ВЉЮФ
жЇдЬхШчКЮШыЧжЕНЯИАћФк  ОЋбЁ
ОЋбЁ
|
жЇдЬхЪєгкШэЬхЪєЃЌвдЦфаЁЛљвђзщДѓаЁЃЈ500-1300 kbЃЉКЭгаЯоЕФЩњЮяКЯГЩФмСІЖјжјГЦЁЃЫќУЧЭЈЙ§ЧжШыИїжжЯИАћРраЭвдзїЮЊЯИАћФкВЁдЬхДцЛюЖјБэЯжГіжТВЁадЁЃеГИНЪЧГЩЙІЧжЯЎЕФЙиМќЯШОіЬѕМўЃЌгЩжЇдЬхБэУцеГИНЫигыЫожїЯИАћФЄЩЯЕФЬивьадЪмЬхжЎМфЕФЯрЛЅзїгУаЕїЖјГЩЁЃЧжЯЎдкКмДѓГЬЖШЩЯвРРЕгкЭјИёЕААзКЭаЁЮбНщЕМЕФФкЛЏЃЌАщгаЖржжЛюЛЏЕФМЄУИЁЂЯИАћЙЧМмжиХХКЭЮоЪ§ЕФаЮЬЌИФБфЃЌР§ШчФЄФкЯнЁЂКЫЗЪДѓКЭОлМЏЁЂЯИАћжЪЫЎжзКЭПеХнЛЏЁЃвЛЕЉжЇдЬхГЩЙІЧжШыЫожїЯИАћЃЌЫќУЧОЭЛсдкФвХнЁЂЯИАћжЪЁЂКЫжмЧјгђКЭЯИАћКЫжаНЈСЂгаЕЏадЕФБгЛЄЫљЃЌЦфжаЬиЖЈЕФЛЗОГЬѕМўгаРћгкГЄЦкЩњДцЁЃЫфШЛШмУИЬхНЕНтКЭздЪЩПЩвдЯћГ§ДѓЖрЪ§ШыЧжЕФжЇдЬхЃЌЕЋвЛаЉЛюЯИОњПЩвдЭЈЙ§АћЭТзїгУЪЭЗХЕНЯИАћЭтЛЗОГжаЃЌетЪЧбгГЄИаШОГжајадЕФЙиМќвђЫиЁЃетЦЊзлЪіЬНЬжСЫжЇдЬхЧжШыЫожїЯИАћВЂЪЙЦфФбвдзНУўЕФЩњДцгРОУЛЏЕФИДдгЛњжЦЃЌжМдкЧПЕїИљГ§етжжЩёУиЯИОњЕФЬєеНЁЃ
Xiu F, Li X, Liu L, Xi Y, Yi X, Li Y, You X. Mycoplasma invasion into host cells: An integrated model of infection strategy. Mol Microbiol. 2024 Apr;121(4):814-830. doi: 10.1111/mmi.15232. Epub 2024 Jan 31. PMID: 38293733.
ПЦбЇЭјЁЊжЇдЬхИаШО - ЫябЇОќЕФВЉЮФ (sciencenet.cn)
ПЦбЇЭјЁЊ[зЊди]жЇдЬхЁЊЁЊвўаЮЧвзюаЁЕФЯИОњВЁдЬхЃЌЛЙЛсгеЗЂЁАЗЮЭтжЂзДЁБ - ХЃвЋЗМЕФВЉЮФ (sciencenet.cn)
1 ЧАбд
жЇдЬхЪЧзюаЁЕФздЮвИДжЦЩњЮяжЎвЛЁЃЫќУЧОпгабЯИёЕФЫожїЗЖЮЇКЭзщжЏЬивьадЃЌВЂЧвЖдШЫРрЁЂЖЏЮяКЭжВЮяЕФЖржжМВВЁИКгад№ШЮЁЃР§ШчЃЌдкШЫРржаЃЌжЇдЬхгыМБадКєЮќЕРМВВЁЁЂЙиНкИаШОвдМАЩњжГЕРКЭФђЕРИаШОгаЙиЁЃЖЏЮяПЩФмЛсЛМЩЯДЋШОадХЃаиФЄЗЮбзКЭДЋШОадЩНбђаиФЄЗЮбзЁЃЯрБШжЎЯТЃЌжВЮяПЩФмЪмЕНЛЦЛЏВЁЁЂХЎЮзЩЈАбВЁКЭИЬщйЭчЙЬВЁЕФгАЯьЁЃОЁЙмжЇдЬхИаШОЭЈГЃЪЧздЯоадЕФЃЌЕЋФГаЉжЇдЬхФмЬгБмУтвпМрЪгЃЌГЃдкЫожїЬхФкНЈСЂТ§адИаШОЁЃР§ШчЃЌЮЂаЁыхдЬхдкУкФђЩњжГЕРжаПЩДцЛюГЄДя14ЬьЁЃетаЉЛњжЦАќРЈЗжзгФЃФтЁЂПЙдБфвьЁЂПЙбѕЛЏгІМЄПЙадЁЂЩњЮяФЄаЮГЩЁЂжаадСЃЯИАћЯнкхНЕНтЁЂУтвпЯЕЭГЕїНкКЭЯИАћФкЩњДцЁЃОЁЙмЯИАћКЭЬхвКУтвпдкИаШОКѓЯћГ§ДѓЖрЪ§жЇдЬхЗНУцОпгаЧПДѓЕФзїгУЃЌЕЋдкЖЬЦкФкЭъ؎Țç¨дЬхЪЧвЛИіМшОоЕФЬєеНЁЃетвЛРЇФбжївЊдДгкетаЉГєУћебжјЕФжЇдЬхФмЙЛЧжШыАќРЈЩЯЦЄЯИАћЁЂФкЦЄЯИАћЁЂГЩЯЫЮЌЯИАћКЭУтвпЯИАћдкФкЕФИїжжЫожїЯИАћРраЭЃЈБэ1ЃЉЁЃетЮЊГЄЦкЯИАћФкДцЛюЦЬЦНСЫЕРТЗЃЌИјСйДВжЮСЦДјРДСЫЯрЕБДѓЕФЬєеНЁЃ
2БЛжТВЁаджЇдЬхЧжШыЕФЫожїЯИАћЁЃ
жЇдЬхШыЧжЩцМАвЛЯЕСаИДдгЧвОЋЯИЕФЪТМўЁЃЕБдтгіЫожїЪБЃЌжЇдЬхЭЈЙ§ИїжжеГИНЫигыЯИАћЭтЛљжЪЛђЬиЖЈЕФЫожїЯИАћБэУцЪмЬхНсКЯРДЦєЖЏШыЧжЁЃетжжНсКЯДЅЗЂСЫвЛЯЕСаЪТМўЃЌАќРЈЯИАћГЌЮЂНсЙЙЕФИФБфЁЂЯИАћЙЧМмЕФжиХХвдМАИїжжМЄУИЕФМЄЛюЁЃЫцКѓЃЌжЇдЬхЭЈЙ§ЭјИёЕААзЛђаЁЮбЕААзНщЕМЕФФкЭЬзїгУНјШыЫожїЯИАћЃЌВЂБЛдЫЪфЕНдчЦкФкЬхЁЃДцЛюЕФжЇдЬхПЩвдБЛдЫЪфЕНЭэЦкФкЬхЁЃОЁЙмШмУИЬхЛђздЪЩПЩвдНЋжЇдЬхБъМЧЮЊНЕНтФПБъЃЌЕЋгааЉФмЙЛЭЈЙ§дйбЛЗФкЬхНјааЭтХХЃЈЭМ1ЃЉЁЃдкИДдгЕФШыЧжЙ§ГЬжаЃЌжЇдЬхЛёШЁБиашгЊбјЫиЁЂгеЕМЖОЫиВњЩњЃЌВЂЭЈЙ§ШХТваХКХзЊЕМЭЈТЗКЭЯИАћЙЧМмзщжЏЃЌЦЦЛЕЫожїЯИАћЩњРэЙІФмЕФЮЂУюЦНКтЁЃШЛЖјЃЌгЩгкЩњГЄЛКТ§вдМАШыЧжЙ§ГЬжааэЖргАЯьвђЫижЎМфИДдгЕФЯрЛЅзїгУЃЌЮвУЧЖджЇдЬхЕФШыЧжЛњжЦШдДцдкЯрЕБДѓЕФШЯЪЖВюОрЁЃдкБОзлЪіжаЃЌжМдкНщЩмжЇдЬхШыЧжЕФИДдгЙ§ГЬКЭЖРЬиЬиЕуЃЌЗсИЛЮвУЧЖдЦфгыЫожїЯИАћжЎМфДэзлИДдгЕФЯрЛЅзїгУЕФРэНтЁЃ
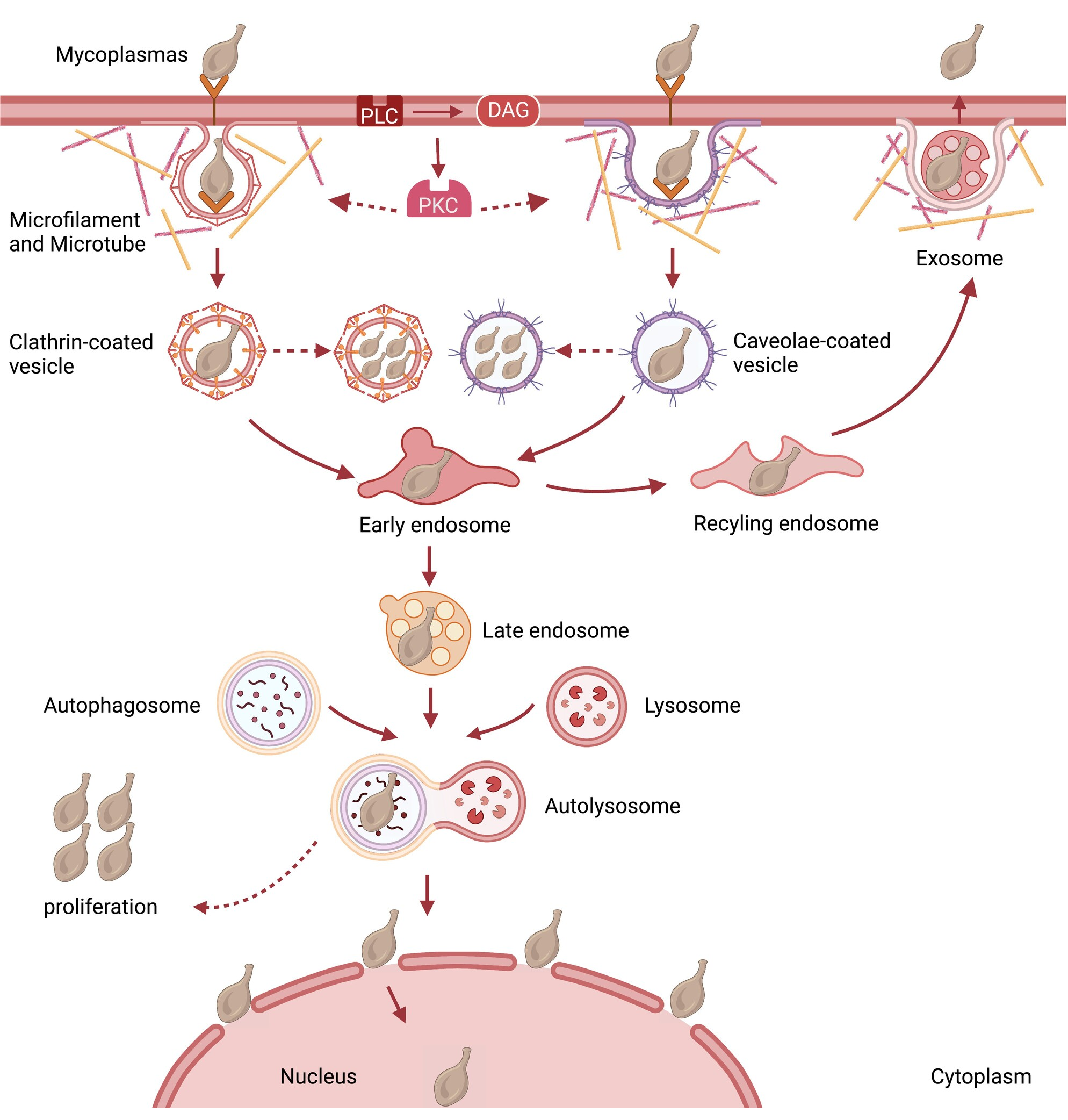
ЭМ1ЃКжЇдЬхЧжШыЫожїЯИАћЕФФЃаЭЁЃжЇдЬхБэУцеГИНЫигыЫожїЯИАћЪмЬхЯрЛЅзїгУЃЌв§ЗЂгЩЮЂЫПЁЂЮЂЙмКЭМЄУИаЭЌЕїПиЕФвЛЯЕСаЯИАћЪТМўЁЃж№НЅЕиЃЌЗЂЩњФЄФкЯнЃЌаЮГЩЫцКѓБЛдЫЪфЕНдчЦкФкЬхЕФЭјИёЕААзЛђаЁЮбЕААзАќБЛЕФФвХнЁЃДѓЖрЪ§жЇдЬхдкФЄАќЮЇЕФФвХнФкДцЛюЃЌЧАЭљЭэЦкФкЬхВЂгыШмУИЬхШкКЯвдЪЕЯжЯИАћФкЩњДцЁЃШЛЖјЃЌвЛВПЗжжЇдЬхЧќЗўгкШмУИЬхНЕНтЛђздЪЩЃЌЖјСэвЛаЉдђБЛдЫЪфЕНдйбЛЗФкЬхЃЌВЂЭЈЙ§ЭтХХзїгУЪЭЗХЕНЯИАћЭтЛЗОГжаЁЃЭМЦЌИФБрздNishiumiЕШШЫЃЈ2017ЃЉMicrobiologyOpenЁЃ
Бэ1ЃКБЛжТВЁаджЇдЬхЧжШыЕФАаЯИАћЁЃ

жТВЁаджЇдЬхЧжШыЫожїецКЫЯИАћБЛШЯЮЊЪЧЕпИВУтвпЯЕЭГКЭНЈСЂТ§адГжајИаШОЕФЪзвЊЪТМўЁЃетЪЧвђЮЊЫќУЧЮЊИїжжШЫРрКЭЖЏЮяжЇдЬхДДдьСЫЩњДцПеМфВЂЛёШЁСЫБиашЕФгЊбјЮяжЪЁЃжЕЕУзЂвтЕФЪЧЃЌВЂЗЧЫљгажЇдЬхЖМгаЧжШыЫожїЯИАћЕФЧуЯђЃЌвВВЂЗЧЫљгаШЮвтецКЫЯИАћЖМвзЪмЧжШОЁЃжЇдЬхПЩЧжШыАќРЈЩЯЦЄЯИАћЁЂФкЦЄЯИАћЁЂГЩЯЫЮЌЯИАћФЫжСУтвпЯИАћдкФкЕФЖржжЯИАћРраЭЃЈБэ1ЃЉЁЃ
ШЫРрЕФжЇдЬхАќРЈЗЮбзжЇдЬх(M. pneumoniae)ЁЂЩњжГЕРжЇдЬх(M. genitalium)ЁЂШЫаЭжЇдЬх(M. hominis)ЁЂЗЂНЭжЇдЬх(M. fermentans)ЁЂДЉЭИжЇдЬх(M. penetrans)КЭЮЂаЁыхдЬх(U. parvum)ЁЃетаЉИКд№в§Ц№КєЮќЕРКЭУкФђЩњжГЕРИаШОЁЃМИКѕЫљгаЕФжЇдЬхЖМФмЧжШыHeLaЯИАћЃЌМДЙуЗКгУгкбаОПЯИАћФкЯИОњХрбјКЭВЁдЬхЖОСІЛњжЦЕФгРЩњЛЏЙЌОБАЉЩЯЦЄЯИАћЯЕЁЃHep-2ШЫРрКэАЉЩЯЦЄЯИАћЯЕЭЈГЃгУгкбаОПЗЮбзжЇдЬхЁЂЩњжГЕРжЇдЬхКЭДЉЭИжЇдЬхЕФШыЧжЁЃЫќгыHeLaЯИАћгааэЖрЙВЭЌЬиеїЃЌВЂЧввзгкВйзїЁЃГ§СЫетаЉГЃгУЕФЯИАћЯЕЃЌЗЮбзжЇдЬхЛЙФмЧжШыАќРЈЦјЕРЩЯЦЄЯИАћЁЂжзСібмЩњЯИАћКЭдДње§ГЃШЫРржЇЦјЙмЩЯЦЄЯИАћдкФкЕФЦфЫћЖржжЯИАћРраЭЁЃДЫЭтЃЌЗЮбзжЇдЬхФмЧжШыОоЪЩЯИАћЃЌЖјШЫаЭжЇдЬхЭЈЙ§ДЉЭИОЋзгЭЗВПКЭжаЖЮЧјгђЕФАћжЪПеМфЯдЪОГіЖРЬиЕФШыЧжФЃЪНЃЌБэУїЦфгыЩњжГЯЕЭГЕФИДдгЙиЯЕЁЃжЕЕУзЂвтЕФЪЧЃЌЗЂНЭжЇдЬхФмЙЛГЩЙІЕиЬгБмУтвпЗДгІКЭПЙЩњЫиЃЌЭЈЙ§дкЭЬЪЩЬхЛђЭЬЪЩШмУИЬхФкЩњДцРДБЃЛЄздМКЕФАВШЋЁЃ
ЖЏЮяжаЕФжТВЁаджЇдЬхЃЌАќРЈМІАмбЊжЇдЬх(M. gallisepticum)ЁЂЖрбљаджЇдЬх(U. diversum)ЁЂЛЌвКжЇдЬх(M. synoviae)ЁЂХЃжЇдЬх(M. bovis)ЁЂжэБЧжЇдЬх(M. hyorhinis)ЁЂЩНбђжЇдЬх(M. agalactiae)ЁЂжэжЇдЬх(M. suis)КЭжэЗЮбзжЇдЬх(M. hyopneumoniae)ЃЌв§Ц№МвЧнЁЂМваѓКЭвАЩњЖЏЮяЕФЗЮбзЁЂШщЯйбзЁЂЙиНкбзЁЂЖњбзКЭЩњжГЯЕЭГМВВЁЁЃHeLaЯИАћвВвзЪмM. gallisepticumЁЂM. agalactiaeКЭM. suisЕШВЁдЬхЕФЧжЯЎЁЃОЁЙмШчДЫЃЌжЇдЬхдкЫожїбмЩњЯИАћжаБэЯжГіИќИпЕФШыЧжаЇТЪЃЌе§ШчMatyushkinaЕШШЫЃЈ2016ЃЉЫљЪОЁЃЫћУЧЪЙгУHeLaЁЂМІКьбЊЧђФИЯИАћЯЕЃЈHD3ЯИАћЃЉКЭаЁЪѓХпЬЅИЩЯИАћЃЈmESЯИАћЃЉРДЗжЮігыЫожїЯИАћЕФЯрЛЅзїгУЁЃЫћУЧЗЂЯжM. gallisepticumдкМІбмЩњЯИАћжаБэЯжГізюИпЕФШыЧжаЇТЪЃЌБэУїHD3ЯИАћзїЮЊM. gallisepticumЬхФкШыЧжЕФзюНгНќФЃаЭЁЃГ§СЫHep-2ЯИАћЃЌM. synoviaeЛЙФмЧжШыМІКьЯИАћЁЂШэЙЧЯИАћЃЈCCHЯИАћЃЉЁЂХпЬЅЯИАћЯЕЃЈCEC-32ЃЉКЭЛЌФЄЧЪЯИАћЁЃM. bovisФмЧжШыХпЬЅаЁХЃБЧМзЯИАћЁЂХпЬЅХЃЦјЙмЯИАћЁЂЗЮЯИАћЁЂбзжЂЯИАћЁЂИЮЯИАћКЭЩіаЁЙмЩЯЦЄЯИАћЁЃДЫЭтЃЌM. gallisepticumЁЂM. synoviaeЁЂM. bovisКЭM. suisЖдКьЯИАћОпгаУїЯдЕФЧїадЃЌВЂФмдкетаЉЯИАћФкДцЛюНЯГЄЪБМфЁЃM. suisЕФШыЧжв§ЗЂбЯжиЕФШмбЊЃЌетПЩФмЪЧШмбЊадЦЖбЊЗЂеЙЕФЙиМќвђЫиЁЃжЕЕУзЂвтЕФЪЧЃЌЭЌвЛжЇдЬхЕФШыЧжаЇТЪдкВЛЭЌЫожїЯИАћжагаЫљВЛЭЌЁЃР§ШчЃЌM. bovisдкЭтжмбЊЕЅКЫЯИАћжаЕФЪ§СПИпгкКьЯИАћЁЃ
3 ВЮгыжЇдЬхЧжЯЎЕФ№ЄИНЫиКЭЪмЬх
ЯИАћеГИНДњБэСЫжЇдЬхБэУцеЙЪОЕФеГИНЫигыЫожїзщЗжжЎМфЕФвЛжжИДдгЕФЯрЛЅзїгУЁЃетЪЧИаШОКЭЫцКѓЕФЯИАћШыЧжЕФГѕЪМЧвБивЊВНжшЃЌетвЛЕуЛљгкШБЗІеГИНФмСІЕФЭЛБфЬхЪЇШЅСЫШыЧжФмСІЁЃШЛЖјЃЌФГаЉЗЧЧжШыадЕФжЇдЬхФмЙЛеГИНЕНЫожїЯИАћЩЯЃЌетБэУїЯИАћеГИНЪЧИаШОЕФЯШОіЬѕМўЃЌзюжеЮЊШыЧжЕФЦєЖЏзіКУСЫзМБИЃЌОЁЙмЫќВЂВЛБиШЛДЅЗЂжЇдЬхЕФШыЧжЁЃвђДЫЃЌеГИНКЭШыЧжЫЦКѕЪЧЖРСЂЕЋЯрЛЅЙиСЊЕФЙ§ГЬЃЌЫќУЧаЭЌзїгУЁЃ
3.1 №ЄИНЫи
еГИНЫиЭЈЙ§СНжжЙиМќЛњжЦНщЕМЯИАћШыЧжЃКЯИАћЭтЛљжЪ(ECM)НЕНтКЭПЙдБфвьЃЈБэ2ЃЉЁЃM. pneumoniaeКЭM. genitaliumЕФеГИНЛњжЦвбБЛЙуЗКбаОПЃЌАќРЈЭЈЙ§ФЉЖЫЦїЙйЕФМЋадНсЙЙНјааЕФЯИАћеГИНКЭЛЌаадЫЖЏЁЃдкетаЉжаЃЌM. pneumoniaeЕФP1ЁЂP40/P90еГИНЫиКЭM. genitaliumЕФP140КЭP110еГИНЫиаЮГЩСЫБЛГЦЮЊЁАNapЁБЕФвьдДЖўОлЬхЖўОлЬхЁЃетаЉеГИНЫиЖЈЮЛЕНФЉЖЫЦїЙйЫЦКѕЪЧЦєЖЏЖЈжВЕФгааЇВпТдЁЃM. pneumoniaeЭЈЙ§ФЉЖЫНсЙЙЕФОлМЏеГИНЫиГЩЙІЧжШыЫожїЯИАћЃЌаЮГЩСЫвЛИігЩеГИНЫиКЭЦфЫћгыЯИАћеГИНЯрЙиЕФЕААзжЪШчP30ЁЂP65КЭHMW1-HMW3зщГЩЕФИДдгЭјТчЁЃетИіЭјТчЪьСЗЕиАѓЖЈЕНЫожїЯИАћФЄЩЯЃЌв§ЗЂвЛЯЕСааХКХзЊЕМЪТМўЁЃЭЌЪБЃЌгыФЉЖЫНсЙЙЯрЙиЕФЛЌаадЫЖЏЭЈЙ§ДйНјЯИАћВуДЉЭИРДдіМгШыЧжЕФПЩФмадЃЈTaylor-Robinson & Jensen, 2011ЃЉЁЃДЫЭтЃЌM. pneumoniaeЕФБэУцеЙЪОЕФЬЧНЭНтУИЃЌШчБћЭЊЫсЭбЧтУИA-CЁЂИЪгЭШЉ-3-СзЫсЭбЧтУИЁЂШщЫсЭбЧтУИЁЂСзЫсИЪгЭЫсЛЅБфУИЁЂБћЭЊЫсМЄУИКЭзЊЭЊУИЃЌгыECMЕААзЯрЛЅзїгУЃЌАќРЈЯЫЮЌЕААздЁЂЯЫЮЌСЌНгЕААзЁЂШщЬњЕААзЁЂВуеГСЌЕААзКЭВЃСЌЕААзЁЃетБэУїЃЌОЁЙмдкНјЛЏЙ§ГЬжаЪЇШЅСЫДѓСПЖдЩњУќЛюЖЏБиашЕФЙІФмЛљвђЃЌЕЋетаЉЬЧНтЛљвђЕФЙІФмЖрбљадЮовЩдквЛЖЈГЬЖШЩЯВЙГЅСЫЖОСІЛљвђЪ§СПЕФВЛзуЃЌДгЖјзюДѓЛЏСЫЫќУЧЕФжТВЁЧБСІЃЌАќРЈеГИНКЭШыЧжЛюЖЏЁЃеГИНЫиP140КЭP110гыM. genitaliumЕФШыЧжУмЧаЯрЙиЃЛШЛЖјЃЌОЇЬхНсЙЙЯдЪОP140ВЂВЛжБНгВЮгыеГИНЃЌЖјЪЧЧЖШыдкP110жЎМфЃЌетБэУїЫќдкЯИАћеГИНЙ§ГЬжаПЩФмЦ№зХЮШЖЈP110ЕФзїгУЁЃ
TABLE 2. ВЮгыШыЧжЕФ№ЄИНЫи.
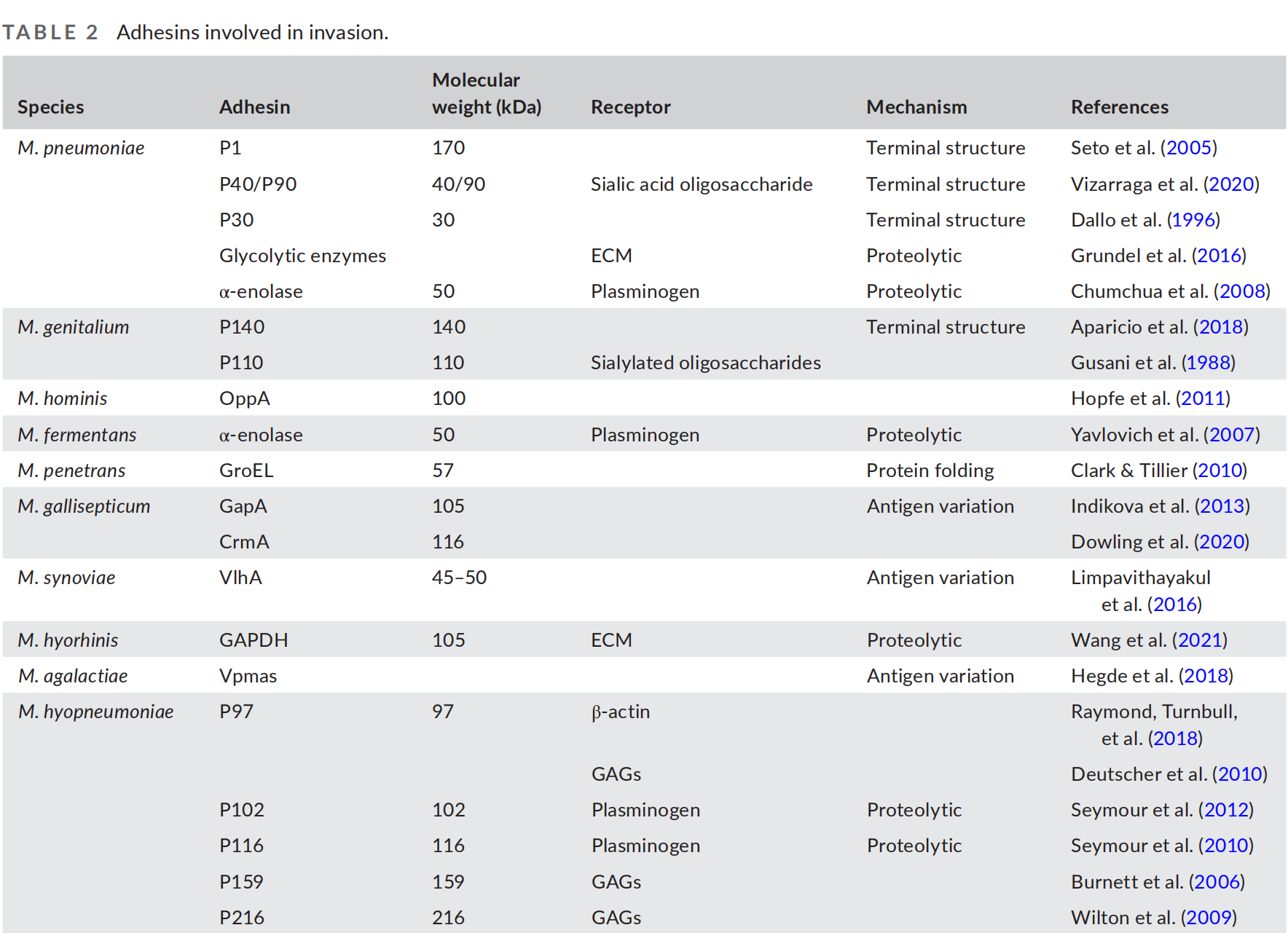
еГИНЫиНщЕМЕФШыЧжЪЧЖЏЮяжЇдЬхЕФвЛжжЦеБщЛњжЦЃЌР§ШчM. hyopneumoniaeБэДяИїжжеГИНЫиШчP97ЁЂP159ЁЂP216КЭP271ЁЃетаЉеГИНЫиЭЈЙ§ОВЕчСІгыКєЮќЕРЯЫУЋЩЯЕФЬЧАЗОлЬЧЃЈGAGsЃЉЕФИпЕчКЩЧјгђЯрЛЅзїгУЃЌУшЛцГівЛжждкВЁдЬхБэУцНсКЯЕААзКЭЫожїЯИАћБэУцжЎМфНЈСЂЮШЖЈЧХСКЕФгааЇВпТдЁЃM. hyopneumoniaeеаФМGAGНсКЯеГИНЫиЕААзвдНсКЯИїжжБиашЕФЫожїЗжзгЕФФмСІЃЌПЩФмДѓДѓгажњгкM. hyopneumoniaeЕФЖЈжВКЭЩЯЦЄЯИАћЕФШыЧжЁЃОЁЙметаЉеГИНЫиЖджЇдЬхШыЧжЕФШЗЧаЙБЯзЩаВЛЧхГўЃЌЕЋЫќУЧдкжЇдЬхГѕДЮеГИНЕНЫожїЯИАћжаЦ№зХЙиМќзїгУЁЃШЛЖјЃЌжЇдЬхШыЧжЩцМАЖржжеГИНЫиЕФЖрЗНУцЯрЛЅзїгУЃЌЭЙЯдСЫИЈжњЗжзгдкДйНјЯИАћеГИНКЭШыЧжжаЕФживЊзїгУЁЃ
OppAЪЧM. hominisжагыШыЧжЯрЙиЕФеГИНЫиЃЌЫќдкеГИНжаЦ№зХЙиМќзїгУЁЃЦфЙІФмЯрЙиЧјгђЕФЭЛБфЛсЫ№КІеГИНЛюадЃЌДгЖјМѕЩйжЇдЬхЕФШыЧжФмСІЁЃДЫЭтЃЌЖдM. synoviaeЗжРыжъЕФађСаЗжЮіНвЪОСЫNЖЫVlhAЕААзжаБрТыИЛКЌИЌАБЫсжиИДађСаЃЈPRPЃЉЕФИпЖШЖрЬЌадЧјгђЁЃОпгаНЯГЄPRPЧјгђЕФОњжъдкM. synoviaeжаБэЯжГіИќИпЕФШыЧжадЁЃЭЌбљЃЌM. agalactiaeеЙЪОЖржжБэУцжЌЕААзЃЈVpmasЃЉНјааЛюдОЕФЯрБфЛЏЁЃдкСљжжVpmasЃЈVpmaU-ZЃЉжаЃЌVpmaVЪЧзюживЊЕФЖОСІвђзгЃЌВЂЯдЪОГізюИпЕФеГИНКЭШыЧжТЪЁЃетаЉЗЂЯжЙВЭЌЧПЕїСЫФГаЉжЇдЬхЭЈЙ§ИДдгЕФПЙдБфвьЛњжЦЕФЯджјЪЪгІадЃЌЪЙЕУеГИНЫиФмЙЛЭЈЙ§БэаЭзЊЛЛЯьгІЛЗОГБфЛЏЖјгХЛЏЁЃетжжЯжЯѓзюжегаРћгкжЇдЬхЕФеГИНКЭШыЧжЃЌЭЙЯдСЫжЇдЬх-ЫожїЯрЛЅзїгУЕФЖЏЬЌаджЪЁЃ
3.2 ЪмЬх
ЮвУЧФПЧАЖдНщЕМжЇдЬхШыЧжЕФЫожїЯИАћЪмЬхЕФСЫНтШдШЛгаЯоЃЛШЛЖјЃЌЮоЭйвКЫсЬЧЕААзЁЂЭйвКЫсЬЧЙВщюЮяКЭСђЫсЛЏЬЧжЌдкНщЕМM. pneumoniaeКЭM. genitaliumЕФИНзХКЭНјШыжаЕФзїгУвбБЛЭЛГіЯдЪОЁЃЭйвКЫсЪЧДцдкгкеГФЄБэУцЕФДјИКЕчЕФЧзЫЎЬМЫЎЛЏКЯЮяЃЌЪЧЭйвКЫсЬЧЕААзЕФзщГЩВПЗжЁЃгЩгкЫќУЧЕФЗжВМЙуЗККЭБиВЛПЩЩйЕФзїгУЃЌВЁдЬхвбОНјЛЏГіеыЖдЫожїЭйвКЫсЕФеГИНЁЂФЃЗТКЭНЕНтЕФВпТдЁЃдкM. pneumoniaeжаЃЌЭйвКЫсзїЮЊИНзХЕНЫожїЯИАћЕФЪмЬхЁЃжЕЕУзЂвтЕФЪЧЃЌгУЩёОАБЫсУИдЄДІРэЫожїЯИАћЃЌетжжУИПЩвдШЅГ§ФЉЖЫЭйвКЫсВаЛљЃЌвжжЦСЫM. pneumoniaeЕФИНзХЁЃЭЌбљЃЌЭйвКЫсЛЏЕФЙбЬЧзїЮЊжївЊЫожїЪмЬхЃЌдкM. genitaliumИаШОЦкМфгыP110ЯрЛЅзїгУЁЃетжжЯрЛЅзїгУЩцМАЩёОАБЫсВПЗжгыSer456-Phe457-Ser458Ш§ыФЕФНсКЯЃЌВЂдкЯИАћеГИНжаЦ№зХжСЙиживЊЕФзїгУЁЃДЫЭтЃЌСђЫсЛЏЬЧЙВщюЮядкШЫРрЩњжГЕРжаЗсИЛЃЌВЂЧвЪЧM. genitaliumКЭM. hominisЕФЧБдкЪмЬхЁЃЪТЪЕЩЯЃЌБЁВуЩЋЦзИВИЧНсКЯВтЖЈвбШЗШЯСђЫсЬЧжЌгыM. hominisЕФЬивьадНсКЯЃЌДйНјжЇдЬхЕФЖЈжВКЭШыЧжЁЃДЫЭтЃЌЭйвКЫсвбБЛШЗЖЈЮЊM. synoviaeКЭM. gallisepticumЯИАћеГИНЕФЬиЖЈЪмЬхЁЃМІШэЙЧЯИАћжажївЊЕФЭйвКЫсЛЏЬЧЕААздМЮЊ70 kDaЃЌЖјдкCEC-32ЯИАћжадМЮЊ60 kDaЁЃШчЧАЫљЪіЃЌM. hyopneumoniaeБэУцБэДяЕФаэЖреГИНЫивдСбНтЦЌЖЮЕФаЮЪННсКЯЕНИпЖШСђЫсЛЏЕФGAGsЁЃОЁЙметжжЧХНгЛњжЦдкУЛгааЮГЩзЈгУгкетаЉеГИНЫиЕФЬиЖЈЪмЬхЕФЧщПіЯТдЫзїЃЌЕЋЫќвтЮЖзХжэКєЮќЕРЩЯЦЄЯИАћЯЫУЋБэУцЕФGAGsаоЪЮПЩФмзїЮЊM. hyopneumoniaeБэУцеГИНЫиЕФКђбЁЪмЬхЁЃ
зюНќЃЌЮвУЧЖджЇдЬхФкЛЏЕФРэНтвбОРЉеЙЕНАќРЈФЃЪНЪЖБ№ЪмЬхЕФВЮгыЁЃдкетаЉЪмЬхжаЃЌTollбљЪмЬх4ЃЈTLR4ЃЉвбОГЩЮЊM. pneumoniaeФкЛЏЕФвЛИіЪмЬхЃЌРЉеЙСЫЦфдкЪЖБ№ИяРМвѕадЯИОњжаЕФжкЫљжмжЊЕФзїгУЁЃжЕЕУзЂвтЕФЪЧЃЌетжжФкЛЏЪЧВЛвРРЕTollбљЪмЬх2ЃЈTLR2ЃЉЕФЃЌвђЮЊгУTLR2ШБЯнЕФОоЪЩЯИАћИаШОВЂУЛгаЯћГ§ЯИАћвђзгЕФВњЩњЁЃЯрБШжЎЯТЃЌTLR4КЭTLR2ЫЋжиШБЯнЕФОоЪЩЯИАћЮДФмЗжУкЯИАћвђзгЃЌетБэУїTLR4ПЩФмНщЕМM. pneumoniaeЕФШыЧжЛђЪмЬхФкЛЏЁЃжЕЕУзЂвтЕФЪЧЃЌTLR4НщЕМЕФM. pneumoniaeФкЛЏПЩФмЪЧЯШЬьУтвпЯЕЭГгУРДЯћГ§ИУЩњЮяЬхЕФвЛжжЛњжЦЃЌЖјВЛЪЧM. pneumoniaeгУРДЬгБмЫожїУтвпЗДгІЕФВпТдЁЃдкM. pneumoniaeжаЃЌTLR4ЪЖБ№ЕФХфЬхЫЦКѕЪЧжЌжЪЁЃ
жївЊЯИАћЙЧМмЕААзМЁЖЏЕААзгЩгкЦфгыИїжжЯИОњВЁдЬхШчBзщСДЧђОњЁЂM. suisКЭОќЭХОњЗЮбзЕФЯрЛЅзїгУЖјв§Ц№СЫЬиБ№ЕФаЫШЄЁЃдкM. suisЧжШыКьЯИАћКѓЃЌКьЯИАћФкЕФІТ-МЁЖЏЕААзгыM. suisЕФMSG1ЯрЛЅзїгУЃЌетвтЮЖзХІТ-МЁЖЏЕААзПЩФмзїЮЊM. suisЕФЧБдкЪмЬхЁЃRaymond, MadhkoorЕШШЫЃЈ2018ФъЃЉЪЙгУБэУцЩњЮяЫиЛЏКЭ3DНсЙЙЛЏееУїЯдЮЂОЕжЄУїСЫМЁЖЏЕААздкжэЩЯЦЄЕЅВуЃЈPK-15ЃЉЯИАћЩЯЕФЯИАћЭтДцдкЁЃІТ-МЁЖЏЕААззїЮЊеГИНЫиP97ЕФЪмЬхЃЌНщЕМM. hyopneumoniaeЖдPK15ЯИАћЕФеГИНЁЃвђДЫЃЌІТ-МЁЖЏЕААзПЩФмдкжэЗЮФкзїЮЊM. hyopneumoniaeЕФЙиМќЪмЬхЃЌАяжњКѓајЕФШыЧжЁЃОпЬхРДЫЕЃЌЧђзДІТ-МЁЖЏЕААззїЮЊM. hyopneumoniaeЕФжївЊЪмЬхЃЌНсКЯЖрИіБэУцБЉТЖЕФеГИНЫиЃЌЖјЯЫЮЌзДІТ-МЁЖЏЕААзгыM. hyopneumoniaeЙВЖЈЮЛЁЃ
жЕЕУзЂвтЕФЪЧЃЌЯЫШмУИдзїЮЊЖржжБэУцБЉТЖЕФеГИНЫиЕФЪмЬхЃЌАќРЈЬЧНЭНтУИКЭЗжзгАщТТGroELКЭDNAKЃЌЫќУЧзїЮЊЗжзгВЖзНЦїгыЯЫШмУИдЕФЯИАћБэУцНсКЯЃЌЭЌЪБгыECMзщЗжЯрЛЅзїгУЁЃвЛЕЉНсКЯЃЌБэУцЯрЙиЕФЯЫШмУИддкзщжЏЯЫШмУИдМЄЛюМСЃЈtPAЃЉЛђФђМЄУИаЭЯЫШмУИдМЄЛюМСЃЈuPAЃЉЕФДцдкЯТзЊЛЏЮЊЯЫШмУИЁЃетжжМЄЛюв§ЗЂСЫАщЫцECMНЕНтЕФЕААзЫЎНтМЖСЊЗДгІЃЌЦЦЛЕзщжЏЦСеЯЁЃвЛаЉБэУцЬЧНЭНтУИгыECMЬиЖЈзщЗжЛђЯЫШмУИджЎМфЕФЯрЛЅзїгУБЛЙлВьЕНЁЃР§ШчЃЌІС-СзЫсЯЉДМЪНУИПЩвдЭЈЙ§ИЛКЌРЕАБЫсЕФНсКЯЖЏЛњгыЯЫШмУИдЕФАБЛљФЉЖЫРЕАБЫсНсКЯЮЛЕуНсКЯЁЃетжжЯрЛЅзїгУДЅЗЂСЫЯЫШмУИЕФВњЩњЃЌДгЖјЧаИюECMзщЗжЁЃЭЌбљЃЌжЇдЬхЕФЬЧНЭНтУИЃЌШчИЪгЭШЉ-3-СзЫсЭбЧтУИЃЈGAPDHЃЉЃЌПЩвдзїЮЊеГИНЫиКЭЯЫШмУИдХфЬхЃЌгажњгкECMНЕНтЃЌЮЊжЇдЬхШыЧжДДдьгаРћЬѕМўЁЃ
ЯЫСЌЕААзвВзїЮЊжЇдЬхФкЛЏЕФНсКЯЪмЬхЁЃР§ШчЃЌM. hyopneumoniaeЯИАћдкИаШОВПЮЛИєРыдчЦкБэДяЕФЯЫСЌЕААзЃЌВЂгыећКЯЫиІТ1ЯрЛЅзїгУЃЌв§Ц№ЯТгЮаХКХЪТМўЃЌЕМжТецКЫЯИАћЙЧМмжиХХЃЌДйНјВЁдЬхНјШыЭјИёЕААзАќБЛЕФаЁХнЃЈCCVsЃЉКЭНбХнЁЃетБэУїM. hyopneumoniaeПЩФмНйГжСЫФкдкЕФецКЫећКЯЫиІТ1-ЯЫСЌЕААзЛиЪеЭООЖвдНјШыЫожїЯИАћЃЌдкФЧРяЫќПЩвддкЯИАћФкГжајДцдкЁЃОЁЙметЪЧЮЈвЛвЛЗнЙигкЯЫСЌЕААзКЭећКЯЫиІТ1жЎМфЯрЛЅзїгУЕїНкжЇдЬхФкЛЏЕФБЈЕРЃЌЕЋдкеГИНЕФдчЦкНзЖЮЯЫСЌЕААзЕФНсКЯПЩФмЪЧДѓЖрЪ§ЩѕжСЫљгажЇдЬхЯИАћШыЧжЕФЛљДЁЪТМўЃЌвђЮЊЯЫСЌЕААззїЮЊаэЖржЇдЬхЕФЪмЬхЁЃОЁЙмжЇГжетвЛМйЩшЕФжЄОнгаЯоЃЌЕЋзувдЙФРјНјвЛВНЬНЫїЁЃ
4.жЇдЬхЧжШыЫожїЯИАћ
4.1ЭЈЙ§ФкЭЬзїгУДЉЭИЫожїЯИАћ
ФкЭЬзїгУЪЧжЇдЬхЧжШыЫожїЯИАћЕФжївЊЛњжЦЁЃжЇдЬхЭЈЙ§ЫГађзЊдЫЃЌДгАМЯнЕФФЄНјШыЕНдчЦкФкЬхЃЌШЛКѓЭЈЙ§ЭэЦкФкЬхЁЃетжжОЋаФаЕїЕФНјеЙЭЈЙ§дчЦкФкЬхПЙд1ЃЈEEA1ЃЉКЭRab5ЕФДцдкЕУЕНжЄУїЃЌЫќУЧзїЮЊдчЦкФкЬхНзЖЮЕФБъМЧЮяЃЌвдМАRab7КЭLAMP-1ЃЌЫќУЧжИЪОЭэЦкФкЬхНзЖЮЃЈNishiumiЕШЃЌ2017ФъЃЉЁЃР§ШчЃЌU. parvumгыЛиЪеФкЬхБъМЧЮяRab11ЯрЛЅзїгУЃЌБэУїжЇдЬхФмЙЛДЉдНЛиЪеФкЬхЃЌдкзюжеДгЫожїЯИАћЪЭЗХжЎЧАЁЃАќБЛгаЭјИёЕААзЛђаЁЮбЕФФвХнДйНјU. parvumЕФЩуШЁЃЌЫцКѓгыШмУИЬхШкКЯЃЌЛђдкгыздЪЩБъМЧЮяLC3ЯрЛЅзїгУЪБОРњздЪЩЁЃЛђепЃЌU. parvumПЩвдЭЈЙ§ЭтУкЬхЪЭЗХЕНЯИАћЭтПеМфЁЃвђДЫЃЌВЛФбЯыЯѓетаЉЗжзггажњгкU. parvumдкЩњжГЕРЯИАћжаЕФДЋВЅЁЃЭЌбљЃЌM. hyopneumoniaeПЩвдЭЈЙ§CCVsКЭНбХнБЛФкЛЏЃЌетЭЈЙ§EEA1ЁЂRAB5ЁЂRAB7ЁЂRAB11КЭLAMP1ЕФУтвпгЋЙтдіЧПРДБэУїЁЃАќБЛЫидкНщЕМФкЭЬзїгУжаЕФВЮгыНјвЛВНЕУЕНСЫвЉРэбЇвжжЦМСЕЅЕЄЛЧѕЃПЈДяЮЌКЭТШБћрКЕФжЇГжЃЌетаЉвжжЦМСзЈУХеыЖдCCVЕФаЮГЩКЭФкЬхжаАќБЛЫиЕФЛиЪеЃЌВЂЯджјзшЖЯСЫM. bovisНјШыГѕМЖХпЬЅХЃБЧМзЙЧЃЈPECTЃЉЯИАћЁЃДЫЭтЃЌФкЭЬзїгУвжжЦМСЕЄЛЧѕЃПЈДяЮЌКЭаЁИЩШХRNAвВПЩвдвжжЦжЇдЬхШыЧжЃЌетБэУїАќБЛЫижиСДЃЈCLTCЃЉКЭНбЕААз-1ЃЈCav-1ЃЉВЮгыСЫM. bovisЖдЛЌФЄЯИАћЕФШыЧжЃЌОЁЙмЙлВьЕНM. bovisгыCLTCжЎМфЕФЙВЖЈЮЛБШгыCav-1ИќИпЁЃ
ЦфЫћЗжзгЃЌАќРЈФЄФвЛђАМЯнЁЂжЌЗЄКЭTLR4ЃЌвВЩцМАжЇдЬхШыЧжЁЃР§ШчЃЌЕчзгЯдЮЂОЕНвЪОЃЌдкM. genitaliumЧжШыHLFЯИАћЦкМфЃЌФЄФвЛђАМЯнЕФаЮЬЌЬиеїРрЫЦгкАќБЛгаЭјИёЕААзЕФПгЃЌетБэУїетаЉЗжзгПЩФмжДаагыАќБЛЫиРрЫЦЕФЙІФмЁЃSimvastatinЪЧвЛжжНЕЕЭЕЈЙЬДМЕФвЉЮяЃЌЫќвжжЦСЫжЌЗЄНщЕМЕФФкЭЬЭООЖЁЃChlamydiaвВЪЙгУжЌЗЄНщЕМЕФЭООЖЧжШыЫожїЯИАћЃЌетЬсГіСЫM. bovisПЩФмВЩгУРрЫЦФкЭЬЭООЖЕФПЩФмадЁЃгЩTLR4НщЕМЕФФкЛЏдкжЇдЬхШыЧжжаЦ№зХЙиМќзїгУЁЃдкгыХфЬхНсКЯКѓЃЌTLR4дкжЪФЄЩЯЦєЖЏаХКХДЋЕМЃЌВЂаЮГЩTLR4/Mal/MyD88ИДКЯЮяЃЌЫцКѓЭЈЙ§ФкЬхНјааФкЛЏЁЃShimizuЕШШЫЗЂЯжЃЌM. pneumoniaeИаШОКѓгеЕМДйбзЯИАћвђзгЕФВњЩњЪЧЭЈЙ§TLR4НщЕМЕФФкЭЬзїгУвРРЕадЙ§ГЬЃЌетЭЛГіСЫЦфзїЮЊЪмЬхдкM. pneumoniaeШыЧжКЭФкЛЏвдв§ЗЂбзжЂЗДгІжаЕФживЊзїгУЁЃ
4.2ВЮгыШыЧжЛюЖЏЕФаХКХЭЈТЗ
гызЈадЯИАћФкМФЩњЬхЯрБШЃЌжЇдЬхеЙЯжГіУїЯдНЯЕЭЕФШыЧжаЇТЪЃЌетИјВћУїЫожїЯИАћШыЧжЫљЩцМАЕФаХКХЭЈТЗДјРДСЫЬєеНЁЃР§ШчЃЌЙРМЦжЛгаДѓдМ8%ЕФM. hyponeumoniaeЯИАћДцдкгкЩЯЦЄЯИАћЕЅВужаЁЃДЫЭтЃЌжЇдЬхЕФШыЧжаЇТЪЪмЖржжвђЫигАЯьЃЌМДЪЙдкЭЌвЛжжжЇдЬхЮяжжФквВЛсЕМжТБфвьадЃЌгАЯьЪЕбщЕФПЩжиИДадЁЃвђДЫЃЌФПЧАЖджЇдЬхШыЧжЫљЩцМАЕФаХКХЭЈТЗЕФРэНтШдШЛВЛзуЃЌДѓЖрЪ§ЗЂЯжНіЯогкМИжжЮяжжЃЌШчM. fermentansЁЂM. pneumoniaeКЭФГаЉЖЏЮяжЇдЬхЁЃ1995ФъЃЌAndreevЕШШЫдкHeLaЯИАћжаМјЖЈГівЛжж145 kDaЕФСзЫсЛЏЕААзЃЌетЪЧдкM. penetransИаШОКѓЗЂЯжЕФЃЈAndreevЕШЃЌ1995ФъЃЉЁЃОЁЙметжжСзЫсЛЏгыШыЧжжЎМфЕФШЗЧаЙиЯЕЩаВЛЧхГўЃЌЕЋРвАБЫсСзЫсЛЏПЩФмдкжЇдЬхШыЧжжаЗЂЛгЙиМќзїгУЁЃДЫЭтЃЌОнЭЦВтЃЌM. penetransгыHeLaЯИАћжЎМфЕФЯрЛЅзїгУЕМжТЕААзМЄУИCЃЈPKCЃЉЕФМЄЛюЃЌетЭЈЙ§дкИаШОЛюЕФM. penetransЯИАћКѓ20ЗжжгФкЃЌPKCДгЯИАћжЪзЊвЦЕНжЪФЄЕФЧщПіЕУЕНжЄЪЕЃЌетБэУїPKCПЩФмдкжЇдЬхШыЧжжаЗЂЛгЙиМќзїгУЁЃжЕЕУзЂвтЕФЪЧЃЌдкMolt-3СмАЭЯИАћжаЃЌЙлВьЕНдкM. penetransИаШОКѓжЌжЪзЊЛЛЕФИФБфЃЌАќРЈЖўѕЃИЪгЭКЭздгЩжЌЗОЫсЃЈFFAЃЉВПЗжЕФЛ§РлЁЃетаЉЙлВьПЩФмжЇГжетбљЕФМйЩшЃКM. penetransДЬМЄСзжЌУИЧаИюФЄСзжЌЃЌЕМжТДЅЗЂФЄКЭЯИАћЙЧМмаоЪЮЕФаХКХМЖСЊЃЌДгЖјдЪаэжЇдЬхШыЧжЁЃЧжЯЎаджЇдЬхЃЌАќРЈU. diversumЃЌОпгаСзжЌУИCЃЈPLCЃЉЛюадЃЌетгыЫожїЯИАћЙЧМмзщЗжжиХХКЭаХКХзЊЕМЯрЙиЁЃU. diversumжаДцдкСзжЌУИЛюадБэУїЦфзїЮЊжТВЁвђзгЕФЧБСІЃЌгажњгкЯИАћЫ№ЩЫКЭШыЧжЙ§ГЬЁЃ
4.3 ЫожїЯИАћжаЕФЯИАћЙЧМмжиХХ
вЛаЉЧжЯЎаджЇдЬхдкгыЫожїЯИАћЯрЛЅзїгУКѓЦєЖЏЯИАћЙЧМмжиХХЃЌЭЈЙ§МЁЖЏЕААзЮЂЫПКЭЮЂЙмвРРЕадЙ§ГЬДйНјЫќУЧЕФФкЛЏЁЃетвЛЕуЭЈЙ§гУЮЂЙмКЭЮЂЫПЦЦЛЕвЉЮядЄДІРэРДжЄЪЕЃЌЯдЪОГідкгУМЁЖЏЕААзОлКЯвжжЦМСcytochalasin-DДІРэКѓЃЌФкЛЏЕФM. penetransдкHeLaЯИАћжаЕФЪ§СПМѕЩйЁЃЭЌбљЃЌM. penetransЕФФкЛЏПЩвдБЛЦЦЛЕЮЂЙмЕФtaxolКЭvinblastineвжжЦЁЃетаЉЪ§ОнБэУїЃЌЮЂЫПКЭЮЂЙмЕФОлКЯЖЏЬЌКЭЮШЖЈадЖдгкM. penetransЕФШыЧжжСЙиживЊЁЃM. penetransгыHep-2ЯИАћжЎМфЕФЯрЛЅзїгУгеЕМЯИАћЙЧМмБфЛЏЃЌЭЈЙ§УтвпгЋЙтЙлВьЕНЯИАћЙЧМмзщЗжШчЮЂЙмЕААзЁЂІС-МЁЖЏЕААзКЭФГаЉСзЫсЛЏЕААзЕФОлМЏЁЃДЫЭтЃЌЙВОлНЙМЄЙтЯдЮЂОЕжЄЪЕЃЌдкM. hominisИаШОHeLaЯИАћ48аЁЪБКѓЃЌМЁЖЏЕААзЮЂЫПЕФФ§ОлКЭЯИАћФЄЩЯЭЛЦ№ЕФаЮГЩЃЌЧПЛЏСЫШыЧжгыЫожїЯИАћжаМЁЖЏЕААзЯИАћЙЧМмжиХХЯрЙиЕФЙлФюЁЃШЛЖјЃЌгаељвщЕФНсЙћБэУїЃЌЮЂЙмвжжЦМСnocodazoleНЕЕЭСЫM. gallisepticumШыЧжHeLa-229ЯИАћЕФЦЕТЪЃЌЖјcytochalasin-DУЛгаУїЯдЕФвжжЦаЇЙћЃЌетБэУїM. gallisepticumПЩФмдкЗЧЭЬЪЩадЫожїЯИАћжаВЩгУгыM. penetransВЛЭЌЕФФкЛЏВпТдЁЃДЫЭтЃЌдкPK-15ЯИАћжаИаШОM. hyopneumoniaeКѓИпБэДяЕФМЁЖЏЕААзБэУїЃЌетжжЧжШыадЯИОњПЩФмдкИаШОЦкМфгеЕМЯИАћЙЧМмжиХХвдДйНјЦфЩуШЁЁЃ
4.4 ЫожїЯИАћЧжЯЎКѓГЌЮЂСЃаЮЬЌЕФБфЛЏ
жЇдЬхШыЧждкЫожїЯИАћжав§ЗЂСЫвЛЯЕСааЮЬЌбЇБфЛЏЃЌЦфЯджјЬиеїЪЧдкШыЧждчЦкНзЖЮЯИАћФЄЕФФкЯнЁЃР§ШчЃЌдкM. suisИаШОЕФКьЯИАћжаЃЌЫцзХШыЧжЕФНјеЙЃЌжЪФЄЕФАМЖШМгЩюЃЌзюжеаЮГЩФвХнзДНсЙЙЁЃЯИАћФЄЕФПЊПкдкНјШыКьЯИАћЪБШкКЯЁЃЭЌбљЕиЃЌЕчзгЯдЮЂОЕЯТЙлВьM. bovisШыЧжPECTЯИАћЪБЃЌЗЂЯжФкЯнДІгажБОЖ50ФЩУзЕФПЊПкЃЌвдМАЯИАћФкЕФФвХнКЭАћжЪжаЕФM. bovisФЄБпНчЁЃдкM. bovisШыЧжЛЌФЄЯИАћЙ§ГЬжавВЙлВьЕНСЫРрЫЦЕФФкЯнЁЃжЕЕУзЂвтЕФЪЧЃЌM. genitaliumПЩвддкHLFЯИАћЕФжЪФЄЩЯгеЕМЯђФкЕФСІЃЌаЮГЩБзДАМЯнНсЙЙЃЌЫцКѓВњЩњФЄПкДќЛђПгЁЃетаЉаЮЬЌбЇБфЛЏБэУїЃЌецКЫЯИАћФЄЕФФкЯнЪЧвЛжжЦеБщЕФГЌЮЂНсЙЙБфЛЏЃЌДѓЖрЪ§жЇдЬхВЩгУФкЭЬЭООЖНјааШыЧжЁЃЦфЫћЙлВьЕНЕФБфЛЏАќРЈХЃШщЯйЩЯЦЄЯИАћдкИаШОM. bovisЃЈST172ЛђST173ЃЉ24аЁЪБКѓЯИАћЪеЫѕЁЂАћжЪШОЩЋЩюЁЂЯИАћЭъећадЩЅЪЇЁЃДЫЭтЃЌM. penetransЕФШыЧжПЩЕМжТHep-2ЯИАћЕЅВуЕФЦЦЛЕКЭЯИАћЫРЭіЁЃЛЙгаЃЌвЛВПЗжБЛM. hominisИаШОЕФШЫРрОЋзгБэЯжГіЭфЧњЛђХЬШЦЕФЮВВПКЭжаЖЮдіКёЁЃ
5 гАЯьжЇдЬхШыЧжЕФвђЫи
жЇдЬхдкШыЧжЫожїЯИАћЕФФмСІЩЯДцдкВювьЁЃеГИНЪЧШыЧжЕФЯШОіЬѕМўЃЌЖјЦфЧжШыадЭЈЙ§еГИНЫиКЭжЇдЬхЕФЛЌаадЫЖЏЯджјдіЧПЁЃШЛЖјЃЌвЛаЉжЇдЬхЕФФкдквђЫиЃЌШчЧжШыадКЫЫсУИЃЈMpn133ЁЂMG_186КЭMbovNaseЃЉЃЌвдМАЭтВПвђЫиЃЌШчХрбјЬѕМўКЭЛЏбЇЪдМСЃЌвВдкДйНјжЇдЬхНјШыЫожїЯИАћжаЗЂЛгзїгУЁЃПЙдБфвьвВдіМгСЫФГаЉРДздХЃКЭвАХЃЕФM. bovisЗжРыжъЕФЧжШыадЃЌЮЊжЇдЬхВЩгУЕФЧжШыЛњжЦдіЬэСЫСэвЛВуИДдгадЁЃ
5.1 ФкдквђЫи
ОЁЙмгаЯоЕФжЇдЬхЛљвђзщЯожЦСЫЖдЫожївРРЕЕФДњаЛЧАЬхЕФЛёШЁЃЌЕЋФкдкКЫЫсУИЕФДцдкУжВЙСЫетвЛШБЯнЁЃетаЉКЫЫсУИЃЌАќРЈMpn133ЁЂMG_186КЭMbovNaseЃЌЖдгкКЫЫсЧАЬхКЯГЩЪЧБиашЕФЃЌВЂдкжЇдЬхШыЧжжаАчбнЙиМќНЧЩЋЁЃMpn133ЪЧM. pneumoniaeжаЗЂЯжЕФвЛжжCa2+вРРЕадЯИАћЖОадКЫЫсУИЃЌОпгавЛИіБиВЛПЩЩйЕФЙШАБЫсЁЂРЕАБЫсКЭЫПАБЫсИЛМЏЧјгђЃЈEKSЧјгђЃЉЃЌИУЧјгђЖдгкM. pneumoniaeФкЛЏНјШыA549ЯИАћжСЙиживЊЁЃЩОГ§ИУЧјгђЛсЯћГ§ШыЧжФмСІЃЌетвЛЕуЭЈЙ§EKSБъМЧЕФmCherryЃЈEKS-mCherryЃЉЕААзЕФЯИАћжЪЁЂЮЇКЫКЭКЫЖЈЮЛЕУЕНжЄУїЃЌЖјВЛНіНіЪЧmCherryБОЩэЁЃЭЌбљЕиЃЌMG186ЪЧMpn133ЕФЭЌдДЮяЃЌШБЗІEKSЧјгђЃЌПЩФмВЮгыM. genitaliumЕФШыЧжЃЛШЛЖјЃЌЦфШЗЧазїгУашвЊНјвЛВНбаОПЃЈLiЕШЃЌ2010ФъЃЉЁЃдкM. bovisжаЃЌMBOV_RS02825ЃЈMbovNaseЃЉМШЪЧФЄНсКЯгжЪЧЗжУкаЭКЫЫсУИЃЌОпгаIаЭаХКХађСаЃЌЦфЛюадвРРЕгкTNASE_3ЧјгђЁЃжЕЕУзЂвтЕФЪЧЃЌШБЗІTNASE_3ЧјгђЕФЭЛБфЬхЯдЪОГіУИЛюадЁЂеГИНКЭФкЛЏЗНУцЕФШБЯнЁЃетаЉНсЙћБэУїЃЌMbovNaseдкНщЕМM. bovisНјШыЫожїЯИАћжаЦ№зХЙиМќзїгУЁЃШЛЖјЃЌвВгаПЩФмЙлВьЕНЕФжЇдЬхЭЛБфЬхШыЧжФмСІНЕЕЭгыКЫЫсКЯГЩЪмЯогаЙиЃЌЖјВЛЪЧжБНгНщЕМШыЧжБОЩэЁЃ
жЇдЬхШыЧжвВЪмЕНЦфЫћУИРрШчСзжЌУИКЭЯЫШмУИЕФгАЯьЃЌетаЉУИеыЖдВЂЦЦЛЕЫожїЯИАћФЄЕФЭъећадЁЃдкM. hyorhinisжаЃЌСзжЌУИAЃЈPLAЃЉЧаИюИЪгЭСзжЌЖўѕЅВњЩњИЪгЭ-3-СзЫсЃЌЕМжТЫожїЯИАћФкаЮГЩЙ§бѕЛЏЧтКЭЯИАћЖОадЁЃДЫЭтЃЌPLAНщЕМЕФЫЎНтЪЭЗХздгЩжЌЗОЫсКЭШмбЊСзжЌЃЌетПЩФмЛсЦЦЛЕЫожїЯИАћФЄЕФЭъећадЃЌДйНјM. hyorhinisЕФШыЧжЁЃДЫЭтЃЌМИжжЪовНВЁдЬхЯдЪОГідкЦфБэУцеаФМЯЫШмУИдЕФФмСІЃЌдіЧПЦфзЊЛЏЮЊЯЫШмУИвддіМгЖОСІЁЃетжжаЇгІдкuPA/tPAДцдкЕФЧщПіЯТБШЕЅЖРЕФЯЫШмУИдЯджјдіЧПЁЃР§ШчЃЌдкM. hyopneumoniaeжаЃЌеГИНЫиP102КЭP116дкЩњРэЯрЙиХЈЖШЯТНсКЯЯЫШмУИдЃЌгЩtPAМЄЛюЯЫШмУИЃЌЕМжТЯЫЮЌСЌНгЕААзЕФНЕНтЁЃЬхФкбаОПвВБэУїЃЌдкИаШОЦкМфЯЫШмУИЛюаддіМгЃЌжЇГжСЫM. hyopneumoniaeдкЦфЖЈжВНзЖЮгыЯЫШмУИдЕФЯрЛЅзїгУЁЃетвЛЙ§ГЬгажњгкУтвпЬгБмЁЂНЕНтКЭДЉЭИЫожїзщжЏЃЌДгЖјДйНјЫожїЯИАћЕФГЩЙІШыЧжЁЃетаЉНсЙћЭЛЯдСЫСзжЌУИКЭЯЫШмУИдкжЇдЬхШыЧжЙ§ГЬжаЕФживЊадЁЃзюНќЃЌNunoyaЕШШЫЗЂЯжM. bovisИаШОЖджЇЦјЙмЩЯЦЄЯИАћгаВЛСМгАЯьЃЌАќРЈЯИАћжЪЫЎжзКЭИНзХВПЮЛЕФЯИАћЙЧМмЭјТчЦЦЛЕЁЃетаЉЫ№ЩЫЭЦВтЪЧгЩбѕЛЏМСв§Ц№ЕФЃЌВЂдіМгСЫM. bovisДЉЭИЫожїЯИАћЕФПЩФмадЁЃ
ПЙдБфвьЪЧжЇдЬхШыЧжЕФживЊФкдкОіЖЈвђЫиЁЃетвЛЯджјЯжЯѓдкжюШчM. synoviaeКЭM. agalactiaeетбљЕФжЇдЬхжаЙлВьЕНЃЌЫќУЧВпТдадЕиЧаЛЛеГИНЫиЕФБэДяЃЌАќРЈVlhAКЭVpmasЁЃетгажњгкеГИНКЭжЇдЬхШыЧжЃЌЪЙЫќУЧФмдкЫожїЕаЖдЕФАћФкЛЗОГжаДцЛюЁЃДЫЭтЃЌВЛЭЌЕФM. bovisЗжРыжъБэЯжГіВЛЭЌЕФШыЧжФмСІЃЌетПЩФмЙщвђгкдкШыЧжЙ§ГЬжаОРњДѓСППЙдБфвьЕФПЩБфБэУцжЌЕААзЕФБэДяЁЃ
еГИНЫиЕФЗвыКѓМгЙЄЖджЇдЬхШыЧжжСЙиживЊЁЃР§ШчЃЌM. pneumoniaeЕФP1ЁЂP40КЭP90вдМАM. hyopneumoniaeЕФP159ЁЂP102КЭP216ПЩвддкЖрИіЮЛЕуОРњЕААзЫЎНтМгЙЄЁЃгЩДЫВњЩњЕФЧаИюЦЌЖЮОпгагыИїжжЫожїЗжзгЯрЛЅзїгУЕФФмСІЁЃетжжЕААзЫЎНтЙ§ГЬЯдЪОГівЛЖЈГЬЖШЕФбЁдёадЃЌДгЖјдіЧПСЫжЇдЬхЕФЙІФмЕААзжЪзщПтЃЌВЂЕїНкБэУцЕААзЕФЖрбљадЁЃЫцзХЕААзжЪЧаИюЪТМўЪ§СПЕФдіМгЃЌжЇдЬхгыECMЕААзНсКЯЕФФмСІвВЫцжЎдіМгЁЃетгажњгкЬсИпЫожїЯИАћШыЧжаЇТЪЁЃ
5.2ЭтВПвђЫи
Г§СЫжЇдЬхздЩэЫљгЕгаЕФФкдкЖОСІвђзгЭтЃЌЫожїЯИАћЛЗОГдкЕїНкШыЧжаЇТЪЗНУцвВАчбнСЫживЊНЧЩЋЁЃЮТЖШКЭИаШОГжајЪБМфЪЧгАЯьжЇдЬхИаШОЕФЦеБщвђЫиЁЃгы4ЁуCЯрБШЃЌ37ЁуCЪБЯИАћФкЕФM. penetransЪ§СПЯджјдіМгЃЌЖјЯИАћПеХнЛЏГЬЖШдкИаШО4аЁЪБКѓБШ2аЁЪБКѓИќЮЊУїЯдЁЃетаЉЗЂЯжгыM. penetransЕФзюМбЩњГЄЮТЖШЯрЗћЃЌБэУїбгГЄИаШОЪБМфНјвЛВНгажњгкжЇдЬхЕФФкЭЬзїгУЁЃгЊбјбЙСІЪЧСэвЛИіОіЖЈВЁдЬхШыЧжЕФвђЫиЁЃжЕЕУзЂвтЕФЪЧЃЌМДЪЙдкзюЕЭТэбЊЧхХЈЖШЕФЬѕМўЯТЃЌM. penetransвВФмБэЯжГіЯджјЕФЩњГЄЃЌЖјдк5%ТэбЊЧхДцдкЕФЧщПіЯТЃЌЦфШыЧжФмСІЯджјНЕЕЭЃЌетвтЮЖзХЕЭТэбЊЧхЫЎЦНПЩвдгеЕМжЌжЪЦзЕФБфЛЏЃЌгАЯьM. penetransЕФЧжШыадЁЃДЫЭтЃЌM. gallisepticumЕФЕЭДњДЮШКЬхЃЈRlowЃЉБЃСєСЫШыЧжЗЧЭЬЪЩадецКЫЯИАћЕФФмСІЃЌЖјИпДњДЮШКЬхЃЈRhighЃЉдђЪЇШЅСЫетжжФмСІЃЌЭЌЪББЃГжСЫеГИНФмСІЁЃетаЉЗЂЯжБэУїЃЌM. gallisepticumЕФЗБжГДњДЮПЩФмЛсгАЯьЦфШыЧжЧБСІЃЌОЁЙмдкетаЉОњжъЖрДЮДЋДњКѓПЩФмЗЂЩњЕФЭЛБфЕФЧБдкгАЯьвВВЛШнКіЪгЁЃДЫЭтЃЌЛЏбЇЪдМСДІРэПЩвдЬсИпжЇдЬхЕФШыЧжФмСІЁЃР§ШчЃЌОлввЖўДМ800ПЩвдИФБфЯИАћФЄБэУцЫЎЕФНсЙЙЃЌДйНјM. penetransЕФНсКЯКЭMolt-3СмАЭЯИАћЕФШыЧжЁЃЯрБШжЎЯТЃЌбѕЛЏМСШчЙ§бѕЛЏЧтКЭГЌбѕЮяПЩвдЦЦЛЕЯИАћжЪВЂШХТвЯИАћЙЧМмЭјТчЃЌПЩФмЮЊM. bovisДДдьНјШыЮЛЕуЁЃжЕЕУзЂвтЕФЪЧЃЌгУЕААзУИKдЄДІРэЯджјдіЧПСЫM. hyorhinisеГИНКЭШыЧжКкЩЋЫиСіЯИАћЕФФмСІЃЌПЩФмЪЧЭЈЙ§МЄЛюЛђБЉТЖжЇдЬхБэУцеГИНЫиЁЃвђДЫЃЌСйДВЩЯЪЙгУгАЯьЯИАћФЄЭЈЭИадЕФвЉЮяРДжЮСЦФГаЉИаШОадМВВЁПЩФмЛсдіМгМЬЗЂаджЇдЬхИаШОЕФЗчЯеЁЃ
6 ЯИАћФкЩњДцЕФвтвх
6.1 жЇдЬхЕФЯИАћФкЩњДц
НјШыЫожїЯИАћКѓЃЌжЇдЬхжївЊЖЈЮЛгкЯИАћжЪКЭКЫжмЧјгђЁЃР§ШчЃЌM. pneumoniaeДцдкгкИїжжЯИАћРраЭЕФЯИАћжЪКЭКЫжмЧјгђЃЌШчHep-G2ЁЂN2-AКЭA549ЯИАћЁЃЭЌбљЃЌM. genitaliumБэЯжГіУїЯдЕФКЫжмКЭКЫАаЯђЧуЯђЃЌВЂдкHeLaКЭEM42ЯИАћжаГжајДцдкНЯГЄЪБМфЁЃДЫЭтЃЌU. diversumЁЂM. synoviaeКЭM. canisЙуЗКЗжВМдкетаЉЧјгђЃЌЖјM. hyorhinisЧжШыКкЩЋЫиСіЯИАћЃЌДѓВПЗжФкЛЏЕФжЇдЬхЫцЛњЗжЩЂдкППНќЯИАћФЄЕФЯИАћжЪжаЁЃ
ФГаЉжЇдЬхдкдіжГЙ§ГЬжаОгзЁдкФЄАќЮЇЕФФвХнФкЃЌЕчзгЯдЮЂОЕЙлВьвбжЄЪЕСЫетвЛЕуЁЃР§ШчЃЌM. genitaliumдкVeroЯИАћФкЗБжГЃЌКѓДњжЇдЬхОгзЁдкФвХнжаЁЃM. genitaliumЕФШыЧжЫЦКѕЪЧЖЏЬЌЕФЃЌвђЮЊЫцзХИаШОЪБМфЕФбгГЄЃЌЯИАћФкжЇдЬхЕФЪ§СПЯджјдіМгЃЌВЂжСЩйБЃГж48аЁЪБЕФИпЫЎЦНЁЃЯрЗДЃЌгыM. genitaliumЯрБШЃЌM. penetransЖдHep-2ЯИАћЕФШыЧжЫйЖШЯрЖдНЯПьЃЌФкЛЏЕФЩњЮяЬхвЊУДздгЩДцдкгкЯИАћжЪжаЃЌвЊУДЗтБедкФвХнФкЁЃЖдгкHeLaЯИАћЃЌМДЪЙдкИаШО24аЁЪБКѓЃЌM. penetransвВПЩвдОгзЁдкФкжЪЭјжаЁЃM. bovisдкPECTsжааЮГЩДизДЮяЃЌУЛгаЬиЖЈЖЈЮЛЕНЬиЖЈЕФЯИАћЪвЃЌВЂЧвдк78аЁЪБЕФИаШОЦкФкПЩвддкЯИАћФкдіжГЃЌЕМжТВЁдЬхЙуЗКДЋВЅЁЃШЛЖјЃЌM. gallisepticumВЂВЛзмЪЧЫцзХИаШОНјеЙЖјдіжГЁЃгЩгкздЮвЯћЛЏЛђДгЫожїЯИАћЪЭЗХЃЌФкЛЏЕФжЇдЬхЪ§СППЩФмЛсМѕЩйЁЃОЁЙмШчДЫЃЌдкИаШОЦкМфДцдкЧьДѓУЙЫиЪБЃЌФкЛЏЕФВЁдЬхЪ§СПШдШЛдіМгЃЌетБэУїM. gallisepticumПЩФмОпгаНјааЯИАћФкИДжЦЕФФмСІЁЃ
жЇдЬхдкЫожїЯИАћФкГЄЦкДцЛюЕФШЗЧаЛњжЦШдШЛВЛЧхГўЁЃНќЦкбаОПЧПЕїСЫгажњгкЯИАћФкДцЛюЕФвђЫиЁЃЬиБ№гаШЄЕФЪЧЃЌдкЫожїЯИАћжаЗЂЯжСЫМзЛљЛЏвРРЕадГСФЌЛњжЦЃЌетЕМжТШыЧжЕФжЇдЬхCGМзЛљЛЏЫЎЦННЕЕЭЁЃИпЖШCGМзЛљЛЏЕФM. hyorhinisБэЯжГіЪмЫ№ЕФЗБжГФмСІЃЌЖјНЯЕЭЕФCGМзЛљЛЏЫЎЦНгаРћгкжЇдЬхдкБЛИаШОЕФЫожїЯИАћФкДцЛюЁЃдкM. gallisepticumжаЩЯЕїSpxAБэДяПЩФмгажњгкЦфбгГЄЯИАћФкЕФДцЛюЪБМфЃЌвђЮЊдкBacillus subtilisжаSpxAБэДяИГгшСЫЖдИїжжбЙСІвђЫиЕФЗРгљФмСІЁЃвђДЫЃЌПЩвдКЯРэМйЩшM. gallisepticumжаSpxAЕФЩЯЕїдкЦфЪЪгІЯИАћФкЛЗОГЙ§ГЬжаЗЂЛгСЫЙиМќзїгУЁЃДЫЭтЃЌMG_186вбБЛШЗЖЈЮЊM. genitaliumЯИАћФкДцЛюЕФвЛИіБивЊвђЫиЃЌвђЮЊЫќдкЫожїжаЕїНкКЫЫсНЕНтвдДйНјжЇдЬхЩњЮяКЯГЩЙІФмКЭИДжЦЁЃ
СэвЛИіЗЂШЫЩюЪЁЕФЪТМўЪЧжЇдЬхФмЙЛШЬЪмГЄЦкЕФЯИАћФкДцЛюЃЌВЂЭЈЙ§ЭтХХзїгУЪЭЗХЕНЯИАћЭтЛЗОГжаЃЌДгЖјЬгБмФкЬхНЕНтКЭздЪЩЁЃетжжВпТддЪаэЯИОњдкЯИАћЭтЛЗОГжавдЛюадзДЬЌЗБжГЃЌДгЖјжїЖЏЧжШыИїжжЫожїЯИАћВЂДйНјЫќУЧЕФЯЕЭГДЋВЅЁЃ
6.2 ЯИАћШыЧжЕФвтвх
ЕБШыЧжЕНЫожїЯИАћКѓЃЌжЇдЬхЖдЫожїЯИАћЕФЩњДцКЭЩњРэЙІФмВњЩњСЫЩюдЖЕФгАЯьЁЃжЕЕУзЂвтЕФЪЧЃЌЫожїЯИАћЮЊжЇдЬхЕФдіжГЬсЙЉСЫвЛИіЬиШЈЩњЬЌЮЛЃЌЪЙЕУетаЉГєУћебжјЕФВЁдЬхЖдПЙЩњЫиЛђУтвпНщЕМЕФЩБЩЫОпгаЯджјЕФЕжПЙСІЁЃОЁЙмдкИаШОЦкМфДцдкВЛЭъШЋЕФУтвпЗДгІЃЌетжжЕжПЙСІШдШЛДцдкЁЃСэвЛЗНУцЃЌФГаЉЯИАћЖОадВњЮяЃЌШчКЫЫсУИЃЌПЩвддкЫожїЯИАћФкв§Ц№DNAЫ№ЩЫЃЌвђЮЊжЇдЬхдкЯИАћФкГжајДцдкЁЃдкжЇдЬхгыЫожїЯИАћжЎМфЕФЯрЛЅзїгУЙ§ГЬжаЃЌГіЯжСЫИїжжЯИАћЩњЮябЇааЮЊЕФБфЛЏЃЌР§ШчвжжЦЕђЭіКЭПЙжзСіЪТМўЃЌЙВЭЌДйГЩСЫгыжЇдЬхЯрЙиМВВЁЕФИДдгЗЂВЁЛњжЦЁЃ
ОЁЙмжЇдЬхКмдчОЭБЛЯШЬьУтвпЯЕЭГЪЖБ№ЃЌДЅЗЂСЫЭЬЪЩЯИАћЕФЭЬЪЩЙ§ГЬЛђTLR4НщЕМЕФФкВПЛЏвдДйНјДйбзЯИАћвђзгЕФЗжУкЃЌЕЋЫожїЯИАћЕФПьЫйШыЧжЖдЧхГ§ЯИАћФкжЇдЬхЙЙГЩСЫОоДѓеЯАЁЃМДЪЙдкНЈСЂгааЇЕФУтвпЗДгІЪБЃЌетвЛЬєеНШдШЛДцдкЃЌОЁЙмУтвпЯЕЭГгаФмСІМѕЩйЯИОњИКЕЃЁЃжЇдЬхДЉЙ§еГФЄЦСеЯВЂНЈСЂЕжПЙУтвпЗДгІКЭПЙЩњЫиИЩдЄЕФРэЯыЩњЬЌЮЛЕФФмСІНјвЛВНдіМгСЫЭъШЋИљГ§ЕФМшОоШЮЮёЁЃДЫЭтЃЌШыЧжЕФжЇдЬхдкЫожїЯИАћФкОРњПЙдБфвьЃЌМгОчСЫПЙИаШОУтвпСІЯћГ§етжжЬгБмадВЁдЬхЕФЬєеНЁЃ
ОЁЙмВЛЭЌЫожїЯИАћЕФжЇдЬхШыЧжДјРДИїжжЗчЯеЃЌвЛаЉжЇдЬхЭЈЙ§ЗжУкгаЖОУИЛђДњаЛВњЮяБэЯжГіеце§ЕФжТВЁадЃЌетаЉВњЮядкЫожїЯИАћФкв§Ц№DNAЛђбѕЛЏгІМЄЫ№ЩЫЁЃжЕЕУзЂвтЕФЪЧЃЌЗжУкадКЫЫсУИШчMG-186ЁЂMpn133КЭMbovNaseЕФЛюадгыКЫDNAНЕНтгаЙиЃЌЖјИЪгЭДњаЛвьГЃЕМжТгаКІЙ§бѕЛЏЮяжждкЯИАћФкЛ§РлЁЃ
ШыЧжКѓЃЌжЇдЬхЭЈЙ§ШХТвЫожїЯИАћЕФдіжГЖЏЬЌв§ШыСЫжиДѓЗчЯеЁЃжЕЕУзЂвтЕФЪЧЃЌU. diversumИаШОЖдЯИАћЩњГЄОпгаЫЋжигАЯьЃЌдчЦкДйЕђЭіЪТМўЙ§ЖЩЕНЭэЦкПЙЕђЭіЪТМўЁЃетаЉИДдгЕФЪТМўПЩФмВПЗжЭЈЙ§ЕїНкNF-ІЪBЛюадНщЕМЃЌЮЊжЇдЬхдкЫожїЯИАћБмФбЫљФкЕФЩњДцЁЂИДжЦКЭДЋВЅЬсЙЉСЫгаРћЛЗОГЁЃЫожїЯИАћдіжГаХКХЕФИФБфЫЦКѕЪЧжЇдЬхжТВЁадЕФВЛПЩЛђШБЕФзщГЩВПЗжЁЃДгЛњжЦЩЯНВЃЌжЇдЬхжаЕФЗжзгАщТТЕААздкетаЉЪТМўжаАчбнживЊНЧЩЋЁЃР§ШчЃЌвЛаЉжЇдЬхБэДяDnaKЃЌвжжЦDNAаоИДЛњжЦВЂвжжЦp53ЕФзЊТМЛюадЁЃетБэУїЫќУЧПЩФмОпгаЧБдкЕФжТАЉЬиадЁЃДЫЭтЃЌжЇдЬхгыЫожїЯИАћжЎМфЕФГЄЦкТ§адЯрЛЅзїгУПЩФмЕМжТЯИАћааЮЊЕФВЛПЩФцБфЛЏЁЃжЕЕУзЂвтЕФЪЧЃЌM. fermentasЁЂM. hominisКЭM. genitaliumвбдкжзСіЮЂЛЗОГжаБЛМьВтЕНЃЌОЁЙмашвЊНјвЛВНбаОПвдШЗЖЈжЇдЬхдкетвЛБГОАЯТЕФШЗЧажТАЉЛюадЁЃ
7 ЮДРДеЙЭћ
ОЁЙмзд1980ФъДњвдРДОЭгаЙигкжЇдЬхЯИАћФкЩњДцЕФжЄОнЃЌЕЋгЩгкжЇдЬхЖдгЊбјЕФбЯИёвЊЧѓКЭШБЗІЯШНјЕФЛљвђБрМЙЄОпЃЌЮвУЧЖджЇдЬхШыЧжЕФИДдгЛњжЦЕФРэНтШдШЛЪмЕНбЯжиЯожЦЁЃеГИНЪЧШыЧжЕФВЛПЩЛђШБЕФЯШОіЬѕМўЁЃШЛЖјЃЌШЗЖЈФФаЉЬиЖЈРраЭЕФеГИНгажњгкГЩЙІШыЧжШдШЛЪЧвЛИіЬєеНЁЃДЫЭтЃЌМДЪЙдкЪЙгУЯрЭЌРраЭКЭЯрЭЌеГИНЙІФмЕФжЇдЬхИаШОвдМАЪЕбщЬѕМўЯТЃЌвВЙлВьЕНЯИАћШыЧжаЇТЪЕФЯджјВювьЁЃетв§ЗЂСЫвЛИіЛљБОЮЪЬтЃЌЮЊЪВУДжЛгавЛаЁВПЗжжЇдЬхФмГЩЙІШыЧжЫожїЯИАћЃЌЖјДѓЖрЪ§дђЪЇАмЃЌетБэУїМШгажЇдЬхвВгаЫожїЯИАћЕФВЮгыЁЃЬНЫїжЇдЬхФкдкЕФЖОСІвђзгКЭЩцМАЫожїЯИАћФЄШыЧжЕФЪмЬхЪЧЮДРДбаОПЕФЙиМќЭООЖЁЃШЛЖјЃЌгЩгкжЇдЬхЛљвђзщЕФОЋМђЬиЕуЃЌЪЖБ№етаЉЖОСІвђзгЬсГіСЫОоДѓЬєеНЃЌЪЙЕУЛљвђБрМБфЕУвьГЃМшФбЁЃОЁЙмзюНќГіЯжЕФЁАГЌЯЁгажизщЪТМўбЁдёЁББрМММЪѕдкЭЦНјжЇдЬхЙІФмЛљвђбаОПЗНУцГфТњЯЃЭћЃЌЕЋгаПЩФмЫцЛњВхШыЕНФПБъЮЛЕуЕФЙІФмадУИЃЈGP35КЭCreЃЉПЩФмВЛЛсЭъШЋГСФЌЛљвђЕФЙІФмЁЃДЫЭтЃЌжизщЬхдкЖрДЮДЋДњКѓЪЧЗёБЃГжЮШЖЈвВДцдкељвщЁЃвђДЫЃЌвЊШЋУцСЫНтПижЦжЇдЬхШыЧжЕФИДдгЛњжЦЃЌШдгаДѓСПЙЄзївЊзіЁЃ
ЦЦвыЩцМАжЇдЬхШыЧжЕФИДдгаХКХЭЈТЗЪЧвЛЯюжиДѓЕФПЦбЇХЌСІЁЃжЕЕУзЂвтЕФЪЧЃЌОЁЙмФкЭЬзїгУИпЖШвРРЕгкЭјИёЕААзКЭНбЕААзЃЌжЌЗЄЁЂЮЂЙмЁЂЮЂЫПвдМАЭЈЙ§ИЪгЭДњаЛВњЩњЕФH2O2вВПЩФмгажњгкжЇдЬхШыЧжЁЃШЛЖјЃЌЙиМќЗжзгЛђМЄУИЕФШЗЧаЩэЗнШдШЛВЛУїЁЃЙЬгаЕФВЛЮШЖЈадЛђПЩжиИДадВюЃЌМгЩЯЕЭаЇТЪКЭТўГЄЕФжЇдЬхШыЧжжмЦкЃЌЮЊЦЦвыШыЧжБГКѓЕФИДдгаХКХЭЈТЗЙЙГЩСЫОоДѓеЯАЁЃДЫЭтЃЌжЇдЬхдкШыЧжКѓШчКЮЗЕЛиЕНЯИАћЭтПеМфЕФЛњжЦШдШЛВЛЧхГўЁЃЦљНёЮЊжЙЃЌбаОПжївЊМЏжадкУшЪіКЭБэеїИУЩњЮяЬхЃЌвђЮЊжЇдЬхЕФЛКТ§ЩњГЄКЭгаЯоЕФКЯГЩФмСІЮЊЯЕЭГбаОПДјРДСЫЪЕжЪадЬєеНЁЃ
жЇдЬхШыЧжЕФЩњРэвтвхБЛЙуЗКШЯЮЊЪЧЬгБмЫожїУтвпгІД№ЕФВпТдЁЃдкЫожїЯИАћФкНЈСЂБмФбЫљЃЌжЇдЬхНЈСЂСЫвЛИіздШЛБгЛЄЫљЃЌБЃЛЄЫќУЧУтЪмУтвпЧхГ§ЃЌЪЙЕУДЋЭГПЙЩњЫижЮСЦБфЕУРЇФбЁЃДЫЭтЃЌетжжШыЧждкТ§адИаШОжаЗЂЛгЙиМќзїгУЃЌЬиБ№ЪЧдкУкФђЩњжГЕРФкЁЃШЛЖјЃЌЮЪЬтШдШЛДцдкЃЌМДЮЊЪВУДЯИАћФкЕФжЇдЬхФбвдБЛЯИАћФкФЃЪНЪЖБ№ЪмЬхЪЖБ№ЃЌзшАСЫгааЇУтвпгІД№ЕФЦєЖЏЃЌетЪЧдкЦфЫћЯИАћФкМФЩњГцжаЙлВьЕНЕФЯжЯѓЁЃзЊТМзщЗжЮігаЯЃЭћНвЪОгаМћЕиЕФЯпЫїЃЛШЛЖјЃЌНЈСЂвЛИізМШЗФЃФтМђЕЅЯИАћФкжЇдЬхИаШОЕФЧПДѓЯИАћФЃаЭДњБэСЫСэвЛИіЯрЕБДѓЕФЬєеНЁЃзмжЎЃЌЬНЫїжЇдЬхгыецКЫЫожїЯИАћдкЯИОњШыЧжЁЂЯИАћФкЩњДцКЭЙуЗКДЋВЅЕФИїИіНзЖЮЕФЯрЛЅзїгУНвЪОСЫжЊЪЖЕФЙиМќЕуЁЃдкетИіСьгђФкЃЌВћУїПижЦжЇдЬхЩјЭИШыЫожїЯИАћЕФЩёУиЛњжЦЃЌжИЕМЦфКѓајТЗОЖЁЃЭЈЙ§гІгУЙІФмЛљвђзщбЇЃЌАќРЈОЋШЗЕФвХДЋВйзїЙиМќЛљвђЛђдкФПБъЫожїЯИАћжаПижЦЙиМќЛљвђЕФБэДяЃЌЪЖБ№гаЯЃЭћЕФжЮСЦАаБъКЭвпУчКђбЁепБфЕУдНРДдНПЩааЁЃвђДЫЃЌетЮЊПЊЗЂгааЇЕФЬцДњКЭ/ЛђИЈжњжЮСЦЗНЗЈКЭдЄЗРВпТдПЊБйСЫЭООЖЁЃ
https://blog.sciencenet.cn/blog-41174-1431606.html
ЩЯвЛЦЊЃКЪГЮядЊЫижмЦкБэМЦЛЎ
ЯТвЛЦЊЃКВЛЭЌРДдДЧтЗжзгздгЩЛљЧхГ§ЛюадБШНЯЁО22еТЁП
