ЃЉЁЃетРяМђвЊНщЩмЫФИіддђЁЃШчЦфзюГѕжЦЖЈЪБЫљЪі22ЃЌЫцзХбѕЛЏЛЙдбаОПНвЪОаТЕФМћНтЃЌЫќПЊЗХгкИќаТЃЌзюНќЕФРЉеЙЕНжВЮяЪРНчвВжЄУїСЫетвЛЕу261ЁЃЯрЙибаОПЕФЕБЧАЮФЯздкетРявдЪОЗЖадЕФЗНЪНГЪЯжЁЃ
ЕквЛддђЃКЩњЮяФмСПбЇЁЂЗжНтДњаЛКЭКЯГЩДњаЛЭЈЙ§ЗжБ№ВйзїНгНќгыжабыДњаЛШМСЯЦНКтЕФИпСїСПNADHКЭNADPHЯЕЭГРДзщжЏЁЃ
NADHКЭNADPHЯЕЭГЕФбѕЛЏЛЙдЕїПидкИїжжбЧЯИАћЪвжаЗЂЩњ128ЁЃетаЉИЈУИЯЕЭГдкЯИАћжаЕФКЌСПЭЈЙ§МИжжЗНЪНгЩЕїНкУИПижЦЁЃNAD+МЄУИЃЈNADKЃЉДгNAD+ВЙГфNADP+ЃЈВЮПМЮФЯз262ЃЉЃЌЖјФмСПСЌНгаЭбЬѕЃАЗКЫмеЫсзЊЧтУИЃЈNNTЃЉДгNADHВЙГфNADPH263,264ЁЃNADPHдкбѕЛЏЛЙдЕїПижаОпгаЫЋжиЙІФмЃКЫќЮЊredoxinsЃЈМДСђбѕЛЙЕААзКЭЙШбѕЛЙЕААзЯЕЭГЃЉвдМАЙШызИЪыФЖўСђЛЏЮяЛЙдУИЬсЙЉЛЙдЕШМлЮяЃЌЭЌЪБЭЈЙ§NADPHбѕЛЏУИЃЈNOXЃЉДпЛЏЕФЗДгІНЋЗжзгбѕЛЙдЮЊГЌбѕвѕРызгЖјВњЩњбѕЛЏМСЁЃNAD+ЪЧаэЖрВЮгыЕїНкДњаЛЕФУИЕФЙВЕзЮяЃЌАќРЈsirtuinМвзхЕФЕААзжЪШЅввѕЃЛЏУИ265ЁЃ
ЕкЖўддђЃКДѓЗжзгНсЙЙКЭЛюадЭЈЙ§ЖЏСІбЇПижЦЕФСђПЊЙигыNADHКЭNADPHЯЕЭГЯрСЌЃЌдкбѕЛЏЛЙдЕААзжЪзщжаЁЃ
бѕЛЏМСКЭЧзЕчЬхЕФбѕЛЏЛЙдаХКХжївЊЭЈЙ§ПЩФцЕФЗвыКѓЕААзжЪАыызАБЫсаоЪЮЗЂЩњЃЌетгАЯьСЫДѓЗжзгНсЙЙКЭЛюад13ЁЃбѕЛЏМСЕФбѕЛЏЛЙдаХКХЃЌгШЦфЪЧH2O2ЃЌЭЈЙ§ПЩФцЕФЗжзглЯЛљбѕЛЏЛЙдПЊЙиНјааЁЃетаЉЪЧЬиБ№ЪЪКЯгкПЩФцаХКХзЊЕМЕФЕААзжЪАыызАБЫсКЭМзСђАБЫсВаЛљЃЌвђЮЊЫќУЧЕФЗДгІадЁЂЗсИЛадКЭЮЛжУ169ЁЃЯИАћбѕЛЏЛЙдЕААзжЪзщвдЗжзгЪѕгяУшЪіСЫећЬхЕФбѕЛЏЛЙдзДЬЌЃЌШчЭЈЙ§ЁАOxiMouseЁБЪ§ОнМЏ153ЫљЪОР§ЕФЃЌПЩдкЁЎOxiMouseЁЏЗУЮЪЁЃ
ЕкШ§ддђЃКгыNADHКЭNADPHЯЕЭГЯрСЌНгЕФH2O2ВњЩњМЄЛюКЭЪЇЛюжмЦкжЇГжбѕЛЏЛЙдаХКХДЋЕМКЭЪБПеађЙсЯьгІЃЌгУгкЗжЛЏКЭЖрЯИАћЗЂг§ЁЃ
ФкдДКЭЭтдДЯпЫїдкЬиЖЈЯИАћЮЛжУДЅЗЂбѕЛЏЮяЕФВњЩњЁЃетаЉаХКХзїЮЊЕкЖўаХЪЙДЋЕнжСФПБъЮЛЕуЃЌжївЊЭЈЙ§бѕЛЏЛЙджаМЬЃЌР§ШчРћгУЙ§бѕЛЏЮяЛЙдУИДЋЕнH2O2аХКХЁЃЖдгк•NOЃЌаХКХДЋЕнЭЈЙ§ЯђФПБъРЉЩЂЛђЭЈЙ§S-бЧЯѕЛљЙШызИЪыФЃЈGSNOЃЉНјаазЊбЧЯѕЛљЛЏРДЪЕЯжЁЃ
ЕкЫФддђЃКДгЮЂЛЗОГЕНбЧЯИАћКЭЯИАћзщжЏЕФбѕЛЏЛЙдЭјТчЃЌЙЙГЩСЫвЛИіЪЪгІЯЕЭГвдЯьгІЛЗОГЁЃ
бѕЛЏЛЙдЕїНкаХКХИпЖШећКЯЁЃЧЖШыдкбѕЛЏЛЙдЭјТчжаЕФбѕЛЏЮяЩњВњепЁЂПЙбѕЛЏЯЕЭГКЭбѕЛЏЛЙдУєИагІД№ЦїДйНјЯИАћФкКЭЯИАћМфЕФбѕЛЏЛЙдЭЈаХЁЃЙ§бѕЛЏЕААздЪаэH2O2ДЉЙ§ФЄЃЌетдкЮЌГжЯИАћФкH2O2ЬнЖШЗНУцКмживЊЁЃЛђепЃЌбѕЛЏЮяПЩвдЭЈЙ§МфЯЖСЌНгжБНгдкЯИАћжЎМфвЦЖЏЃЌЛђепЭЈЙ§ЯИАћЭтПеМф136МфНгвЦЖЏЃЌЛђепЭЈЙ§ЯИАћЭтФвХн74ЁЃЪБПебѕЛЏЛЙдПижЦЗЂЩњдкНЋбѕЛЏЛЙдаХКХгыЦфЫћгІМЄЯьгІаХКХЭЈТЗКЭМЄЫивдМАЦїЙйЕНЦїЙйКЭЯИАћЕНЯИАћЕФаХКХзЊЕМСЌНгЦ№РДЕФЕїПиЪрХІ16ЁЃ
ЖўЁЂбѕЛЏЛЙдЮШЬЌЁЂбѕЛЏгІМЄКЭЛЙдгІМЄ
габѕЯИАћДњаЛжабѕЛЏЛЙдЗДгІЕФжївЊЗНЯђЪЧГЏЯђбѕЛЏЃЌетгЩЪЙгУNADHКЭNADPHЕФЛЙдЗДгІРДЦНКтЁЃбѕЛЏгІМЄЕФИХФюЃЈМћВЮПМЮФЯз30,31ЃЉжИЕФЪЧЦЋРыЩњРэадбѕЛЏЛЙдЦНКтЁЃбѕЛЏгІМЄБЛЖЈвхЮЊЁАбѕЛЏЮяКЭПЙбѕЛЏМСжЎМфЕФЪЇКтЃЌЧуЯђгкбѕЛЏЮяЃЌЕМжТбѕЛЏЛЙдаХКХКЭПижЦКЭ/ЛђЗжзгЫ№ЩЫЕФЦЦЛЕЁБ30ЁЃЙизЂЫ№КІЗНУцЕФбаОПвЛжБдкИУСьгђеМОнжїЕМЕиЮЛЃЌжБЕНзюНќЁЃШЛЖјЃЌШЫУЧж№НЅШЯЪЖЕНбѕЛЏгІМЄОпгаЫЋжиадЃКзЂвтСІвбДгГЌЩњРэХЈЖШЕФбѕЛЏЮяЕФгАЯьзЊвЦЕНЩњРэХЈЖШЗЖЮЇФкЕФбѕЛЏЮяЕФзїгУЁЃжЎЧАЬсЕНЕФГжајбѕЛЏЛЙдМрВтЕФЙлФюЕМжТСЫЁАeustressЁБЃЈЯЃРАгяЁАeuЁББэЪОСМКУЕФбЙСІЃЉвЛДЪЕФГіЯжЃЌгУвдБэЪОбѕЛЏЛЙдЕїПижаЕФЖЏЬЌБОжЪЁЃетжже§ГЃЩњРэЦЋВюЕФЗЖЮЇвВБЛГЦЮЊhomeodynamic space32ЁЂН№ЗЂЙУФяЕиДј33ЛђЛЦН№ОљжЕЃЈaurea mediocritasЃЉ34ЁЃH2O2ЪЧбѕЛЏЛЙдЮШЬЌжаЕФКЫаФбѕЛЏЛЙдаХКХЗжзгЃЌЭЈЙ§ЕААзжЪлЯЛљЕФСНЕчзгбѕЛЏРДЗЂЛгзїгУ35ЁЃвЛЕчзгЛњжЦЕФбѕЛЏЛЙдаХКХЃЌМДЭЈЙ§здгЩЛљЃЌвВПЩФмдкЮШЬЌжаЗЂЩњЃЌЕЋеташвЊНјвЛВНЬНЫї36ЁЃЭМ2УшЪіСЫдкЩњРэЯпЫїШчЖЭСЖЛђгЊбјзДЬЌБфЛЏЯТЃЌИажЊбѕЛЏЮявдМЄЛюЖдЮЌГжЯИАћЙІФмЁЂЪЪгІадКЭШЭаджСЙиживЊЕФгавцбѕЛЏЛЙдЮШЬЌаХКХЕФhomeodynamicЗЖЮЇЁЃОЁЙмБОзлЪіжївЊМЏжадкбѕЛЏЛЙдЕїПижаЕФЮШЬЌЩЯЃЌЮвУЧвВНЋМђвЊЬсМАЯИАћЖдГЌЩњРэадбѕЛЏЛЙдЦНКтЦЋВюЃЈМДЃЌбѕЛЏгІМЄЃЉЕФЯьгІЃЌетОпгаЖрЗНУцЕФВЁРэЩњРэбЇКЭвНбЇКѓЙћЁЃЯрЗДЃЌПЙбѕЛЏМСЛђNADPHЕШЛЙдМСЕФЙ§СПвВЛсДЅЗЂГЦЮЊЛЙдгІМЄЕФбЙСІЗДгІЃЌетвВЛсЕМжТЯИАћЙІФмеЯАЃЈЭМ2ЃЉЁЃ
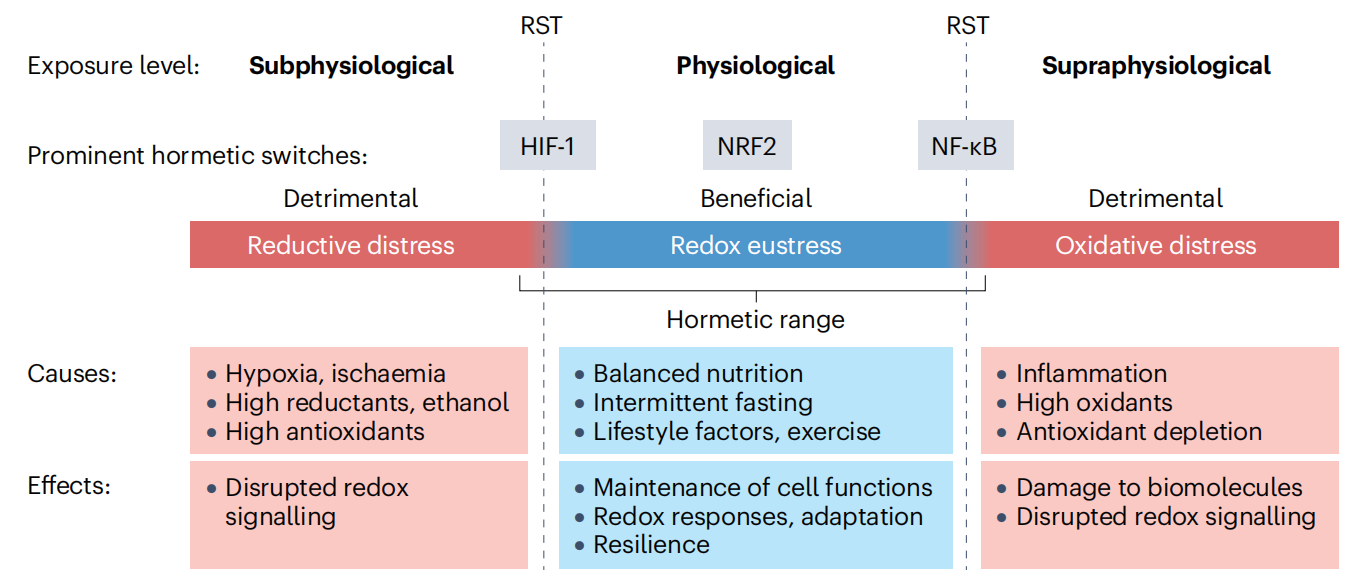
ЭМ2бѕЛЏЮяБЉТЖЕФЗЖЮЇДгзюаЁЃЈЛЙдгІМЄЃЉЕНЩњРэадЃЈбѕЛЏЛЙдЮШЬЌЃЉЕНЗЧГЃИпЕФЫЎЦНЃЈбѕЛЏгІМЄЃЉЁЃЖОЮяаЫЗмаЇгІЗЖЮЇАќРЈДгЩњРэадЃЈРЖЩЋЃЉЕНЗЧЩњРэадЃЈКьЩЋЃЉБЉТЖЫЎЦНЕФЙ§ЖЩНзЖЮЃЌМДЃЌЖОЮяаЫЗмаЇгІМССП-ЗДгІЃЌетЕМжТМЄЛюДйНјЯИАћЪЪгІЕФбѕЛЏЛЙдИагІзЊТМвђзгЁЃДгЩњРэЫЎЦНЕНЕЭгкЩњРэЛђИпгкЩњРэЫЎЦНЕФСйНчуажЕБЛГЦЮЊбѕЛЏЛЙдгІМЄаХКХуажЕЃЈRSTsЃЉ65ЁЃШБбѕгеЕМвђзг1ЃЈHIF-1ЃЉжївЊдкбѕЛЏЮяЫЎЦНЕЭЪБЃЈЧїЯђгкЕЭгкЩњРэЫЎЦНвЛВрЃЉБЛМЄЛюЃЌЖјКЫвђзг-ІЪBЃЈNF-kBЃЉдкЧїЯђгкИпгкЩњРэЫЎЦНвЛВрЦ№зїгУЃЌКЫвђзгКьЯИАћЩњГЩЫи2ЯрЙивђзгЃЈNRF2ЃЉжївЊКИЧЩњРэЗЖЮЇЁЃЭМЕзВПСаГіСЫв§Ц№бѕЛЏЛЙдЮШЬЌКЭСНжжаЮЪНбѕЛЏЛЙдгІМЄЕФвЛаЉжївЊдвђМАЦфЫцКѓдкжїЮФжаЯъЯИЬсЕНЕФгАЯьЁЃ
ЃЈзїепгУШ§жжбѕЛЏзДЬЌЖдбѕЛЏЛЙдЦНКтНјааСЫШЋУцзмНсЃЌЕЭбѕЛЏЪЧЛЙдЃЌИпбѕЛЏЙ§ЖШЃЌе§ГЃбѕЛЏЃЌЕЭбѕЪЧРфзДЬЌЕЭбѕгеЕМвђзгИКд№ЃЌИпбѕЪЧШШЬЌбзжЂЬЌЪЧNF-kBЃЌГЃЬЌдђЪЧNrf2ИКд№ЃЌетЪЧБШНЯвтЭтЕФЁЃЃЉ
ЯИАћЖдбѕЛЏгІМЄЕФЗДгІ
ЯИАћЖдбѕЛЏЛЙдгІМЄЕФРэНтжївЊРДздбаОПбѕЛЏМСХЈЖШдіМгЖдЯИАћКЭЩњЮяЬхЕФЩњЛЏКЭЩњРэгАЯьЁЃШчЭМ1ЫљЪОЃЌЯИАћРћгУИїжжЕААзжЪЁЂУИКЭзЊТМЭјТчРДЯьгІбѕЛЏМСЕФдіМгБЉТЖЃЌетЛсЦЦЛЕЮШЬЌаХКХВЂПЩФмЕМжТВЛПЩФцЕФДѓЗжзгЫ№ЩЫЁЃЖдбѕЛЏЛЙдгІМЄЕФЯьгІгЩБЛбѕЛЏЮяаоЪЮЕФбѕЛЏЛЙдУєИаЕААзПижЦЃЌдкетжжаоЪЮаЮЪНЯТЫќУЧЕФЙІФмЗЂЩњБфЛЏ37ЁЃвЛзщжївЊЕФВЮгыЖдПЙбѕЛЏгІМЄЕФбѕЛЏЛЙдЕїПиЕААзЪЧзЊТМЕїНквђзгЃЈБэ1ЃЉЃЌЫќУЧв§ЗЂЖдЯИАћбѕЛЏЩњРэдіМгЕФЪЪгІадЯьгІ38,39ЁЃзюНќЕФвЛЯюбаОПМьВщСЫдкБЉТЖгкбѕЛЏМСЪБзЊТМвђзгМЄЛюЕФБфЛЏЪЧШчКЮдкЪБМфЩЯаЕїЕФ40ЃЌДњаЛКЭзЊТМзщЯьгІвРРЕгкЯИАћРраЭКЭбѕЛЏМСХЈЖШ41ЁЃвЛИіЬиБ№ФмБэеїЩњРэбѕЛЏМСЫЎЦНБфЛЏЕФДЋИаЦїЪЧЦеБщБэДяЕФКЫвђзгКьЯИАћЩњГЩЫи2ЯрЙивђзгЃЈNRF2ЃЉМАЦфвжжЦМСKelchбљECHЯрЙиЕААз1ЃЈKEAP1ЃЉ42ЁЃKEAP1ЪЧвЛИібѕЛЏМСКЭЧзЕчЪдМСДЋИаЦїЃЌЕБбѕЛЏМСЫЎЦНЕЭЪБЃЌЫќгыCullin 3ЃЈCul3ЃЉЛљЗКЫиСЌНгУИНсКЯЃЌНЋNRF2БъМЧЮЊЕААзУИЬхНЕНт43ЁЃбѕЛЏМСдіМгДЅЗЂKEAP1жаЬиЖЈАыызАБЫсЕФбѕЛЏЃЌЕМжТNRF2ЪЭЗХВЂРЉЩЂЕНКЫФкЃЌДгЖјБэДявЛЯЕСаПЙбѕЛЏЛљвђЃЌетаЉЛљвђЪЧGSHЩњЮяКЯГЩКЭЯћГ§H2O2ЫљБиашЕФЃЈР§ШчЃЌGSHЙ§бѕЛЏЮяУИЃЉ44ЁЃетжжЛљгкNRF2ЕФЛљвђБэДяЩЯЕїЙЙГЩСЫЖОЮяаЫЗмаЇгІМССП-ЗДгІЛњжЦЕФЛљДЁ45ЃЈЭМ2ЃЉЁЃзїЮЊЕЏадЕФЛљБОЛњжЦЃЌЖОЮяаЫЗмаЇгІЕФвЛАуддђ46ОпгаЙуЗКЕФживЊадЃЌР§ШчдкЩёОБЃЛЄЗНУцЕФЮФЯзжагаМЧТМ47ЁЃNRF2ЛЙМЄЛюNRF2геЕМЕФЗжУкБэаЭЛљвђЃЌетЪЧвЛИіУтвпМрЪгЯЕЭГЃЌЫќПЩвдЧхГ§вбЫ№ЛЕЕНЮоЗЈаоИДЕФЯИАћ48ЁЃЦфЫћбѕЛЏЛЙдЕїПиЕФзЊТМвђзгАќРЈШБбѕгеЕМвђзг1ЃЈHIF-1ІСЃЉКЭШШанПЫвђзг1ЃЈHSF-1ЃЉЁЃШБбѕКЭШШанПЫЬѕМўЖМЛсЕМжТЯпСЃЬхдкКЫжмЮЇЛ§ОлЃЌВЂдкКЫФкв§Ц№ROSВњЩњЕФдіМг49,50ЁЃЫфШЛHIF-1ІСЫЦКѕЭЈЙ§бѕЛЏМСНщЕМЕФИЌАБѕЃєЧЛЏУИЪЇЛюЖјЮШЖЈЃЌВЂгеЕМаэЖрВЮгыЬЧНЭНтКЭGSHЩњЮяКЯГЩЕФЛљвђЃЌЕЋHSF-1ЛюадЖдбѕЛЏМСНщЕМЕФАыызАБЫсДЮЛЧѕЃЛЏУєИаЃЌВЂгеЕМЗжзгАщТТКЭЦфЫћЕААзжЪЮШЬЌЭјТчзщЗжЕФБэДя51ЁЃ
Бэ1ЃКЪмбѕЛЏЛЙдПижЦЕФЯИАћЕїНкЕААз
Г§СЫжБНгЛђМфНгБЛбѕЛЏМСМЄЛюЭтЃЌМИИігыбѕЛЏЛЙдЯрЙиЕФзЊТМвђзгЛЙЪмЕНЖЬЃЈ<200 ntЃЌsncRNAЃЉЛђГЄЃЈ>200 ntЃЌlncRNAЃЉЗЧБрТыRNAЕФЕїНкЃЌетаЉRNAЕФБэДяЫЎЦНгЩH2O2геЕМЃЈВЮПМЮФЯз52ЃЉЁЃР§ШчЃЌЯёLINC00239етбљЕФlncRNAКЭmiRNA-200aДЅЗЂKEAP1 mRNAНЕНтЃЌДгЖјдіМгNRF2ЕФЮШЖЈад53ЁЃЗДвхlncRNA HIFALдіЧПСЫHIF-1НщЕМЕФЬЧНЭНтЛљвђЕФзЊТМ54ЃЌЖјlncRNA HEATЪЧHSF-1ЕФзЊТМЩВГЕЃЌгУгкдкВЛдйашвЊЪБШЁЯћШШанПЫЗДгІ55ЁЃетаЉЗЧБрТыRNAЙВЭЌзїЮЊвЛИіЕўМгЕФаХКХЃЌгыбѕЛЏМСвЛЦ№ЙЄзїЃЌЕїНкЙиМќЕФбѕЛЏЛЙдУєИазЊТМвђзгЃЌМЄЛюЯИАћгІМЄЗДгІЁЃ
ЯИАћЖдЛЙдгІМЄЕФЗДгІ
ЛЙдгІМЄвВПЩФмЖдЯИАћЙЙГЩЮЃЯеЃЌПЩФмЪЧЖдТ§адNRF2МЄЛюЕФЗДгІЁЃР§ШчЃЌЫќПЩвдЭЈЙ§діМгввШЉЭбЧтУИ3ЕФБэДяРДв§ЗЂЃЌетЪЧвЛжжВњЩњNADHЕФУИ56ЁЃЮЊСЫЕжЯћЛЙдгІМЄЃЌДцдкЬиЖЈЕФДЋИаЦїЕААзЃЌЕБЫќУЧБЛЛЙдЪБЃЌЛсМЄЛюВњЩњбѕЛЏМСЕФЭООЖ57ЃЈЭМ2ЃЉЁЃвЛИіетбљЕФДЋИаЦїЕААзЪЧгыFollicleЯрЛЅзїгУЕААз1ЃЈFNIP1ЃЉЃЌдкЦфбѕЛЏаЮЪНЯТЃЌЭЈЙ§вжжЦФћУЪЫсбЛЗДњаЛКЭбѕЛЏСзЫсЛЏзїгУЃЌзїЮЊЯпСЃЬхH2O2ЩњГЩЕФУХЮР57ЁЃетжжЕААзЕФЛЙддіМгСЫЕчзгЭЈЙ§ЕчзгДЋЕнСДЃЈETCЃЉЕФСїЖЏЃЌДгЖјдіМгСЫбѕЛЏМСЕФВњЩњЁЃДЫЭтЃЌЛЙдгІМЄПЩвдЭЈЙ§геЕМШщЫсЭбЧтУИРДЛКНт56ЁЃвђДЫЃЌЛЙдгІМЄИагІЪЧвЛжжжСЙиживЊЕФЗДгІЃЌашвЊгУРДЖдПЙТ§адЙ§ЖШДЬМЄЕФбѕЛЏМСЯьгІЯЕЭГЃЌШчNRF2ЁЃ
ЯИАћгІМЄЗДгІЕФзлКЯ
вЛИіЙиМќЕФбаОПСьгђЩцМАдкКЮЪБвдМАШчКЮдкбѕЛЏЛЙдгІМЄКѓМЄЛюИїжжЯИАћгІМЄЗДгІЕФЮЪЬт58ЁЃбѕЛЏгІМЄЗДгІгыЦфЫћРраЭЕФгІМЄЗДгІЯрЛЅСЊЯЕВЂОЋЯИЕїНкЃЌР§ШчзлКЯгІМЄЗДгІ59ЁЂЮДелЕўЕААзЗДгІЃЈUPRЃЉ60ЁЂШШанПЫЗДгІЃЈHSRЃЉ51ЁЂDNAЫ№ЩЫЗДгІЃЈDDRЃЉ61ЕШЁЃNRF2ЁЂNF-ІЪBЁЂHIF-1КЭHSF-1гызлКЯгІМЄЗДгІЁЂUPRЁЂHSRКЭDDRЯрЛЅСЊЯЕЃЌНЋбѕЛЏЛЙдИагІећКЯЕНетаЉгІМЄЗДгІжа62,63ЁЃдкДЫЃЌживЊЕФЪЧвЊжИГіЃЌМДNRF2ЁЂNF-ІЪBЁЂHIF-1ЕШжївЊЕФбѕЛЏЛЙдгІМЄЗДгІЯЕЭГЃЈБэ1КЭЭМ2ЃЉвдЕЭПедиЫйТЪдЫааЃЌЖјВЛЪЧзїЮЊПЊ/ЙиПЊЙиЃЌЕїећЛљвђБэДяЗДгІвдТњзуЯИАћЕФбѕЛЏЛЙдашЧѓЁЃетдЪаэЖдШЮКЮЭЛШЛдіМгЕФбѕЛЏМСзіГібИЫйЗДгІЁЃЖЈвхСйНчЕуЕФЪЧЪВУДЃЌМДДггавцгІМЄзЊБфЮЊгаКІгІМЄЕФЙ§ЖЩЃПД№АИПЩФмвђЧщОГКЭЯИАћЖјвьЁЃетПЩвддкЕЅЯИАћЫЎЦНЩЯНјааМрВтКЭНјвЛВНЬНЫї64ЁЃдкауРівўИЫЯпГцжаМЦЫубѕЛЏЛЙдгІМЄаХКХуажЕЃЈRSTЃЉЃЈЭМ2ЃЉвбдкЪЕбщЩЯНјааСЫГЂЪдЁЃОЁЙмБЉТЖгкГЌбѕвѕРызгЃЈO2•−ЃЉЗЂЩњЦїАйВнПндкЕЭХЈЖШЯТЛсдіМгЪйУќЃЌЕЋИќИпЫЎЦНЕФБЉТЖЕМжТЪйУќЫѕЖЬ65ЁЃетаЉЪЕбщЛЎЖЈСЫC. elegansгУРДМЄЛюбѕЛЏЛЙдЕїНкЭООЖЕФбѕЛЏМСХЈЖШЗЖЮЇЃЌвдМАЗЂЩњбѕЛЏгІМЄЕФуажЕЁЃЪЙгУЯИАћХрбјЃЌЗЂЯжЕЭМССПЕФH2O2ДЅЗЂUPRКЭHSRЃЌетЖдгкдкЧсЮЂШШдЄДІРэЯТЮЌГжЕААзжЪЮШЖЈадЪЧБиашЕФ66ЁЃЭЌбљЃЌЭЈЙ§ЧсЮЂЕФбѕЛЏгІМЄМЄЛюDDRЃЌЪЙЯИАћЖдУїЯдЕФгІМЄЬєеНгаЫљзМБИ67ЁЃ
Ш§ЁЂЯИАћФкКЭЯИАћМфЕФбѕЛЏЛЙдаХКХДЋЕМ
бѕЛЏЛЙдбаОПжаЕФвЛИіЛюдОЛАЬтЪЧЙигкШчКЮдкОжВПВњЩњЕФбѕЛЏМСКЭШЋОжЕФбѕЛЏЛЙдзДЬЌБфЛЏдкЯИАћФкВПКЭВЛЭЌЯИАћжЎМфНјааЭЈаХЕФЮЪЬтЁЃаэЖрЩњГЄвђзгЪмЬх68ЁЂећКЯЫи69ЁЂЯИАћвђзгКЭЦфЫћаХКХФЃЪНЃЈМћЁАЛЗОГЪЪгІЁБВПЗжЃЉжБНгдкжЪФЄДІЦєЖЏвЛИібѕЛЏЛЙдМЖСЊЃЌЕМжТЯИАћФкКЭВЛЭЌЯИАћЮЛжУжЎМфЕФбѕЛЏЛЙдЭЈаХ70ЁЃБОНкНщЩмСЫгаЙибѕЛЏЛЙдЭЈаХЕФЕБЧАжЊЪЖЁЃ
ЯИАћФкбѕЛЏЛЙдЭЈаХ
ИїжжбЧЯИАћЯИАћбЇПеМфИїгаЦфЬиЖЈЕФбѕЛЏЛЙдЮЂЛЗОГЃЌдкетаЉЮЂЛЗОГжаДцдкЯрЕБДѓЕФвьжЪадКЭВЈЖЏЁЃЮЊСЫЬжТлЯИАћФкбѕЛЏЛЙдЭЈаХЃЌЮвУЧЪзЯШНЋНщЩмИїИіЯИАћЮЛжУЕФЙиМќбѕЛЏЛЙдЬиеїЁЃ
ЯИАћжЪФЄ
ЯИАћжЪФЄжаЧЖШыЕФNADPHбѕЛЏУИЃЈNOXЃЉЭЈЙ§аХКХНщЕМЕФСзЫсЛЏзїгУМЄЛюЃЌВњЩњГЌбѕвѕРызгздгЩЛљЃЌЫцКѓдкАћЭтГЌбѕЛЏЮяЦчЛЏУИЃЈSOD3ЃЉДпЛЏЕФЗДгІжазЊЛЏЮЊH2O2ЃЈЭМ3ЃЉЁЃетжжNOX-SOD3АћЭтбѕЛЏЛЙдДЎСЊЖдгкбѕЛЏЛЙдаХКХДЋЕМжСЙиживЊЃЌР§ШчЃЌДгОоЪЩЯИАћЕНЦфЫћЯИАћРраЭЕФаХКХДЋЕн71ЁЃАћЭтЕФH2O2ЭЈЙ§зЈУХЕФЫЎЭЈЕРЕААзЃЈAQPsЃЉЁЂЙ§бѕЛЏЮяЭЈЕР72ЛђаЮГЩОпгабѕЛЏЛЙдЛюадЕФФкЬх73РЉЩЂНјШыЯИАћЃЌетаЉФкЬхБЛГЦЮЊЁАredoxosomesЁБ74ЃЌЮЊЫцКѓЕФЯИАћФкбѕЛЏЛЙдаХКХДЋЕМЦЬЦНСЫЕРТЗЁЃRedoxosomesДгЯИАћЪЭЗХЕНАћЭтПеМфЃЌЪЙЫќУЧФмЙЛДйНјЯИАћМфЕФбѕЛЏЛЙдаХКХзЊЕМ75ЁЃ
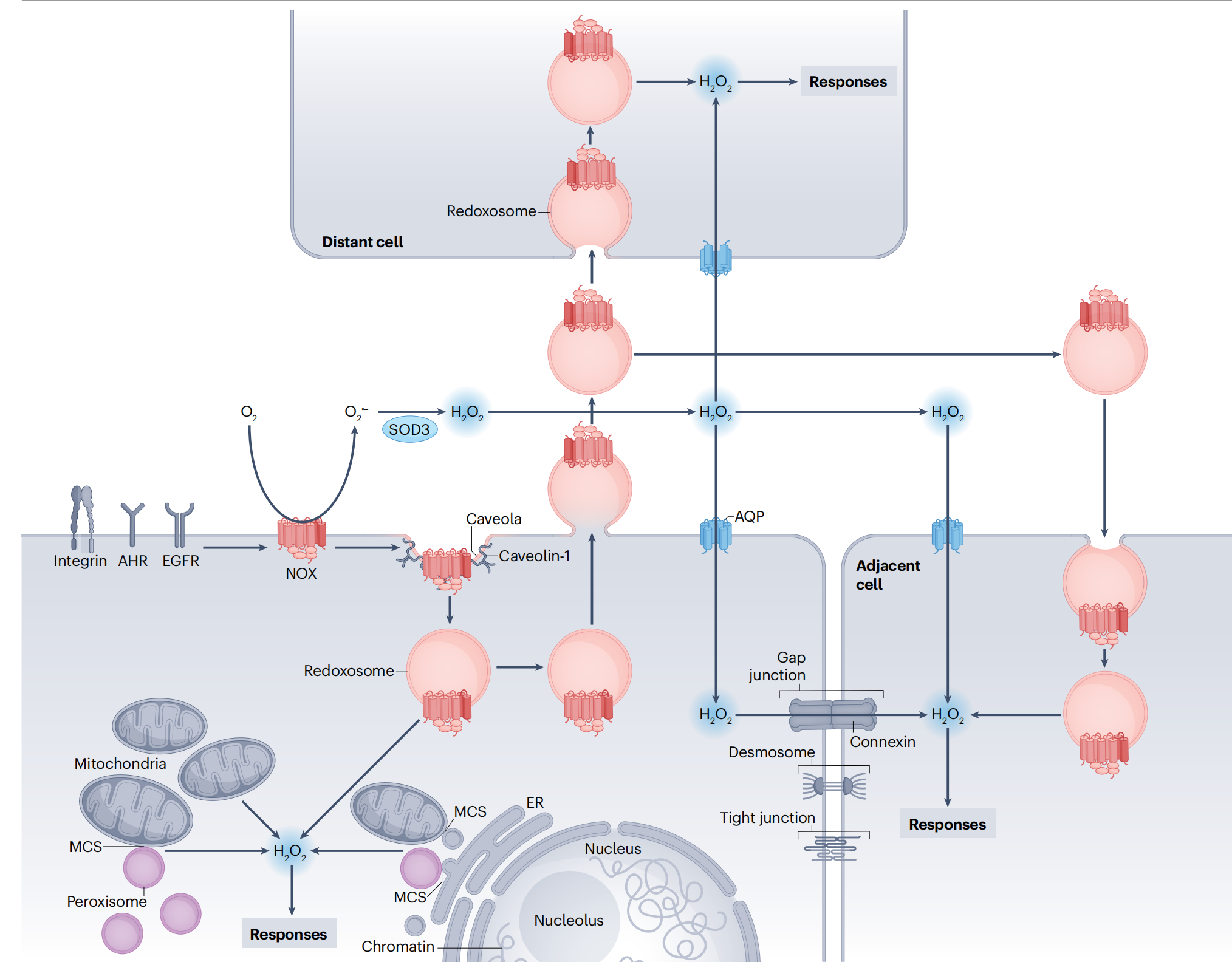
ЭМ3ЃКЯИАћФкЭтбѕЛЏЛЙдЭЈаХЕФИХРРЭМ
етИіМђЛЏЕФЗНАИОлНЙгкH2O2зїЮЊвЛжжжївЊЕФЯИАћаХКХбѕЛЏМСЁЃЖЈЮЛгкЯИАћжЪФЄЕФNADPHбѕЛЏУИЃЈNOXЃЉдкАћЭтВњЩњO2•−ЃЌШЛКѓБЛГЌбѕЛЏЮяЦчЛЏУИЃЈSOD3ЃЉзЊЛЏЮЊH2O2ЁЃH2O2НјШыЯИАћвЊУДЭЈЙ§зЈУХЕФЫЎЭЈЕРЕААзЃЈМДЙ§бѕЛЏЮяЭЈЕРЃЉЃЌвЊУДЭЈЙ§ФкЭЬзїгУаЮГЩОпгабѕЛЏЛЙдЛюадЕФФкЬх73ЃЌетаЉФкЬхБЛГЦЮЊЁЎredoxosomesЁЏ75ЃЌжївЊдкНбзДНсЙЙДІаЮГЩЁЃдкЯИАћФкЃЌШчЯпСЃЬхЁЂЙ§бѕЛЏЮяУИЬхКЭФкжЪЭјЃЈERЃЉЕШЯИАћЦївдМАДѓдМ40жжбѕЛЏУИВњЩњгУгкаХКХДЋЕнЕФH2O2ЃЈВЮМћВЮПМЮФЯз9ЃЉЁЃетШ§ИіЯИАћЦїЭЈЙ§ФЄНгДЅЕуЃЈMCSsЃЉДІЕФФЄУЊЖЈЖјНєУмНгДЅЁЃЯИАћМфЕФбѕЛЏЛЙдЭЈаХЭЈЙ§жБНгЯрСкЯИАћжЎМфЕФСЌНгзгНјааМфЯЖСЌНгЭЈаХЃЌСЌНгзггЩСНИіСЌНгЕААзАыЭЈЕРзщГЩЃЌЛђепЭЈЙ§ЫЎЭЈЕРЕААзЛђФкЭЬзїгУЕФАћЭтФвХнЃЈEVsЃЉЃЈЧАУцЬсЕНЕФredoxosomesЃЉЁЃЧХСЃКЭНєУмСЌНггажњгкНЋЯИАћБЫДЫСЌНгЦ№РДЃЌНЋНгЪебѕЛЏЛЙдаХКХЕФСкНќЯИАћДјЕНВњЩњH2O2аХЪЙЕФЯИАћИННќЁЃЕААзжЪdesminОпгавЛИіАыызАБЫсЃЈCys333ЃЉЃЌЦфбѕЛЏЛсЕМжТЧХСЃЦЦЛЕ259ЁЃЖдгкдЖДІЕФЯИАћЃЌH2O2аХКХПЩвдЭЈЙ§ФкЭЬзїгУЕФredoxosomesЛђЭЈЙ§ЫЎЭЈЕРЕААзНјааЁЃжЕЕУзЂвтЕФЪЧЃЌЛЙДцдкЦфЫћбѕЛЏЛЙдЭЈаХФЃЪНЃЌР§ШчЃЌЪЭЗХбѕЛЏЛЙдЛюадУИНјааЯИАћМфЕФХдЗжУкаХКХДЋЕн141ЃЌвдМАДгвЛбѕЛЏЕЊКЯУИЪЭЗХвЛбѕЛЏЕЊЁЃredoxosomeКЭЦфЫћEVsдкЯИАћМфбѕЛЏЛЙдаХКХДЋЕнжаЕФШЋВПЧБСІгаД§ЬНЫїЁЃAHRЃЌЗМЬўЪмЬхЃЛEGFRЃЌБэЦЄЩњГЄвђзгЪмЬхЁЃ
ЯИАћЙЧМм
МЁЖЏЕААзЯИАћЙЧМмЪЧбѕЛЏЛЙдЕїНкЕФжївЊФПБъ76,77ЁЃдкІТ-МЁЖЏЕААзжаЃЌCys374ЕФбѕЛЏНЕЕЭСЫЫПзДЃЈF-ЃЉМЁЖЏЕААзОлКЯЕФЫйТЪЃЌВЂгеЕММЁЖЏЕААз-МЁЧђЕААзНтОлЃЌетгажњгкдкЯИАћЧЈвЦЙ§ГЬжаЯИАћЙЧМмЕФЪеЫѕЁЃжЕЕУзЂвтЕФЪЧЃЌЯИАћЙЧМмгыNRF2-KEAP1ПЙбѕЛЏЯЕЭГжЎМфДцдкЙІФмЙиЯЕЃКKEAP1ЭЈЙ§ЦфЫЋИЪАБЫсжиИДНсЙЙгђгыМЁЖЏЕААзЫПНсКЯЃЌетБЛЗЂЯжЪЧЦфвжжЦNRF2ЛюадЫљБиашЕФ78ЁЃзїЮЊЯИАћЙЧМмзщГЩВПЗжЕФЮЂЙмвВЪмЕНбѕЛЏаоЪЮ79ЁЃдкЯИАћЙЧМмЕФИїжжЫПЯЕЭГжаЃЌАќРЈНЧЕААзЁЂvimentinКЭlaminдкФкЕФжаМфЫПзщГЩЕФЯИАћЕААздкбЙСІШЭаджаОпгабѕЛЏЛЙдЯьгІзїгУ80ЁЃ
Й§бѕЛЏЮяУИЬх
Й§бѕЛЏЮяУИЬхдкбѕЛЏЛЙдЕїНкжаЕФзїгУЃЈдкЦфЫћЕиЗНзлЪі81ЃЉвбОДѓДѓРЉеЙ82,83ЁЃбаОПБэУїЃЌЙ§бѕЛЏЮяУИЬхВњЩњЕФH2O2АаЯђГЌЙ§400жжЕААзжЪЃЌетаЉЕААзжЪЮЛгкЙ§бѕЛЏЮяУИЬхФкЃЌВЂЧвживЊЕФЪЧЃЌвВдкЯИАћжЪКЭЯпСЃЬхжаЕФЯИАћЦїЭтЁЃЙ§бѕЛЏЮяУИЬхжаВњЩњЕФH2O2ЭЈЙ§ЕїНкЦфЛЧѕЃЛЏТжРЊгАЯьАаЕААзЃЌетгажњгкЯИАћЦїМфЕФбѕЛЏЛЙдаХКХДЋЕн84ЁЃДЫЭтЃЌЙ§бѕЛЏЮяУИЬхЭЈЙ§вжжЦЙ§бѕЛЏЧтУИНјШыЙ§бѕЛЏЮяУИЬхРДЖдПЙбѕЛЏЬєеН85ЃЌДгЖјЭЈЙ§діЧПЙ§бѕЛЏЮяУИЬхЭтЙ§бѕЛЏЧтУИЕФФмСІ86ЃЌдіМгЯИАћЖдбѕЛЏгІМЄЕФЕжПЙСІЁЃЙ§бѕЛЏЮяУИЬхЕФбѕЛЏЛЙдаХКХЭЈЙ§МИжжЛњжЦЕїНкЯИАћУтвпДњаЛЃЌАќРЈЭЈЙ§NF-ІЪBМЄЛю87ЁЃ
ФкжЪЭј
ФкжЪЭјБЛШЯЮЊЪЧФЄдЫЪфКЭЯИАћЦїЙІФмМАЖЏЬЌЕФжїЕМЕїНкЦїЃЈЫцКѓЬжТлЃЉ88ЁЃдкЦфжкЖрЙІФмжаЃЌФкжЪЭјдкбѕЛЏЕААзелЕўвдЙЉЗжУкЗНУцЦ№зХжївЊзїгУЁЃЖўСђМќЕФаЮГЩЪЧгЩЕААзжЪЖўСђМќвьЙЙУИКЭФкжЪЭјбѕЛЏЛЙдУИ-1ЕФСЊКЯзїгУЭъГЩЕФЃЌУПИіЖўСђМќВњЩњвЛИіH2O2 89ЁЃЮЌГжФкжЪЭјжаЕФбѕЛЏЛЙдЦНКтЖдгкЕжЯћЕААзжЪЖОадгІМЄМАЦфКѓЙћКмживЊ90ЁЃ
ЯпСЃЬх
ЯпСЃЬхЪЧбѕЛЏЛЙдЕїНкЕФЛљЪЏ91,92ЃЌЦфЗЖЮЇДгсеЖЏСІбЇЕНгыЖржжбѕЛЏЛЙдЯрЙиЙІФмЕФЯИАћЦїМфНЛСї93ЃЌетаЉЙІФмГЌдНСЫЩњЮяФмСПбЇ94,95,96ЁЃЯпСЃЬхШчКЮВњЩњбѕЛЏЮяГЄЦквдРДвЛжБЪЧвЛИіЛюдОЕФбаОПСьгђ97ЃЌЕБЧАЕФНЙЕудкгкФкжЪЭјКЭЯпСЃЬхжЎМфдкГЦЮЊФкжЪЭј-ЯпСЃЬхЮЂгђЕФФЄНгДЅЕуЩЯЕФH2O2ЭЈСПЖЏЬЌ98,99ЁЃЙигкЯпСЃЬхЬиБ№ЪЧЕчзгДЋЕнСДдкбѕЛЏЛЙдЕїНкжаЕФзїгУЃЌвбгаДѓСПЮФЯзБЈЕР100ЁЃСНИіЕфаЭЕФЪмбѕЛЏЛЙдЕїНкЕФЯпСЃЬхЙ§ГЬЩцМАВњШШ101КЭжчвЙНкТЩ102ЃЌетРяВЛдйЯъЯИеЙПЊЁЃ
ЯИАћКЫ
ОЁЙмЯИАћКЫжЛЪЧбѕЛЏЮяЩњГЩЕФДЮвЊГЁЫљЃЌЕЋЫќдкЯИАћЖрИіЗНУцЕФбѕЛЏЛЙдЕїНкжаЗЂЛгзїгУЁЃЮЛгкЯИАћКЫжаЕФСђбѕЛЙЕААзМвзхГЩдБКЫЛЦЫибѕЛЏЛЙдУИ103ЪЧЯИАћбѕЛЏЛЙдПижЦЕФЙиМќЕїНкЦїЁЃЫќдкЯИАћКЫКЭЯИАћжЪжЎМфДЉЫѓЃЌВЂвдбѕЛЏЛЙдУєИаЕФЗНЪНгыМИжжЕААзжЪЯрЛЅзїгУ104ЁЃDNAЖдбѕЛЏЛЙдЕїНкЕФНјвЛВНМфНгЙБЯзЪЧDNAжабѕЛЏВњЩњЕФМюЛљаоЪЮЁЃЬиБ№ЪЧЃЌгыаоИДЬЧмеУИOGG1НсКЯЕФ7,8-ЖўЧт-8-бѕДњФёрбпЪЃЈ8-oxoGЃЉзїЮЊБэЙлвХДЋЕїПиБъМЧ105ЁЃOGG1НсКЯ8-oxoGВЂДйНјМЄЛюЕААзеаФМ106ЁЃ8-oxoGдкбѕЛЏЛЙдЕїНкжаЕФЩњЮябЇвтвхЛЙРЉеЙЕНБэзЊТМаоЪЮЃЌР§ШчRNAЕФЙВМлаоЪЮ107ЁЃШОЩЋжЪжиЫмвђзгШчРЕАБЫсЬивьадзщЕААзШЅМзЛљУИ1AЃЌгыЦфЫћШОЩЋжЪЕїНкЕААзШчзщЕААзЭбввѕЃУИНсКЯЃЌвВБЛШЯЮЊдкDNAаоИДЕААзКЭзщЕААзШЅМзЛљЛЏЕФНќЖЫбѕЛЏЛЙдЕїНкжаВњЩњH2O2 108ЁЃ
ШчЧАЫљЪіЃЌЗЧБрТыRNAЭЈЙ§гАЯьбѕЛЏЛЙдПижЦЛњжЦЬсЙЉВювьЛљвђБэДяЕФИДдгВуДЮ109ЁЃОЁЙмбѕЛЏгІМЄИФБфmRNAs52КЭlncRNAs110ЕФБэДяЃЌmiRNAs111КЭЛЗзДRNAs112ИФБфжДаабѕЛЏЛЙдЕїНкЕФУИЕФБэДяЁЃР§ШчЃЌmiR-23ЭЈЙ§ЕїНкСђбѕЛЙЕААзЛЙдУИ-1ЕФБэДядкЙЧїРМЁЗжЛЏжаЗЂЛгЙІФмзїгУЃЈВЮПМЮФЯз113ЃЉЁЃp53ЭЈТЗдкncRNAЖдбѕЛЏгІМЄЕФЗДгІжаЦ№зХКЫаФзїгУ114ЁЃ
ЯИАћЦїМфЕФбѕЛЏЛЙдЯрЛЅзїгУ
бЧЯИАћЦїжЎМфЕФбѕЛЏЛЙдаХКХЭЈТЗГЩЮЊПижЦЕФжївЊЪжЖЮЁЃвЛЯЕСааТЕФЯИАћЦїНгДЅСьгђЕФВћУївдМАЮЂгђКЭФЩУзгђЕФНсЙЙКЭЙІФмМјЖЈ115,116вдМАЗжзггЕМЗ117ЕФНщЩмЃЌЮЊЗжЮів§ШыСЫаТЕФЮЌЖШЁЃбЧЯИАћбѕЛЏЛЙдзщжЏЕФвЛАуВМОжЃЈМћВЮПМЮФЯз3,9ЃЉвбОЭЈЙ§дкЭъећЯИАћКЭЦїЙйФкЕФдЮЛЦїЙйбаОПДяЕНСЫаТЕФЫЎЦНЃЌетгыШБЗІжмЮЇЯИАћЮЂЛЗОГЕФЙТСЂЦїЙйжЦБИЕФбаОПЯрЖдЁЃ
ФкжЪЭј-ЯпСЃЬх-Й§бѕЛЏЮяУИЬхНгДЅБЛГЦЮЊЁАбѕЛЏЛЙдШ§НЧЁБЃЌетШ§ИіЯИАћЦїЭЈЙ§ФЄНгДЅЕуБЫДЫППНќЁЃЫќУЧЭЈЙ§ЫЎЭЈЕРЕААзКЭЮДЪЖБ№ЭЈЕРНЋбѕЛЏЮяВњЩњВЂдЫЪфЕНЯИАћжЪжа118ЁЃЭЈЙ§НЋЮяРэЩЯНгНќЕФжЪФЄзїЮЊбѕЛЏЛЙдЕїНкЕФживЊЙБЯзепАќРЈдкФкЃЌЩѕжСПЩвдГЦжЎЮЊЁАбѕЛЏЛЙдЫФБпаЮЁБЁЃДЫЭтЃЌетаЉЯИАћЦїЛЙгыШмУИЬхКЭжЌЕЮаЮГЩбѕЛЏЛЙдЯрЛЅзїгУ119ЁЃФкжЪЭјКЭЯпСЃЬхжЎМфЕФЯпСЃЬхЯрЙиФЄдкжЌжЪКЭCa2+зЊвЦжаЦ№зїгУЃЌВЂЕїНкетаЉЯИАћЦїжЎМфЕФбѕЛЏЛЙдаХКХ120ЁЃЙ§бѕЛЏПзЕААзAQP11дЪаэH2O2ИпаЇЕиПчФкжЪЭјФЄдЫЪф121ЃЌВЂДйНјЯпСЃЬхH2O2ЯђФкжЪЭјЕФзЊвЦ122ЁЃжЪФЄЫЎЭЈЕРЕААздкФкжЪЭј-жЪФЄНгДЅЕугыФкжЪЭјЯрЛЅзїгУ123ЁЃВЛЭЌзщжЏЛЙБэДяЖРЬиЕФAQPЭЌЙЄаЭЃЌетПЩФмгАЯьH2O2ЭЈЙ§етаЉФкжЪЭј-жЪФЄНгДЅЕуЕФдЫЪф124ЁЃЯпСЃЬх-жЪФЄНгДЅгУгкгааЇЕиНЛЛЛДњаЛЮяКЭРызгЃЌВЂЩњГЩИЛКЌИїжжЕААзжЪЕФаХКХЦНЬЈЁЃЯпСЃЬхЩѕжСПЩвдЭЈЙ§ЫэЕРФЩУзЙмдкЯИАћжЎМфзЊвЦ125ЃЌетПЩФмДйНјH2O2ЕФНЛЛЛвдДЋВЅбѕЛЏЛЙдаХКХЁЃЯИАћМфЕФбѕЛЏЛЙдЭЈаХЪЧЦїЙйКЭЯЕЭГЩњРэбЇЕФвЛИіЛљБОЬиеїЃЈМћЭМ3ЃЉЁЃетжжЭЈаХЭЈЙ§МИжжЗНЪНЗЂЩњЃЌАќРЈжБНгКЭМфНгЕФДгЯИАћЕНЯИАћЕФРЉЩЂЁЂЭЈЙ§СЌНгзгЕФМфЯЖСЌНгЭЈаХвдМАЭтУкЬхЁЃЯИАћМфбѕЛЏЛЙдДњаЛЮяРЉЩЂЕФЕфаЭР§згЪЧЭЈЙ§ШщЫс-БћЭЊЫсбѕЛЏЛЙдЖддкЯИАћКЭЦїЙйжЎМфЦНКтЯИАћжЪNADH/NAD+ЕФбѕЛЏЛЙдЕчЮЛ127ЃЌетЪЧвЛИіНЯДѓЕФбаОПСьгђЃЌВЮПМЮФЯз128жагазлЪіЁЃМфЯЖСЌНгЭЈаХЭЈЙ§СЌНгзгЕААзЗЂЩњЃЌетаЉПчФЄЕААздкЯрСкЯИАћЕФФЄжааЮГЩЭЈЕРЃЌГЦЮЊСЌНгзг129ЁЃЕБетаЉЭЈЕРДђПЊЪБЃЌЫќУЧдЪаэаЁЗжзгЃЈИпДядМ1000 DaЃЉПьЫйдкЯИАћМфНЛЛЛЃЌДгЖјСЌНгСНИіЯрСкЯИАћЕФЯИАћжЪЁЃдкСЌНгзгЕААзКЭбѕЛЏЛЙдаХКХжЎМфЕФЯрЛЅзїгУжаЃЌЭЈЕРЕФПЊБезДЬЌЭЈЙ§ЗвыКѓаоЪЮЪмЕНбѕЛЏЛЙдЕїНк130ЁЃ
ЭЈЙ§ЭтУкЬхЕФЯИАћЖдЯИАћЭЈаХЪЧЖЬОрРыКЭГЄОрРыЯрЛЅзїгУЕФСэвЛжжаЮЪН131ЁЃЭтУкЬхЃЌАќРЈЭтРДЬхКЭАћЭтФвХнЃЈЮЂФвХнЃЉЃЌдкжЪФЄЛђФкШмУИЬхЯЕЭГжаВњЩњЁЃЫќУЧБЛШЯЮЊЪЧЯИАћМфЭЈаХЕФЙиМќзщГЩВПЗжЃЌаЏДјВЂдкЬиЖЈЩЯЯТЮФЕФЕїНкЙ§ГЬжаДЋЕнбЁдёадЛѕЮя132ЁЃРэНтЭтУкЬхдкЯИАћМфбѕЛЏЛЙдЭЈаХжаЕФЙІФме§дкбИЫйЗЂеЙ133ЁЃЕБИїжжЯИАћБэУцЪмЬхБЛХфЬхНсКЯМЄЛюЪБЃЌЛсаЮГЩзЈУХЕФбѕЛЏЛЙдЛюадФкЬхЃЌГЦЮЊredoxosomesЃЈМћЭМ3ЃЉЁЃетаЉжЎЧАЬсЕНЙ§ЕФredoxosomes74вВПЩФмБЛДІРэГЩДгЯИАћЪЭЗХГіРДЕФЭтРДЬхЃЌаЏДјзХбѕЛЏЛЙдЛюадГЩЗж134КЭЫЎЭЈЕРЕААз135ЁЃвЛИіЯИАћМфбѕЛЏЛЙдаХКХДЋЕнЕФР§жЄЪЧдкОоЪЩЯИАћРДдДЕФбѕЛЏЛЙдЛюадЭтУкЬхзЊвЦЕНЩёОФкЬхКѓЪмЫ№жсЭЛЕФдйЩњ134ЁЃдкетжжЧщПіЯТЃЌКЌгаNOX2ЕФЭтРДЬхгыЪмЫ№жсЭЛШкКЯЁЃШЛКѓNOX2ВњЩњбѕЛЏЮяЃЌЪЙСзЫсУИPTENЪЇЛюЃЌбгГЄPI3K-AKTЭЈТЗдкЪмЫ№ЩёОдЊжсЭЛдйЩњжаЕФгеЕМ134ЁЃ
ЯИАћжмЮЇЕФбѕЛЏЛЙдЮЂЛЗОГБэЯжГіЯрЕБДѓЕФЖрбљадЁЃзїЮЊЖдгІМЄЗДгІЕФЯИАћМфбѕЛЏЛЙдаХКХДЋВЅвбБЛаЮЯѓЛЏЮЊЁАROSВЈЁБ16,136ЁЃЯрСкЯИАћжЎМфЕФЭЈаХПЩвдв§ЗЂЪЪгІадЗДгІЃЌвддіЧПЖдЛЗОГбЙСІЕФЕжПЙСІЁЃдкВИШщЖЏЮяЯИАћжаЃЌУцЯђЯИАћЭтПеМфЕФжЪФЄЭтвЖгыЯИАћЭтЛљжЪНгДЅ137ЁЃЯИАћЭтЛљжЪЕФзщжЏЬивьадЯИАћЕААзОлЬЧТжРЊгажњгкЕїНкбѕЛЏЛЙдаХКХЭЈТЗЃЌВЂЧвЬЧЕФЙІФмБфЛЏгыбѕЛЏЛЙдзДЬЌжЎМфДцдкСЊЯЕЃЌГЦЮЊЬЧ-бѕЛЏЛЙдЯрЛЅзїгУ138,139ЁЃгЩгкЬЧЛљзЊвЦУИКЭЬЧмеУИЕФзїгУЃЌР§ШчГЌбѕЛЏЮяЦчЛЏУИЕШбѕЛЏЛЙдЛюадУИЕФаджЪПЩвдБЛаоЪЮЁЃбѕЛЏЛЙдБфЛЏгыЕААзжЪЬЧЛљЛЏЃЈжївЊЪЧO-GlcNAcylationЃЉжЎМфЕФЯрЛЅЙиЯЕЙЙГЩСЫгІМЄЗДгІаХКХЭЈТЗНЛВцЖдЛАЕФживЊзщГЩВПЗж140ЁЃЙигкЯИАћжмЮЇЪЭЗХСђбѕЛЙЕААзЁЂЙ§бѕЛЏЮяЛЙдУИЁЂЕААзжЪЖўСђМќвьЙЙУИКЭлЯЛљбѕЛЏУИЕФжЄОнБэУїЫќУЧдкЯИАћЭтбѕЛЏЛЙдаХКХДЋЕнжаЕФзїгУЃЈР§ШчЃЌдкЕїНкУтвпЯИАћЙІФмЗНУцЃЉ141,142ЁЃ
етаЉЙигкЯИАћжмЮЇЛЗОГЕФЙлВьЮЊЬхвКжаЯИАћЭтбѕЛЏЛЙдПижЦПЊБйСЫаТЪгНЧ143ЁЃВИШщЖЏЮяЕФЯИАћЭтПеМфгЩЃЈНЯДѓЕФЃЉМфжЪПеМфКЭбЊЙмДВзщГЩЁЃбЊНЌжаH2O2ЕФХЈЖШвбБЛБЈИцЮЊ1-5 µMЃЈВЮПМЮФЯз144ЃЉЃЌЖјЪЙгУБэУцдіЧПРТќЙтЦзММЪѕЬНВтЯИАћжЪФЄБэУцЕФЩњЮяДЋИаЦїНвЪОСЫВЛОљдШЗжВМЕФАпПщЃЌЦфH2O2ХЈЖШдк1 µMЕН12 µMжЎМфЃЌЖјЯИАћФкH2O2ХЈЖШЮЊ5 nMЃЈВЮПМЮФЯз145ЃЉЁЃ
ЫФЁЂбѕЛЏЛЙдЕїПиЕФЕААзжЪзщ
бѕЛЏЛЙдгХгІМЄвРРЕгкЖдЯИАћДЬМЄВњЩњЕФбѕЛЏМСКЭЛЙдМСЃЈМћЭМ1ЃЉЁЃЯИАћЙІФмЕФбѕЛЏЛЙдЕїНкжївЊЪЧЭЈЙ§ЧПДѓЕФбѕЛЏЛЙдЛюадУИвдМАИїжжЩњЮяЗжзгРДЪЕЯжЕФЃЌетаЉЩњЮяЗжзгАќРЈЗЧБрТыRNAЁЂDNAбмЩњЕФЗжзгЁЂжЌжЪКЭЬМЫЎЛЏКЯЮяЃЌвдМАЕЭЗжзгСПЛЏКЯЮяЃЌШчгЊбјЙЉгІЕФИЈвђзгКЭЮЂСПгЊбјЫиЃЌЛЙгаЮЌЩњЫиКЭЁАЩњЮявђзгЁБЃЈЩњг§ЗгЁЂРрКњТмВЗЫиЁЂПЙЛЕбЊЫсКЭЖрЗгРрЃЉЁЃгХгІМЄаХКХЭЈЙ§бѕЛЏЛЙдЕААзжЪзщДЋВЅЃЌетЪЧвЛИізмГЦЃЌжИЕФЪЧЕААзжЪзщжаОРњПЩФцЛђВЛПЩФцбѕЛЏЛЙдЗДгІЕФзщЗж146ЁЃбѕЛЏЛЙдЕААзжЪзщвРРЕгкАыызАБЫсЁЂМзСђАБЫсЁЂЮјДњАыызАБЫсКЭЮјДњМзСђАБЫсНјааПЩФцбѕЛЏЛЙдЗДгІЕФФмСІЁЃЦфЫћАБЛљЫсЃЌШчЩЋАБЫсЁЂРвАБЫсЁЂОЋАБЫсКЭЕААзжЪжїСДЃЌвВПЩвдБЛбѕЛЏ147ЁЃбѕЛЏЛЙдЕААзжЪзщИажЊЁЂДЋВЅВЂЗХДѓбѕЛЏЛЙдаХКХЁЃдкбѕЛЏЛЙдЕїНкжаЕФаХКХзЊЕМРћгУСЫЧАУцЬсЕНЕФаЁЗжзгЛюадЮяжЪЃЈROSЁЂRNSЁЂRSSЕШЃЉЃЌЫќУЧБЛзмНсЮЊЛюадЮяжЪЯрЛЅзїгУзщ148,149ЃЈМћЭМ1ЃЉЁЃдкВйзїЩЯЃЌдкбѕЛЏЛЙдбаОПжаЃЌШЋЧђадЕФЪѕгябѕЛЏЛЙдЕААзжЪзщЭЈГЃдкИќЯСвхЕФвтвхЩЯгУРДжИДњАыызАБЫсбѕЛЏЛЙдЕААзжЪзщ150ЁЃ
ЕААзжЪАыызАБЫсдкбѕЛЏЛЙдЕїНкжаЕФзїгУ
ЕААзжЪАыызАБЫсЕФбѕЛЏКѓЗвыаоЪЮЃЈBox 2ЃЉдкЯИАћЫЎЦНЁЂзщжЏЗЖЮЇКЭгаЛњЬхФкОпгаживЊЕФЕїНкзїгУ151ЁЃЫцзХФмЙЛЧјЗжИїжжлЯЛљаоЪЮЕФИДдгЖЈСПбѕЛЏЛЙдЕААзжЪзщбЇММЪѕЕФЗЂеЙ152ЃЌвдМАЭЈЙ§жЪЦзМьВтЕЭЗсЖШЕААзЕФИФНј153ЃЌЯждкПЩвддкЕЅЯИАћЫЎЦНбаОПОРњПЩФцАыызАБЫсаоЪЮЕФЕААзжЪЁЃдкФъЧсКЭФъРЯЕФаЁЪѓжаНјааСЫШЋУцЕФЯИАћЬивьадКЭзщжЏЬивьадЕААзжЪАыызАБЫсбѕЛЏЕФЬхФкЦРЙРЃЌЩњГЩСЫвЛИіУћЮЊOximouseЕФЪ§ОнМЏЃЌСаГіСЫ10ИізщжЏжадМ171,000ИіАыызАБЫсВаЛљЕФбѕЛЏзДЬЌ153ЁЃДѓдМ34,000ИіАыызАБЫсЮЛЕудкдМ9,400ИіЕААзжЪжаБЛЗЂЯжОРњПЩФцАыызАБЫсбѕЛЏЃЌетЧПЕїСЫбѕЛЏЛЙдаХКХдкВИШщЖЏЮяжаЪЧвЛИіЦеБщЯжЯѓЁЃетИіЛуБрПЩвдЭЈЙ§вЛИіНЛЛЅЪНЭјТчзЪдДЃЈ'Oximouse'ЃЉЗУЮЪЃЌЫќЪЙЕУбщжЄПижЦУПИізщжЏЖРЬиЩњРэКЭДњаЛЬиеїЕФВЛЭЌАыызАБЫсбѕЛЏЭјТчГЩЮЊПЩФмЁЃжЕЕУзЂвтЕФЪЧЃЌдкНтЪЭбѕЛЏЛЙдЕААзжЪзщЪ§ОнЪБгІИУНїЩїЃЌвђЮЊЕБЧАЗНЗЈЕФЗЧЗВУєИаадЃЌЫќУЧвВПЩвдМьВтЕНВЛЪЧбѕЛЏЛЙдаХКХФПБъЕФАыызАБЫсВаЛљЁЃвђДЫЃЌгІИУНјааКѓајбаОПЃЌвдЬНОПЪЙгУбѕЛЏЛЙдЕААзжЪзщбЇМјЖЈЕФКђбЁАыызАБЫсЕФЯИАћЙІФм154ЁЃЫцзХбѕЛЏЛЙдЕААзжЪзщбЇММЪѕБфЕУИќМгОЋЯИКЭгаеыЖдадЃЌЙигкПЩФцАыызАБЫсПЊЙиЕФЯИАћаЇгІЕФБІЙѓНјвЛВНМћНтНЋБфЕУПЩгУ155ЁЃ
ЛљгкЗДгІЕФгЋЙтЬНеыгУгкМьВтЕААзжЪАыызАБЫсбѕЛЏЃЈCysOxЃЉЪЙбаОПШЫдБФмЙЛзЗзйЕААзжЪS-бЧЛЧѕЃЛЏЮЛЕуЃЌетЪЧлЯЛљбѕЛЏЙ§ГЬЕФЕквЛВНЃЌВЂдкЛюЯИАћжаЪЖБ№ЕїНкадАыызАБЫс156ЁЃДЫЭтЃЌбѕЛЏЛЙдЕААзжЪзщбЇЗжЮіПЩвдгыЛЏбЇвХДЋЙЄОпНсКЯЃЌР§ШчD-АБЛљЫсбѕЛЏУИЃЌЫќдЪаэдкВЙГфD-АБЛљЫсКѓдквЛВПЗжЯИАћЛђзщжЏжаПижЦВњЩњH2O2 157ЁЃЭЈЙ§НЋD-АБЛљЫсбѕЛЏУИгыH2O2ИагІФЃПщШчHyPerНсКЯЃЌПЩвдЭЌЪБДЬМЄКЭЗжЮіЬхФкЕФH2O2КЯГЩ158ЁЃетжжЗНЗЈЮЊбѕЛЏЛЙдУєИаЕААзгыИФБфЕФДњаЛжЎМфЕФЯрЛЅзїгУЬсЙЉСЫЖРЬиЕФаТМћНт159ЃЌе§ШчвЛЯюЙигкH2O2ЖдаФдрЪеЫѕСІгАЯьЕФбаОПЫљЪО160ЁЃHyPerЬНеыЛЙБЛгУРДЭЈЙ§НЋЬНеыШкКЯЕНЕЅИіНЭФИЕААзЕФCФЉЖЫЃЌДДНЈПЩЩИбЁЕФНЭФИОњжъПтЃЌДгЖјдкНЭФИЯИАћжаЪЖБ№H2O2ЮЂгђ161ЁЃ
бѕЛЏЛЙдЕААзжЪзщЕФСЊКЯЗжЮіНвЪОСЫЃЌдкЯИОњЁЂЙћгЌЁЂШфГцЁЂаЁЪѓКЭШЫРрзщжЏжаЃЌбѕЛЏЛЙдУєИаЕААзМИКѕЧЖШыЕНЯИАћЕФУПвЛИіДњаЛЭООЖКЭЩњРэЙ§ГЬжа162,163ЁЃЕААзжЪАыызАБЫсЖдбѕЛЏЕФЬивьадКЭУєИаадЪЧгЩжмЮЇЮяРэЛЏбЇЮЂЛЗОГЕФОжВПОВЕчЕїНкЫљОіЖЈЕФЃЌВЂЧвЪмЕНСкНќДјЕчВрСДЃЈШчArgЛђLysЃЉЕФЧПСвгАЯь153ЁЃЬиБ№ЪЧаПжааФЕФАыызАБЫсХфЮЛН№ЪєжааФвВЪЧбѕЛЏадАыызАБЫсаоЪЮЕФФПБъ164,165ЁЃИљОнЫќУЧдкЯИАћжаЕФЬиЖЈЮЛжУЃЌЕААзжЪШчЕААзСзЫсУИПЩвдЗЂЯжДІгкВЛЭЌРраЭЕФбѕЛЏаоЪЮжа166ЁЃ
лЯЛљбѕЛЏгАЯьЕААзжЪЙІФмЕФЛњжЦЪЧЖржжЖрбљЕФЃЌЭЈГЃашвЊЯъЯИЕФАИР§ж№вЛЩњЛЏКЭНсЙЙЗжЮіЃЛдкБэ1жаИјГіСЫвЛаЉЭЛГіЕФР§згЁЃаэЖрЛюадЮЛЕуАыызАБЫсПЩвдГЩЮЊбѕЛЏаоЪЮЕФФПБъЃЌЕМжТЩњРэЯрЙиЕФЙІФмадбѕЛЏЛЙдПЊЙиЁЃвЛИіЕфаЭР§згЪЧЬЧШЉЫс-3-СзЫсЭбЧтУИЃЌЦфЛюадЮЛЕуАыызАБЫсПЩвдЭЈЙ§МИжжРраЭЕФбѕЛЏКѓЗвыаоЪЮБЛаоИФЃЌв§ЗЂЬиЖЈгкКѓЗвыаоЪЮЕФЯИАћЗДгІ167ЁЃЯьгІгкЯИАћбѕЛЏМСЕФЪЪЖШдіМгЃЌЬЧШЉЫс-3-СзЫсЭбЧтУИЕФбѕЛЏЪЇЛюНЋЦЯЬбЬЧДњаЛжиЖЈЯђЕНЮьЬЧСзЫсЭООЖЃЌетдіМгСЫNADPHЕФВњЩњЃЌвдЮЌГжЯИАћбѕЛЏЛЙдзДЬЌЁЃаэЖрЦфЫћУИЕФЛюадвВЭЈЙ§ПЩФцЕФАыызАБЫсбѕЛЏЖјНЕЕЭЃЌДгЖјЕМжТДњаЛЕФжиаТХфжУвдНјааЯИАћаХКХДЋЕМ168ЁЃ
ЮЛгкЗЧЛюадЮЛЕуЕФАыызАБЫсЕФлЯЛљбѕЛЏвВПЩФмЕМжТЙІФмадНсЙћЃЌЗЖЮЇДгЕААзжЪНЕНтКЭЕААзжЪжиаТЖЈЮЛЕНЛёЕУаТЕФЕААзжЪЙІФмЁЃетаЉЪЧгЩЕААзжЪЙЙЯѓБфЛЏДЅЗЂЕФЃЌетаЉБфЛЏДгаЁЕФИФБфжмЮЇЕФЧтМќЭјТч169ЕНПЩвдЦєЖЏОжВПеЙПЊЪТМўЕФжївЊНсЙЙжиХХ170ЃЌЙбОлзДЬЌЕФБфЛЏ171ЛђШмНтЖШ172ЁЃетаЉЙЙЯѓБфЛЏШЁОігкЪмгАЯьАыызАБЫслЯЛљЕФЗжзгФкЯрЛЅзїгУЭјТчЃЌВЂЭЈЙ§ЬиЖЈЕФлЯЛљбѕЛЏВњЮяЃЈШчS-ЙШызИЪыФЛЏЛђЖўСђМќаЮГЩЃЉЛђМђЕЅЕФЮШЖЈлЯЛљЖЊЪЇ173НщЕМЁЃР§ШчЃЌНЭФИATPase GET3НсКЯСЫетаЉЬиеїЕФвЛаЉЁЃдкСНИіЙиМќАыызАБЫсзюГѕбѕЛЏКѓЃЌGET3ОжВПеЙПЊЃЌЙбОлЛЏЃЌВЂЭЌЪБДгATPвРРЕадАаЯђвђзгЧаЛЛЕНгааЇЕФATPЖРСЂАщТТЃЌБЃЛЄЕААзжЪУтЪмЕААзжЪбѕЛЏЕФгаЖОаЇгІ173ЁЃ
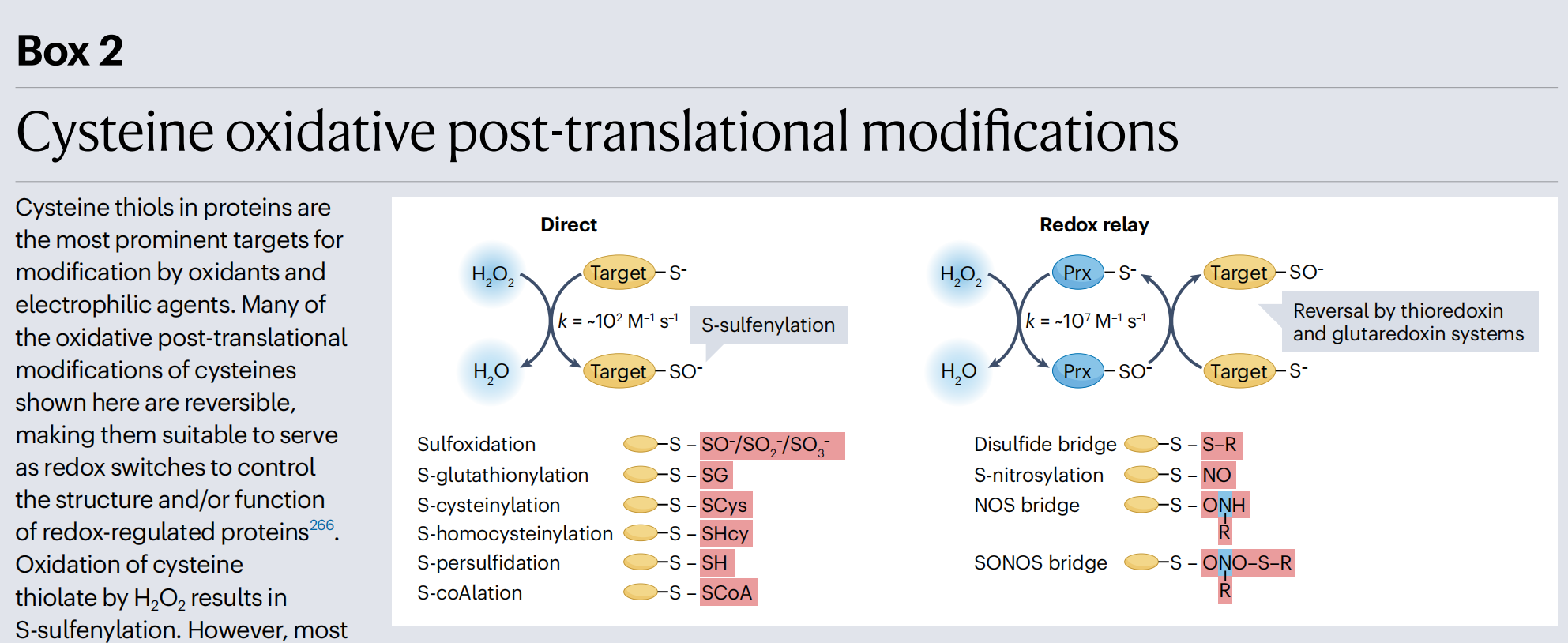
Box 2 АыызАБЫсбѕЛЏКѓЗвыаоЪЮ
ЕААзжЪжаЕФАыызАБЫслЯЛљЪЧбѕЛЏМСКЭЧзЕчЪдМСаоЪЮЕФзюжївЊФПБъЁЃетРяЯдЪОЕФаэЖрАыызАБЫсЕФбѕЛЏКѓЗвыаоЪЮЪЧПЩФцЕФЃЌЪЙЫќУЧЪЪКЯзїЮЊПижЦбѕЛЏЛЙдЕїНкЕААзжЪЕФНсЙЙКЭ/ЛђЙІФмЕФбѕЛЏЛЙдПЊЙи266ЁЃАыызАБЫслЯЛљЕФбѕЛЏЛсЕМжТS-СђѕЃЛЏЁЃШЛЖјЃЌДѓЖрЪ§ЕААзжЪлЯЛљгыH2O2ЕФЗДгІКмТ§ЃЌЯожЦСЫжБНглЯЛљбѕЛЏЕФгАЯьЁЃЯрБШжЎЯТЃЌЙ§бѕЛЏЮяЛЙдУИЃЈPrxЃЌгвЃЉКЌгаЦљНёЮЊжЙзюЛюдОЕФАыызАБЫсЃЌЫќУЧгыH2O2ЕФЖўМЖЗДгІГЃЪ§дМЮЊ107 mol−1 s−1ЃЌвђДЫБШДѓЖрЪ§ЦфЫћЛюадЕААзжЪАыызАБЫсИпЮхЕНСљИіЪ§СПМЖЃЈзѓЃЉ267ЁЃвђДЫЃЌЙ§бѕЛЏЮяЛЙдУИЪЧS-СђѕЃЛЏЕФжївЊФПБъЁЃШЛКѓЙ§бѕЛЏЮяЛЙдУИбѕЛЏАаЕААзжаЕФлЯЛљЃЌетБЛГЦЮЊбѕЛЏЛЙджаМЬЛњжЦ268ЁЃСђЫсѕЅЕФаЮГЩЪЙЙ§бѕЛЏЮяЛЙдУИЪЇЛюЃЌетПЩвдгЩGSHЙ§бѕЛЏЮяУИКЭСђбѕЛЙЕААзбИЫйФцзЊЁЃШЛЖјЃЌСђЫсѕЅвВПЩвдБЛH2O2НјвЛВНбѕЛЏЮЊбЧСђЫсѕЅ269ЁЃбЧСђЫсѕЅПЩвдБЛСђбѕЛЙЕААзФцзЊЁЃЭЈЙ§H2O2ЖдбЧСђЫсѕЅЕФНјвЛВНбѕЛЏВњЩњЛЧЫсѕЅЃЌетдкбѕЛЏгІМЄЦкМфЗЂЩњЁЃЛђепЃЌСђЫсѕЅПЩвдгыКЌлЯЛљЕФЛЏКЯЮяЗДгІЃЌаЮГЩгыGSHЃЈR-SSGЃЉЃЈS-ЙШызИЪыФЛЏЃЉ168ЁЂАыызАБЫсЃЈR-SS-CysЃЉЁЂЭЌаЭАыызАБЫсЃЈR-SS-HCysЃЉ270ЛђИЈУИAЃЈR-SSCoAЃЉ271ЕФЛьКЯЖўСђЛЏЮяЃЌЛђдкгыИННќЕААзжЪАыызАБЫсЗДгІЪБВЮгыЗжзгФкЛђЗжзгМфЖўСђМќЃЈR-SSRЃЉЁЃАыызАБЫслЯЛљгыRNSШчвЛбѕЛЏЕЊЃЈNOЃЉЗДгІЕМжТS-бЧЯѕЛљлЯЛљЃЈR-SNOЃЉ272ЃЌЖјЦфгыСђЛЏЧтЃЈH2SЃЉЕФЯрЛЅзїгУЕМжТЙ§СђЛЏЃЈR-SSHЃЉ270,273ЁЃЙ§СђЛЏЕФАыызАБЫсПЩвдЛЙдЛидЩњлЯЛљ224ЁЃЦфЫћбѕЛЏадАыызАБЫсаоЪЮАќРЈАыызАБЫс-РЕАБЫсЃЈNOSЃЉ274КЭАыызАБЫс-РЕАБЫс-АыызАБЫсЃЈSONOSЃЉ275бѕЛЏЛЙдЧХЃЌетаЉЪЧдкЕЊЃЈNЃЉЁЂбѕЃЈOЃЉКЭСђЃЈSЃЉжЎМфЗЂЩњЕФЕААзжЪНЛСЊЁЃетаЉбѕЛЏКѓЗвыаоЪЮЕФДѓЖрЪ§ЕФЛЙдЪЧгЩСђбѕЛЙЕААзКЭЙШбѕЛЙЕААзМвзхЕФГЩдБДпЛЏЕФЃЌЫќУЧЪЙгУNADPHзїЮЊЦфзюжеЕчзгЙЉЬхЁЃ
дкбѕЛЏЛЙдЕїПижаЃЌЕААзжЪЕФМзСђАБЫс (Met) ПЩвдБЛбѕЛЏГЩМзСђАБЫсбЧэП (MetO)ЃЌШЛКѓЭЈЙ§СЂЬхЬивьадЕФМзСђАБЫсбЧэПЛЙдУИ (Msrs) дйДЮЛЙдЮЊМзСђАБЫс174ЁЃМзСђАБЫсЕФбѕЛЏдкСђдзгЩЯВњЩњСЫвЛИіЪжаджааФЃЌаЮГЩСЫСНжжвьЙЙЬхЃЌR-MetO КЭ S-MetO174ЁЃЯрЖдгІЕиЃЌДцдкСНжжMsrЭЌЙЄУИРДЛЙдетаЉжааФВЂДйНјЕїПиЕФЬивьадЃКMsrAЃЈеыЖдR-MetOЃЉКЭMsrBЃЈеыЖдS-MetOЃЉ174,175ЁЃЕААзжЪМзСђАБЫсВаЛљЕФбѕЛЏПЩвдгЩMICALМвзхЕФЕЅМгбѕУИДпЛЏЃЌВЂгыMsrB1вЛЦ№ПижЦЃЌетвЛЙ§ГЬЪЧЭЈЙ§ЕЅЬхМЁЖЏЕААзЕФОлКЯзїгУРДЕїПиЕФ176ЁЃвбОМјЖЈГіЖрИіЙІФмадФПБъЃЌЫќУЧЦНКтСЫЕААзжЪжаЕФМзСђАБЫсКЭбѕЛЏаЭМзСђАБЫс(MetO)ЕФЫЎЦНЃЌЗЖЮЇДгИЦРызгЕїНкЁЂЗКЫиЛЏЯЕЭГЕНзЊТМвђзгЃЈР§ШчFOXOЃЉ174ЁЃ
дкбѕЛЏЛЙдЕїПиКЭБЃЛЄжаЃЌЕААзжЪЮјДњАыызАБЫсКЭЮјДњМзСђАБЫсвВЗЂЛгзїгУЁЃЮјДњАыызАБЫсЪЧЕк21ИіЕААзжЪАБЛљЫсЃЌАќКЌдк25жжШЫРрЕААзжЪжаЃЌЦфжааэЖрдкбѕЛЏЛЙдЕїПижаЦ№зїгУ177ЁЃЮјДњАыызАБЫсБШАыызАБЫсОпгаИќИпЕФЧзКЫадКЭИќЕЭЕФpKaжЕЃЌЕМжТЦфОпгаИќИпЕФбѕЛЏЛЙдЗДгІад177ЁЃСђбѕЛЙЕААзЛЙдУИКЭЙШызИЪыФЙ§бѕЛЏЮяУИЪЧЮјЕААззщжазюЭЛГіЕФГЩдБЁЃЫќУЧдкбѕЛЏЛЙдЕїПижаЕФзїгУвбБЛЙуЗКбаОПЃЈзюНќЕФзлЪіМћВЮПМЮФЯз178ЃЉЁЃЮјДњМзСђАБЫсдквЛаЉжВЮяКЭецОњжаКЯГЩЃЌЭЈЙ§ЪГЮяСДНјШыВИШщЖЏЮяЬхЯЕЃЌВЂжБНгЬцДњМзСђАБЫсВєШыЕААзжЪжаЁЃЫќЭЈЙ§здгЩЛљНщЕМЕФЛњжЦгыбѕЛЏМСЗДгІЃЌЦфЗДгІЫйТЪДѓдМБШМзСђАБЫсИп10БЖ179ЁЃЮјДњМзСђАБЫсбѕЛЏЮяПЩвдЭЈЙ§ЙШызИЪыФбИЫйгааЇЕиЛЙдЛиЮјДњМзСђАБЫсЃЌНЈСЂСЫЖдПЙЛюадбѕКЭЛюадЕЊЕФгааЇЗРЯп180ЁЃ
дкбѕЛЏЛЙдЕїПижаЃЌЕААзжЪРвАБЫсЕФбѕЛЏЕМжТаЮГЩЮШЖЈЕФЙВМлСЌНгЕФСкЮЛЖўРвАБЫсЛђ3,4-ЖўєЧЛљ-L-БНБћАБЫсЃЈL-dopaЃЉ181ЁЃзюНќдкДѓГІИЫОњжаЕФЕААзжЪзщбЇбаОПНвЪОСЫ70ЖржжВЛЭЌЕФЕААзжЪЃЌдкБЉТЖгкЭтдДЛђФкдДбЙСІЪБЖдЖўРвАБЫсЕФаЮГЩУєИа182ЁЃШЛЖјЃЌМјгкЩаЮДЃЈжСЩйФПЧАЃЉМјЖЈГіЛЙдЯЕЭГЃЌвђДЫРвАБЫсбѕЛЏПЩФмЪЧбѕЛЏЫ№ЩЫЕФНсЙћЃЌЖјВЛЪЧгУРДЕїПиЕААзжЪЛюадЕФбѕЛЏЛЙд182ЁЃЕААзжЪРвАБЫсБЛЙ§бѕбЧЯѕЫсбЮЯѕЛЏаЮГЩ3-ЯѕЛљРвАБЫсЃЌетЭЈЙ§ГЌбѕвѕРызггы•NOЗДгІаЮГЩЙ§бѕбЧЯѕЫсбЮЕФРЉЩЂПижЦЗДгІМфНггыбѕЛЏЛЙдЕїПиЯрЙи183ЁЃвђДЫЃЌЯѕЛЏЕААзжЪзщдкЯИАћаХКХДЋЕМжаЗЂЛгзїгУ10,184ЁЃ
ЮхЁЂЗжзгКЭЩњЮяЬхЕФбѕЛЏЛЙдвьжЪад
ЯИАћФкПЩвдВЩШЁВЛЭЌбѕЛЏЛЙдзДЬЌЕФЩњЮяЗжзгЕФЗжВМЪЧВЛОљдШЕФЃЌКЌгаПЩбѕЛЏАыызАБЫслЯЛљЕФЕААзжЪПЩвдвдЖржжВЛЭЌЕФаЮЪНДцдкЃЌГЦЮЊЁАбѕЛЏаЮЬЌЁБ185ЁЃР§ШчЃЌвЛИіКЌгаШ§ИіАыызАБЫсЕФЕААзжЪПЩвдВЩШЁАЫжжЙІФмЩЯВЛЭЌЕФбѕЛЏзДЬЌЃЈШчЙћжЛПМТЧСНИіАыызАБЫсЕФбѕЛЏЛЙдзДЬЌЃЉЃЌетЮЊЯИАћПеМфФкЕФЕЅИіЕААзжЪЗжзгЬсЙЉСЫЖРЬиЕФЁАбѕЛЏЛЙдЬиеїЁБЁЃШчЙћПМТЧЕНЯИАћВњЩњЖржжРраЭЕФбѕЛЏМСЃЌбѕЛЏаЮЬЌЕФвьжЪадПЩвдЭЈЙ§МИИіЪ§СПМЖНјвЛВНдіМг186ЁЃетжжбѕЛЏаЮЬЌФЃЪНЯджјдіЧПСЫЯИАћФкЕААзжЪЕФбѕЛЏЛЙдвьжЪадЁЃгЩгкбѕЛЏЛЙдЛюадЕААзжЪАыызАБЫсЕФаЮЪНдЖВЛжЙСНжжЃЈМћЭМЃЌBox 2ЃЉЃЌвђДЫдкИјЖЈЯИАћЬѕМўЯТЕААзжЪПЩФмЕФбѕЛЏаЮЬЌЪ§СПДѓДѓдіМгЁЃ
дкИіЬхЯИАћЫЎЦНЩЯвВДцдкзХбѕЛЏЛЙдвьжЪадЁЃЫцзХвХДЋБрТыЕФЬхФкбѕЛЏЛЙдДЋИаЦїЕААзЕФЗЂеЙЃЌР§ШчИагІH2O2ЕФHyPer187ЛђGSHбѕЛЏЛЙдзДЬЌДЋИаЦїGrx-roGFP188ЃЌМДЪЙЪЧИпЖШЭЌВНЕФПЫТЁЯИАћШКЬх189ЛђећИіЭЌЛљвђЩњЮяЬхШчауРівўИЫЯпГц190,191вВЯдЪОГіУїЯдЕФИіЬхМфВювьдкбѕЛЏЛЙдзДЬЌЩЯЁЃЖдОпгаВЛЭЌбѕЛЏЛЙдзДЬЌЕФПЫТЁЯИАћбЧШКЛђауРівўИЫЯпГцЕФЗжЮіНвЪОСЫдкећЬхШКЬхбаОПжаЮДБЛзЂвтЕНЕФбѕЛЏЛЙдУєИаЙ§ГЬЁЃетаЉбаОПБэУїЃЌЩњУќдчЦкЕФФкдкбѕЛЏЛЙдвьжЪадгажњгкИіЬхМфЕФБэЙлвХДЋОАЙлЁЂБэДяФЃЪНвдМАзюжеЩњЮяЬхЕФЪйУќЕФВювь192ЁЃ
ЯИАћЙ§ГЬЕФбѕЛЏЛЙдЕїПи
ЯИАћЕФбѕЛЏЛЙдЛЗОГЭЈЙ§бѕЛЏЛЙдЕїПиЕААзЖдДѓЖрЪ§ЯИАћЙ§ГЬНјааЕїПиЁЃвЛаЉЯджјЕФЙ§ГЬЪОР§ШчЭМ4ЫљЪОВЂдкДЫзмНсЁЃбѕЛЏЛЙдЛЗОГЮЂЕїДњаЛЭООЖВЂЕїНкДњаЛЮязщЃЌетгАЯьУИЕФЛюадЃЌР§ШчNAD+вРРЕадзщЕААзШЅввѕЃЛЏУИЃЈМДГСФЌаХЯЂЕїНкЕААзЃЉЃЌДгЖјИФБфБэЙлЛљвђзщ193,194ЁЃЯьгІбѕЛЏМСЫЎЦНБфЛЏЕФбѕЛЏЛЙдЕїПизщЕААзаоЪЮУИЛсгАЯьЛљвђБэДяЃЌДйНјПЩФмГжОУЩѕжСПчДњЕФМЧвфаЇгІ192,195ЁЃбѕЛЏМСПЩвдЩЯЕїзЊТМвђзгЩњЮяКЯГЩЛђИФБфЫќУЧЕФЮШЖЈадЃЌР§ШчжЎЧАЬжТлЙ§ЕФNRF2дкзЊТМЁЂmRNAЮШЖЈадКЭЗвыЫЎЦНЩЯЕФБфЛЏЁЃзЊТМвђзгЛюадЭЈЙ§бѕЛЏЛЙдЕїПиЕФМЄУИКЭСзЫсУИв§ЗЂЕФЗвыКѓаоЪЮЖјЗЂЩњИФБф39ЁЃМИКѕЕААзжЪКЯГЩЕФУПвЛИіЗНУцЃЌДгmRNAЮШЖЈадКЭЗвыЦ№ЪМЕНКЫЬЧЬхНсЙЙКЭзЊдЫRNAЖЏСІбЇЃЌЖМЩцМАбѕЛЏЛЙдУєИазщЗжЃЌИКд№ИљОнЯИАћЕФбѕЛЏЛЙдЬѕМўЕїећШЋОжЕААзжЪЗвыЫйТЪ3ЁЃЩњРэЫЎЦНЕФбѕЛЏМСЛЙЕїПиМЄУИЁЂСзЫсУИКЭзЊТМвђзгЕФЛюадЃЌвдгеЕМдіжГЁЂЗжЛЏКЭЧЈвЦ11,196,197,198ЁЃзюКѓЃЌЪмЕїПиЕФЯИАћЫРЭіЃЈRCDЃЉЃЌМДЯёЕђЭіетбљЕФЭООЖЃЌЩцМАбѕЛЏЛЙдУєИаЕААзРДПижЦЯИАћЩњДцЛЙЪЧЫРЭіЕФОіЖЈ199ЁЃОЁЙмЫќУЧЕФбѕЛЏЭЈГЃгеЕМЕђЭі200ЃЌЕЋдкжДааепАыызЬьЖЌУИЁЊЁЊзюжеИКд№ЕђЭіадЯИАћЫРЭіЕФЕААзУИЁЊЁЊЕФЛюадЮЛЕуАыызАБЫсЕФаоЪЮБЛгУРДБЃЛЄЯИАћУтЪмВЛЯЃЭћЕФВЛПЩФцЕђЭіадRCDГЬађЕФМЄЛю199ЃЈЭМ4fЃЉЁЃР§ШчЃЌдкОВЯЂЕФHeLaЯИАћжаЃЌЧААыызЬьЖЌУИ3КЭ9ЕФЗЂЩњГжајадЙ§СђЛЏЃЈвђДЫЪЇЛюЃЉзїгУЃЌВЂЭЈЙ§СђбѕЛЙЕААзНщЕМЕФШЅЙ§СђЛЏзїгУЕжЯћ201ЁЃбѕЛЏЛЙдаоЪЮЕФРраЭвдМАДйЕђЭіаХКХШчКЮМЄЛюбѕЛЏЛЙдЕААзНјЖјМЄЛюАыызЬьЖЌУИЫЦКѕШЁОігкЯИАћРраЭКЭОпЬхЧщОГ199ЁЃЬњЫРЭіЃЌвЛжжгЩFe2+НщЕМЕФВЛБЅКЭжЌЗОЫсЙ§бѕЛЏДЅЗЂЕФЗЧЕђЭіаЮЪНЕФRCD202ЃЌБЛGSHЙ§бѕЛЏЮяУИGPX4гааЇзшЖЯЃЌКѓепЪЙгУЮјДњАыызАБЫсЛљДпЛЏРДЛЙджЌжЪЙ§бѕЛЏЮя203ЁЃСђбѕЛЙЕААзЕФЩЅЪЇвВгажњгкЬњЫРЭіЃЌПЩФмЪЧЭЈЙ§бѕЛЏМСЛ§РлМАЦфЖдGSH-ЙШбѕЛЙЕААзЯЕЭГЕФгАЯь204ЁЃгУвЛжжаЁЗжзгferroptocideДІРэЃЌЫќФмЙВМлаоЪЮСђбѕЛЙЕААзЛюадЮЛЕуАыызАБЫсЃЌЗЧГЃгааЇЕив§ЗЂАЉЯИАћжаЕФЬњЫРЭі204ЁЃВЮгыRCDsЕФЦфЫћбѕЛЏЛЙдУєИаВЮгыепАќРЈp62ЃЌЩцМАжївЊгЩбѕЛЏМСНщЕМЕФDNAЫ№ЩЫЕїПиЕФRCDЭООЖparthanatos205ЃЌЩцМАoxeiptosisЕФбѕЛЏЛЙдУєИаKEAP1ЃЈВЮПМЮФЯз206ЃЉЃЌетЪЧвЛжжROSвРРЕадЁЂАыызЬьЖЌУИЖРСЂЕФRCDЃЌвдМАЩцМАбзадЯИАћЫРЭіГЬађНЙЭіЕФКЫмеЫсНсКЯгђбљЪмЬхЕААз3207ЁЃ

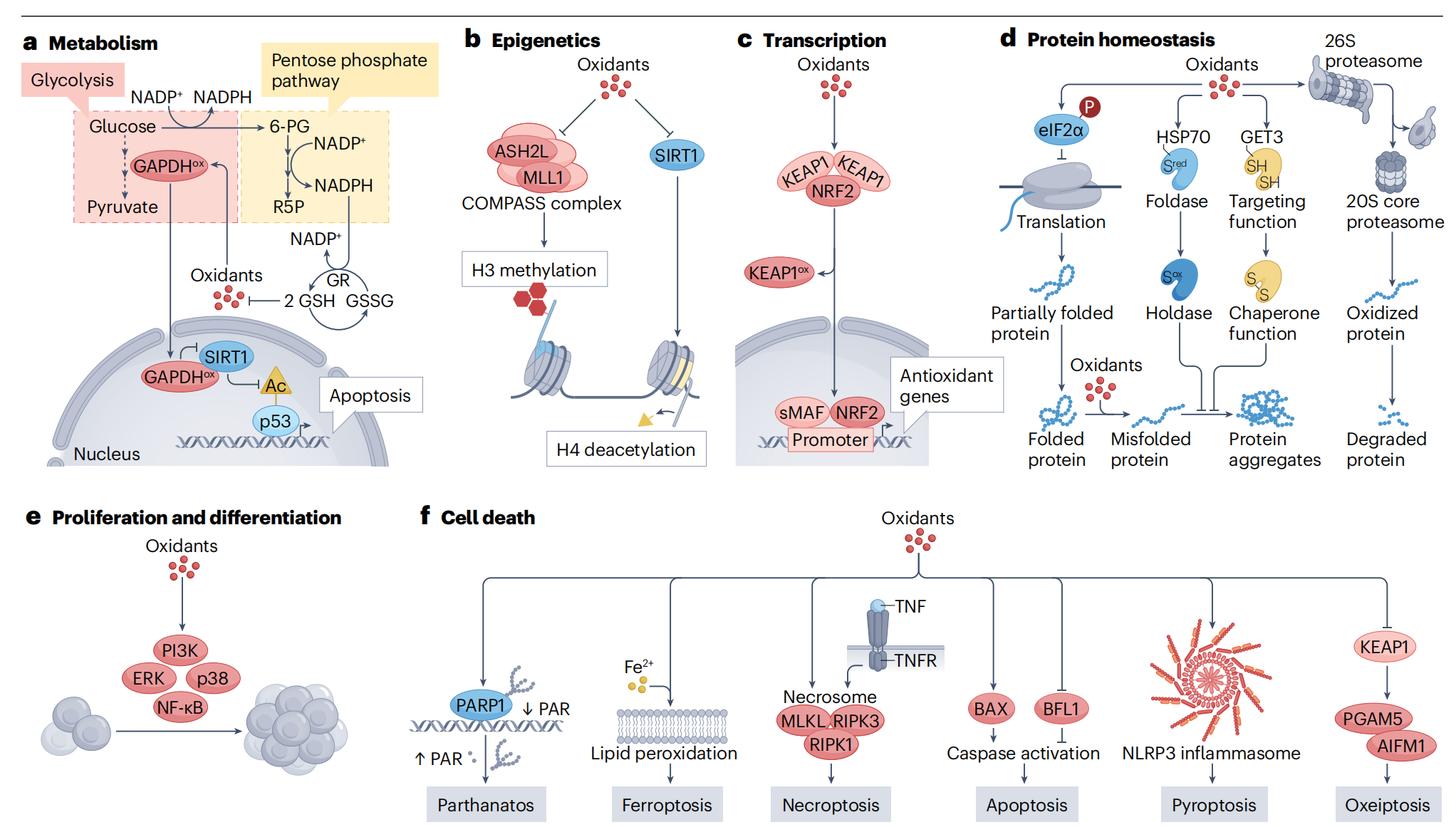
ЭМ4ЃКбѕЛЏЛЙдЕїПиЯТЕФжївЊЯИАћЙ§ГЬЁЃ
aЃЌДњаЛЃКзїЮЊбѕЛЏЛЙдПижЦДњаЛЭООЖЕФЕфаЭР§згЃЌЬЧНЭНтЕФКЫаФУИЁЊЁЊИЪгЭШЉ-3-СзЫсЭбЧтУИЃЈGAPDHЃЉдкЦфЛюадЮЛЕуКЌгавЛИібѕЛЏЛЙдУєИаЕФАыызАБЫсЁЃЫќЕФбѕЛЏаоЪЮЕМжТЭбЧтУИЙІФмЕФЫ№ЪЇЃЈGAPDHoxЃЉВЂзшЖЯЬЧНЭНтЁЃЦЯЬбЬЧБЛзЊЯђЮьЬЧСзЫсЭООЖвдаЮГЩNADPHЃЌетЪЧЛжИДЛЙдаЭЙШызИЪыФЃЈGSHЃЉГиЫљБиашЕФЁЃGAPDHoxЛЙПЩвдДЉЫѓЕНЯИАћКЫжаЃЌдкФЧРяЫќбѕЛЏаоЪЮВЂЪЇЛювРРЕгкNADЕФЕААзШЅввѕЃЛЏУИsirtuin-1ЃЈSIRT1ЃЉЁЃЮДФмШЅввѕЃЛЏp53ПЩФмЕМжТЯИАћЕђЭіЁЃbЃЌБэЙлвХДЋбЇЃКЯИАћФкЕФБэЙлвХДЋОАЙлгЩЦфжїЕМЕФбѕЛЏЛЙдЛЗОГЮЂЕїЁЃетРяеЙЪОСЫСНИібѕЛЏЛЙдУєИаЕФзщЕААзаоЪЮУИЃКзщЕААзРЕАБЫсN-МзЛљзЊвЦУИ2AЃЈMLL1ЃЌвВГЦЮЊKMT2AЃЉЃЌЪЧCOMPASSИДКЯЬхЕФГЩдБЃЌКЌгавЛИібѕЛЏЛЙдУєИаЕФаПжааФЃЌЦфбѕЛЏЕМжТШЋЧђH3K4me3ЫЎЦНЯТНЕЁЃSIRT1ЕФбѕЛЏЕМжТЦфШЅввѕЃЛЏУИЙІФмЕФЪЇЛюЃЌДгЖјЕМжТзщЕААзЩЯввѕЃЛЏРЕАБЫсЕФЛ§РлЁЃетгажњгкЛљвђБэДяЕФИФБфЁЃcЃЌЛљвђБэДяЃКбѕЛЏМСКЭЧзЕчзгЮяжЪжБНгМЄЛюЬиЖЈЕФзЊТМвђзгЃЌЕМжТЛљвђБэДяЕФБфЛЏЃЈБэ1ЃЉЁЃвЛИіЭЛГіЕФР§згЪЧРрЫЦKelchЕФECHЯрЙиЕААз1ЃЈKEAP1ЃЉЃЌЫќдкЗЧгІМЄЬѕМўЯТНсКЯКЫвђзгКьЯИАћ2ЯрЙивђзгЃЈNRF2ЃЉВЂНЋЦфАаЯђНЕНтЁЃKEAP1жабѕЛЏЛЙдУєИаАыызАБЫсЕФбѕЛЏЕМжТNRF2ЕФЪЭЗХЃЌЫцКѓNRF2жиаТЖЈЮЛЕНЯИАћКЫЃЌгыаЁMAFЃЈsMAFЃЉМвзхЕААзвьдДЖўОлЬхЛЏЃЌВЂМЄЛюБрТыПЙбѕЛЏЗРгљЯЕЭГЕААзжЪЕФЛљвђЁЃdЃЌЕААзжЪЗвыЁЂелЕўКЭНЕНтЃКетаЉЙ§ГЬЭЈЙ§бѕЛЏЛЙдЕїПиЕААзжБНгЛђМфНгПижЦЁЃбѕЛЏгІМЄЕМжТЭЈЙ§в§Ц№ЦєЖЏвђзгЃЈР§ШчeIF2ІСЃЉЕФСзЫсЛЏЁЂзЊдЫRNAЖЏЬЌКЭКЫЬЧЬхЙІФмЕФБфЛЏЃЌДгЖјШЋОжЯожЦЕААзжЪКЯГЩЁЃATPвРРЕаделЕўАщТТЃЈМДелЕўУИЃЉШШанПЫЕААз70ЃЈHSP70ЃЉЛђЕААзАаЯђвђзгGET3зїЮЊбѕЛЏгІМЄМЄЛюЕФАщТТЕФР§згЁЃЙиМќСђДМЕФбѕЛЏаоЪЮЕМжТЫќУЧзЊБфЮЊATPЖРСЂаЭБЃГжАщТТЃЈМДГжСєУИЃЉЃЌЫќУЧНсКЯЮДелЕўЕААзВЂЗРжЙЦфОлМЏЁЃетжжгІМЄЬивьадМЄЛюВЙГЅСЫбѕЛЏгІМЄНщЕМЕФЯИАћATPЫЎЦНЯТНЕКЭЫцжЎЖјРДЕФATPвРРЕадАщТТЕФЙІФмЯТЕїЁЃЕААзжЪЗКЫиЛЏКЭ26SЕААзУИЬхЙІФмЪмЕНбѕЛЏЛЙдНщЕМЕФБфЛЏвжжЦЃЌЖј20SЕААзУИЬхЛюаддіМгвдНЕНтбѕЛЏЫ№ЩЫЕФЕААзжЪЁЃeЃЌдіжГЁЂЗжЛЏКЭЧЈвЦЃКаэЖргеЕМдіжГКЭЗжЛЏЕФМЄУИКЭзЊТМвђзгЖМЪмЕНбѕЛЏЛЙдПижЦЁЃМЄЛюЪЧЭЈЙ§АыызАБЫсПЊЙиЕФжБНгбѕЛЏЛђЭЈЙ§ЩЯгЮЭООЖвжжЦМСЕФМфНгвжжЦЗЂЩњЕФЁЃСзжЌѕЃМЁДМ-3-МЄУИЃЈPI3KЃЉЛђЯИАћЭтаХКХЕїНкМЄУИЃЈERKЃЉЕФМЄЛюЭЈЙ§бѕЛЏЛЙдУєИаЕФЩЯгЮСзЫсУИШчСзЫсУИКЭеХСІЕААзЭЌдДЮяЃЈPTENЃЉКЭЕААзРвАБЫсСзЫсУИ1BЃЈPTP1BЃЌЮДЯдЪОЃЉЕФбѕЛЏЪЇЛюЖјбгГЄЁЃКЫвђзгІЪ-ЧсСДдіЧПзгЃЈNF-ІЪBЃЉЭЈЙ§АыызАБЫсбѕЛЏжБНгМЄЛюЃЌВЂЭЈЙ§NF-ІЪBвжжЦМСЕФвжжЦМфНгМЄЛюЁЃЭЌбљЃЌгІМЄМЄЛюЕФЫПСбдЛюЛЏЕААзМЄУИЃЈMAPKЃЉp38ЭЈЙ§бѕЛЏМСЕФДцдкжБНгКЭМфНгМЄЛюЁЃfЃЌЪмЕїПиЕФЯИАћЫРЭіЃЈRCDЃЉФЃЪНЃКparthanatosЪЧвЛИіжївЊгЩбѕЛЏМСНщЕМЕФDNAЫ№ЩЫЕїПиЕФRCDЭООЖЁЃЖрОл[ADP-КЫЬЧ]ОлКЯУИ1ЃЈPARP1ЃЉЕФЙ§ЖШМЄЛюВњЩњЙ§ЖрЕФЖрОлA-КЫЬЧЃЈPARЃЉЃЌЕМжТДгЯпСЃЬхЪЭЗХДйЕђЭівђзгЃЌЕМжТЯИАћЫРЭі205ЁЃЬњЫРЭіЪЧвЛжжЗЧЕђЭіаЮЪНЕФRCDЃЌгЩFe2+НщЕМЕФВЛБЅКЭжЌЗОЫсЙ§бѕЛЏДЅЗЂ202ЁЃЛЕЫРЪЧгЩжзСіЛЕЫРвђзгЃЈTNFЃЉНщЕМЕФЃЌЫќМЄЛювЛИіГЦЮЊЛЕЫРЬхЕФЖрЕААзИДКЯЬхЁЃЛЕЫРЬхЕФвЛИібѕЛЏЛЙдУєИаВЮгыепЪЧЪмЬхЯрЛЅзїгУЕФЫПАБЫс/ЫеАБЫсЕААзМЄУИ1ЃЈRIPK1ЃЉЃЌдкбѕЛЏКѓЃЌЫќОРњздСзЫсЛЏЃЌгыRIPK3КЭЛьКЯЦзЯЕМЄУИНсЙЙгђбљЕААзЃЈMLKLЃЉЯрЛЅзїгУЃЌВЂаЮГЩЛюадЛЕЫРЬх260ЁЃЕђЭіЪЧгЩ(pro)АыызЬьЖЌУИЕФЕААзЫЎНтМЖСЊЗДгІНщЕМЕФЃЌЫќУЧдкМЄЛюаЮЪНЯТНЕНтЯИАћГЩЗжВЂЕМжТЯИАћЫРЭіЁЃЕђЭіЕїНквђзгBAXЕФбѕЛЏМЄЛюЛђПЙЕђЭівђзгBFL1ЃЈвВГЦЮЊBCL2A1ЃЉЕФбѕЛЏЪЇЛюДЅЗЂЕђЭі200ЁЃНЙЭіЪЧвЛжжбзжЂадЯИАћЫРЭіаЮЪНЃЌЩцМАЖрЕААзИДКЯЬхбзадЬхЃЌЦфГЩдБАыызЬьЖЌУИ1МЄЛюПзИДКЯЬхgasdermin DЁЃКЌгаКЫмеЫсНсКЯгђЃЈNODЃЉбљЪмЬхЕААз3ЃЈNLRP3ЃЉзїЮЊЦфДЋИаЦїФЃПщЕФбзадЬхЫЦКѕЖдбѕЛЏМСЬиБ№УєИа207ЁЃOxeiptosisЪЧвЛИіжБНгЪмбѕЛЏЛЙдУєИаВЮгыепKEAP1ПижЦЕФRCDЭООЖ206ЁЃKEAP1бѕЛЏКѓЃЌЯпСЃЬхЫПАБЫс/ЫеАБЫсСзЫсУИPGAM5БЛЪЭЗХЃЌВЂЫцКѓШЅСзЫсЛЏВЂМЄЛюЫРЭівђзгЃЌДйЕђЭівђзгЯпСЃЬх1ЃЈAIFM1ЃЉЁЃ
ЛЗОГЪЪгІадЕФбѕЛЏЛЙдЕїНк
дкЩњУќЕФећИіЙ§ГЬжаЃЌЩњЮяЬхвРРЕгкгыЭтВПЛЗОГЕФЯрЛЅзїгУЁЃ"БЉТЖзщ"ЃЈExposomeЃЉжИЕФЪЧвЛЩњжаБЉТЖгкИїжжЩњЮяКЭЗЧЩњЮявђЫиЕФФЃЪНЃЌАќРЈжСЙиживЊЕФгЊбјГЩЗжКЭбѕЦјЙЉгІЁЃБЉТЖзщжагааэЖрЛЏбЇКЭЮяРэГЩЗжЃЌР§ШчжТАЉЮяЁЂПеЦјЮлШОЁЂдывєЁЂЕчРыЗјЩфКЭЬЋбєЗјЩфЃЌвдМАИаШОКЭЩњЛюЗНЪНвђЫиЁЃдкШЫРрКЭЦфЫћЖЏЮяжаЃЌБЉТЖзщЯрЛЅзїгУЕФжївЊЮЛжУЪЧЩЯЦЄ-ЛЗОГНчУцЃЌАќРЈЮИГІКЭЗЮВППеМфЁЂЦЄЗєКЭблОІЁЃетаЉЕиЗНЪЧДѓСПЮЂЩњЮяЕФМвдАЃЌЭГГЦЮЊЮЂЩњЮязщЁЃгЊбјЁЂЛЏбЇКЭЮяРэБЉТЖОпгагавцвдМАЧБдкЕФЮЃЯеаЇгІЃЌетШЁОігкМССПКЭЪБПегІгУЁЃМрВтБЉТЖзщЕФаХКХВЂНЋЦфећКЯЕНЯьгІКЭМЧвфЙІФмжаЪЧИїжжгІМЄЗДгІЕФЛљБОНЧЩЋЃЈВЮМћЧАУцЬжТлЕФФкШнЃЉЁЃПђ3ЬсабЮвУЧЙизЂБЉТЖзщЃЌВЂЧПЕїбѕЛЏЛЙдЕїНкКЭЗМЯуЬўЪмЬхЃЈAHRЃЉЕФзїгУЃЌКѓепЪЧвЛИіЁАБЉТЖзщЪмЬхЁБЁЃБЉТЖзщгыЕААзжЪзщЁЂЛљвђзщКЭБэЙлЛљвђзщЕФНчУцЩцМАвЛИібѕЛЏЛЙдЭјТчЃЌИУЭјТчНЋЪфШыаХКХећКЯЮЊЪЪгІадЯИАћЗДгІЪфГіЃЈЭМ1ЃЉЁЃAHRаХКХдкЖрИіЩњУќЙ§ГЬжаЕФИДдгзїгУПЊЪМЕУЕНВћУїЃЌР§ШчдкЫЅРЯЙ§ГЬКЭУтвпЯИАћДњаЛЕФбѕЛЏЛЙдЕїНкжаЁЃбѕЛЏЛЙдБфЛЏЕФМЧвфСІаЇгІвбБЛжЄУїПЩвдЕїНкУтвпгІД№ЁЃ
Пђ3 ЭтдДадБЉТЖгкбѕЛЏЮя
жеЩэБЉТЖгкИїжжЯпЫїКЭгАЯьЕФХфжУЃЌГЦЮЊЁАБЉТЖзщЁБЃЌВЛНіАќРЈжСЙиживЊКЭгавцЕФЃЌЛЙАќРЈгыбѕЛЏЛЙдЕїНкЯрЙиЕФЧБдкЕФгаКІГЩЗжЁЃЦфжазюживЊЕФЪЧгЊбјКЭбѕЦјЙЉгІЃЌЛЙгаЛЏбЇЁЂЮяРэЁЂИаШОКЭЩњЛюЗНЪНЕФБЉТЖзщГЩЗжЁЃбѕЛЏЛЙдаХКХЭјТчзїЮЊБЉТЖзщКЭЯИАћааЮЊБфЛЏгеЕМжЎМфЕФНгПкЁЃ
гЊбјБЉТЖЁЃгЊбјЕФвЛИіЛљБОзїгУЪЧЮЊбѕЛЏЛЙдЕїНкЬсЙЉзщжЏЬивьадКЭЯИАћЬивьадЕФЙЙНЈПщЁЃетАќРЈЮЂСПгЊбјЫиКЭПѓЮяжЪгУгкбѕЛЏЛЙдУИЃЌвдМАЕЭЗжзгСПЕФбѕЛЏЛЙдЛюадЩњЮявђзгЃЌШчЮЌЩњЫиКЭжВЮяЛЏбЇЮяжЪЃЈРрКњТмВЗЫиЁЂЛЦЭЊЕШЃЉЁЃКъСПгЊбјЫиЬсЙЉNADPHКЭFADH2ЕФЕзЮяЧтвдЙЉЩњЮяФмСПЪЙгУЁЃЧБдкЕФгаЖОвћЪГГЩЗжПЩвдЭЈЙ§бѕЛЏЛЙдЙ§ГЬДњаЛКЭНтЖОЃЌЬиБ№ЪЧЕЅМгбѕУИЃЈЯИАћЩЋЫиP450ЃЉЁЂЫЋМгбѕУИКЭЦфЫћбѕЛЏУИЁЃ
ЛЏбЇБЉТЖЁЃЗЧгЊбјЛЏбЇЮяжЪЙЙГЩСЫЛЗОГМСЕФОоДѓШКЬхЃЌгШЦфЪЧЮлШОЮяЃЌетЪЧЖОРэбЇЕФЙизЂНЙЕуЁЃбѕЛЏгІМЄКЭбзжЂЪЧЛЗОГЩЫКІЕФЛљБОЬиеїЁЃЗМЯуЬўЪмЬхЃЈAHRЃЉгУгкЦєЖЏЖдИїжжЛЏбЇБЉТЖЕФбѕЛЏЛЙдЯрЙиЗДгІЃЈЁАБЉТЖзщЪмЬхЁБЃЉЁЃAHRЪЧвЛжжИагІБЉТЖзщЯпЫїЕФХфЬхМЄЛюзЊТМвђзгЃЌЫќМЄЛюСЫЖржжЭООЖЕФИДдгзЊТМЗДгІЃЌВЂЭЈЙ§ЕїНкЦїКЭЙВЕїНкЦїРДЮЂЕїЗДгІЁЃЯёЖўЖёгЂРрЛЏКЯЮяетбљЕФAHRХфЬхгыБэЦЄЩњГЄвђзгЪмЬхЃЈEGFRЃЉЕФЯИАћЭтНсЙЙгђЯрЛЅзїгУЃЌДгЖјгАЯьЦфдкбѕЛЏЛЙдЕїНкжаЕФЙІФмЁЃШЫРрЛЗОГБЉТЖзщЕФЩњЮяМрВтЙЄзїСїГЬЪЧПЩгУЕФЁЃ
ЮяРэБЉТЖЁЃЭЈЙ§ГЌЩњРэбЇЫЎЦНЕФЬЋбєЗјЩфКЭЦфЫћаЮЪНЕФЗјЩфв§Ц№ЕФгаКІбѕЛЏЛЙдЗДгІв§Ц№ЙизЂЁЃетаЉАќРЈЕчРыЗјЩфКЭЕчДХЗјЩфЁЂШШЁЂдывєКЭЛњаЕЬєеНЃЌЖдДЫгаЧПДѓЕФгІМЄЗДгІЁЃР§ШчЃЌдывєв§Ц№ЕФаФбЊЙмКЭЩёОЪЇЕїЪЧЭЈЙ§бѕЛЏЛЙдЙ§ГЬНщЕМЕФЁЃЦЄЗєРЯЛЏПЩФмЪЧгЩзЯЭтЯпЗјЩфЛђПеЦјЮлШОЕФбѕЛЏЬєеНв§Ц№ЕФЁЃ
ИаШОКЭУтвпгІД№ЁЃЯИОњКЭВЁЖОИаШОЪЧЛњЬхЭтВПЬєеНЕФвЛВПЗжЃЌбѕЛЏЛЙдЕїНкдкЗРгљЛњжЦжаЦ№зїгУЁЃР§ШчЃЌбѕЛЏМСдкЮИГІЕРУтвпБЃЛЄжажСЙиживЊЁЃ
ЩњЛюЗНЪНвђЫиЁЃдкБЉТЖзщжаАќКЌЕФИїжжааЮЊФЃЪНжаЃЌЬхг§ЖЭСЖЭЈЙ§діМгNADPHбѕЛЏУИв§ЗЂЕФH2O2ВњЩњвдМАЭЈЙ§діЧПжЌжЪЙ§бѕЛЏзїгУгыбѕЛЏЛЙдЛюадЯрСЌЁЃдкгабѕбЕСЗжаЃЌЯЕЭГадбѕЛЏЛЙдЭЈаХЪЧЭЈЙ§бЛЗЕФЯИАћЭтФвХнНщЕМЕФЁЃбЛЗвђзгЃЌШчЩњГЄвђзгКЭЯИАћвђзгЃЌЭГГЦЮЊдЫЖЏЫиЃЌНщЕМЖЭСЖЖдбѕЛЏЛЙдЦНКтЕФгАЯьЁЃ
НсТлгыеЙЭћ
дкВћЪібѕЛЏЛЙдЕїПиЕФЛљБОдРэЪБЃЌБОзлЪіжМдкЬсЙЉвЛИіИХРРЃЌНщЩмЯИАћЙІФмдкгабѕДњаЛжаШчКЮЮЌГжЕФЛљБОзМдђЁЃЮЊСЫГжајЕФЙІФмдЫзїЃЌЯИАћМАећИіЩњЮяЬхЪЙгУвЛИіаЕїЕФбѕЛЏЛЙдПижЦЯЕЭГЃЌИУЯЕЭГАќРЈЖрЗНУцЕФЪфШыЯпЫїЃЌетаЉЯпЫїЕМжТВњЩњбѕЛЏЛЙдаХКХЃЌВЂЫцКѓзЊЕМжСЬиЖЈФПБъЮЛЕуЁЃетаЉЮЛЕуЗДЙ§РДМЄЛюЛљвђзщЁЂБэЙлЛљвђзщЁЂЕААзжЪзщвдМАЦфЫћЁА-зщЁБЃЌвддквЛИіаЕїЕФЭјТчЛюЖЏжаВњЩњЙІФмадЯьгІНсЙћЁЃзмЬхЩшжУЃЈЭМ1ЃЉгЩЪЪЕБЕФЗДРЁЯЕЭГЭъГЩЃЌетаЉЯЕЭГЛсМгЧПЛђМѕШѕбѕЛЏЛЙдаХКХЃЌЦфЯъЯИЛњжЦШдашНвЪОЁЃМјгкетжжаЕїХЌСІЕФГжајЛюЖЏаджЪЃЌетжжзДЬЌБЛГЦЮЊЮШЬЌЖЏСІбЇЃЌвВБЛХЗбЙСІетвЛЪѕгяЫљАЕЪОЁЃЭЈЙ§ЕААзжЪАыызАБЫсЕФЗвыКѓбѕЛЏаоЪЮЃЈбѕЛЏЛЙдЕААзжЪзщЃЉКЭЭЈЙ§бѕЛЏЛЙдУєИазЊТМвђзгЕФЛљвђМЄЛюЛњжЦЃЈБэ1ЃЉЕФЩњЮяЛЏбЇаХКХвбдкКмДѓГЬЖШЩЯЕУЕНВћУїЁЃДЫЭтЃЌЫцзХбЧЯИАћЦїЕФаТЯрЛЅзїгУБЛЗЂЯжЃЈЭМ3ЃЉЃЌЯИАћФкКЭЯИАћМфЕФбѕЛЏЛЙдЭЈаХЭООЖвВБфЕУИќМгЧхЮњЁЃУшЪіЯИАћбѕЛЏЛЙдЗДгІзщжЏЕФвЛЯЕСаддђЃЌМДбѕЛЏЛЙдДњТыЃЈПђ1ЃЉЃЌУшЪіСЫбѕЛЏЛЙдЕїНкЖдЩњУќЙ§ГЬЕФЙБЯз22ЁЃ
ШЛЖјЃЌШдгаМИИіЙиМќЮЪЬтгаД§ЛиД№ЁЃвЛИіжївЊЮЪЬтЩцМАбѕЛЏЛЙдЕїНкЕФВуДЮНсЙЙЁЃе§ШчвбЗЂБэЮФЯзжаЬсЕНЕФЃЌвдМАдкБОзлЪіЕФМИИіЪЕР§жаЃЌДцдкЫљЮНЕФжїЕїНкЦїЃЌАќРЈФкжЪЭј88ЁЂЗЧБрТыRNA111ЁЂAHR216ЁЂКЫКьЕААз104ЁЂPGC-1ІС217ЁЂmiRNA218ЕШЃЌЕЋЮЪЬтРДСЫЃКЫРДЕїНкЕїНкЦїЃПЛЛОфЛАЫЕЃЌЫРДМрЖНЁАЪиУХШЫЁБ219ЃПећЬхжИЛгЪЧШчКЮдЫзївдНЋВЛЭЌаХКХећКЯЮЊЙВЭЌЯьгІЕФЃЌМДЪВУДддђЙЙГЩМССПаЇгІЕФЛљДЁЃПЪЧЪВУДОіЖЈСЫЁАЩшЖЈЕуЁБЃПР§ШчЃЌШчКЮЪЖБ№ОіЖЈНЁПЕНсЙћгыЯИАћЫРЭіЭООЖжЎМфЕФЁАСйНчЕуЁБЃПвЊИќКУЕиРэНтетаЉЮЪЬтЃЌНЋашвЊдкЪБПеФЃЪНЩЯНјааЭјТчЗжЮіЃЌПМТЧЗДРЁКЭЧАРЁЛиТЗЃЌМДЛљвђБэДяЗДРЁЛиТЗ220ЁЃСэвЛИіашвЊПМТЧЕФЮЌЖШЪЧбѕЛЏЛЙдЕїНкгыЦфЫћжївЊДњаЛЕїНкЗНЪНШчСзЫсЛЏ/ШЅСзЫсЛЏЁЂИЦаХКХКЭЕчЛЏбЇаХКХЕФЯрЛЅзїгУЁЃ
СэвЛИіаТаЫбаОПСьгђЩцМАИїжжРраЭЕФЛюадЮяжжжЎМфИќЯИжТЕФЯрЛЅзїгУЃЌР§ШчROSЁЂRNSЁЂRSSЕШЁЃH2O2зїЮЊжївЊЕФбѕЛЏЛЙдаХКХЗжзгвЛжБЪЧБОзлЪіЕФжиЕуЃЌЕЋЖдЦфЫћROSШчГЌбѕвѕРызгСЌЭЌвЛбѕЛЏЕЊ10ЁЂЕЅЯпЬЌбѕЗжзг221ЁЂМЄЗЂЬЌШЉРр222ЁЂжЌжЪЙ§бѕЛЏЮя223ЛђСђДМЛЏЗДгІдкбѕЛЏЛЙдЕїНкжаЕФзїгУ224ЕШаТМћНте§дкгПЯжЁЃ
е§ШчБОзлЪіЫљЪіЃЌЩњРэбЇЩЯЕФбѕЛЏЛЙдЙ§ГЬЕїНкЙЙГЩСЫНЁПЕ225,226КЭНЁПЕРЯЛЏЙ§ГЬ227ЕФЛљБОЬиеїЁЃгыДЫВЛЭЌЃЌМВВЁЙ§ГЬЪЧгЩбѕЛЏЛЙдЪЇКтКЭбѕЛЏЛЙдгІМЄв§Ц№ЕФЛђгыжЎЯрЙиЁЃетИіПьЫйдіГЄЕФбаОПСьгђдкетРяУЛгаЯъЯИЩцМАЁЃЙигкгаЯоЪ§СПЕФЦїЙйЯЕЭГЃЈР§ШчЃЌаФдрВЁбЇЁЂЩёОбЇЁЂУтвпбЇЁЂДњаЛЁЂЙЧїРМЁЁЂАЉжЂЁЂРЯЛЏКЭЩёОЭЫааадМВВЁЃЉЕФбѕЛЏЛЙдвНбЇЗНУцвбдкЦфЫћЕиЗНЛиЙЫ4,9,228,229,230ЁЃвдЯИАћЬивьадКЭЮЛЕуЬивьадЕФЗНЪНдкЪЪЕБЕФЪБПеДЋЕнжаНјаабѕЛЏЛЙдПижЦЃЌНЋгажњгкОЋШЗбѕЛЏЛЙдвНбЇ231ЕФЗЂеЙЁЃбѕЛЏЛЙдЙ§ГЬЕФвЉРэбЇе§дкДгеыЖдЕЅвЛЗжзгАаЕузЊЯђЭјТчвНбЇЃЌгІгУЯЕЭГЩњЮябЇЗЖЪНРДбаОПВЁвђЛњжЦЖјЗЧжЮСЦжЂзД232,233ЁЃетШЁОігкЖдПЙбѕЛЏМСзїгУЛњжЦЕФИќКУРэНт234ЁЃНјвЛВНВћУїЛЗОГЁЂгЊбјЁЂЮЂЩњЮязщКЭИаШОБЉТЖЕШжеЩэБЉТЖгыЛљвђзщЯрЛЅзїгУЕФЗжзгЯИНкЃЌЪЧвЛИіжиДѓЕФбаОПЬєеНЁЃ
https://blog.sciencenet.cn/blog-41174-1433535.html
ЩЯвЛЦЊЃКSiЧтМСЖдгзЪѓЩёОЖОадЕФБЃЛЄзїгУ
ЯТвЛЦЊЃКЫЅРЯЕФЪБжгЪЧШчКЮдЫзЊ
