ВЉЮФ
ДѓЦјЮЂСПЦјЬхЕФЮЂЩњЮябѕЛЏ
|
етЪЧвЛЦЊЙигкРћгУбѕЛЏДѓЦјЮЂСПЦјЬхДцЛюЕФЮЂЩњЮяЯрЙиЕФзлЪіЁЃЮвУЧжиЕуашвЊбЇЯАеЦЮеЕФжЊЪЖЪЧЃЌвРПППеЦјжаКлСПЧтЦјЩњДцЕФЯИОњЪЧЕиЧђећИіЩњЬЌЯЕЭГЕФзюЛљДЁВПЗжЃЌПЩвдЫЕУЛгаетжжППГдЧтЦјајУќЕФДѓСПЯИОњЃЌЮоЗЈЮЌГжЮЂЩњЬЌЯЕЭГЃЌУЛгаЮЂЩњЬЌОЭВЛЛсгаНЁПЕЕФЩњЬЌЯЕЭГЃЌвВОЭвтЮЖзХЕиЧђЩњЮяФбвдЮЌГжЩњДцЁЃЕиЧђДѓЦјжаЧтЦјКЌСПЮЊ531ppbЃЌЛђ0.531ppmЃЌетЪЧМАЦфЯЁЩйЕФХЈЖШЃЌаэЖрМьВтЗНЗЈЖМЮоЗЈМьВтЕНЁЃЕЋЪЧећИіЕиЧђЩЯЮЂЩњЮяУПФъОЙШЛЯћКФTgМЖБ№ЃЌTЪЧЭђвкЪ§СПМЖЃЌетЪЧВЛФмКіЪгЕФЙцФЃЃЌвтЮЖзХФмВњЩњДѓСПФмСПКЭИќДѓСПЕФгаЛњЮяЁЃ
ЮвУЧвЛАуЖдбѕЦјЖўбѕЛЏЬМЕФЩњУќЕиЮЛБШНЯжиЪгЃЌЕЋЪЧПЩФмПеЦјжаМАЦфЯЁЩйЕФЛЙдадЦјЬхЖдгкЕиЧђЩњЬЌЛЗОГЕФЕиЮЛИќживЊЁЃЧтЦјЪЧЕиЧђЩњУќНјЛЏЕФЛљДЁЃЌПЩвдЫЕУЛгаЧтЦјОЭУЛгаЕиЧђЩњУќЁЃЖјИљОнзюаТбаОПЗЂЯжЃЌФПЧАЕиЧђДѓЦјжаЕФКлСПЧтЦјШдШЛвЛжБЮЌГжзХЕиЧђЩњЬЌЯЕЭГЕФЛљДЁЃЌЪЧЕиЧђЩњУќЮЌГжЕФИљБОЁЃЫљвдПЩвдЫЕЃЌУЛгаЧтЦјОЭУЛгаЩњЬЌЯЕЭГЁЃЧтЦјЖдЩњУќРДЫЕЃЌЪЧШЋЗНЮЛКЧЛЄЁЃ
ШчЙћвЛЖЈвЊШЫРрИааЛвЛИіЮяжЪЃЌЧтЦјОЭЪЧЮвУЧЪзЯШвЊбЁдёЕФЁЃ
дФЖСБОЮФШУЮвУЧздМКСЫНтМИИіИХФюЃЌЙигкДњаЛЧтЦјЕШПеЦјжаКлСПЦјЬхЕФЯИОњЃЌЪЧвЛИіЗЧГЃаТЕФбаОПСьгђЃЌЙ§ШЅШЫУЧЖдетаЉЯИОњКмВЛСЫНтЃЌЩѕжСНёЬьЖМВЛЧхГўетаЉЯИОњШчКЮРћгУетаЉКЌСПЗЧГЃЕЭЕФЦјЬхДцЛюЁЃЕЋЪЧгавЛаЉаХЯЂКмживЊЃЌетаЉЯИОњПЩФмЪЧЕиЧђЩњЬЌЯЕЭГЕФзюЕзВуЃЌвВЪЧзюЛљДЁЕФзщГЩЁЃвВОЭЪЧЫЕЃЌУЛгаетаЉДњаЛЧтЦјЕФЯИОњЃЌЮвУЧНёЬьЕФЕиЧђЩњЬЌЯЕЭГПЩФмЛсзпЯђБРРЃЁЃетВЂВЛЪЧЮЃбдЫЪЬ§ЁЃвђДЫСЫНтетЗНУцаХЯЂЪЧУПИіЖдЩњЮявНбЇЯрЙиСьгђгааЫШЄЕФЖСепЖМгІИУСЫНтЕФаТНјеЙЁЃ
![]() ДѓЦјЛЙдЦјЬхЪЧЕиЧђЩњЬЌЕФЛљДЁвђЫизлЪі.pdf
ДѓЦјЛЙдЦјЬхЪЧЕиЧђЩњЬЌЕФЛљДЁвђЫизлЪі.pdf
Greening C, Grinter R. Microbial oxidation of atmospheric trace gases. Nat Rev Microbiol. 2022 Sep;20(9):513-528. doi: 10.1038/s41579-022-00724-x. Epub 2022 Apr 12. PMID: 35414013.
ДѓЦјзюНќБЛШЯЮЊЪЧЮЌГжЩњУќЕФжївЊФмдДЁЃЖржжашбѕЯИОњбѕЛЏДѓЦјжаШ§жжзюЗсИЛЕФЛЙдКлСПЦјЬхЃЌМДЧтЃЈH2ЃЉЁЂвЛбѕЛЏЬМ ЃЈCOЃЉ КЭМзЭщ ЃЈCH4).БОЮФзлЪіСЫетаЉДѓЦјКлСПЦјЬхЮЂЩњЮябѕЛЏЕФЗжРрЗжВМЁЂЩњРэзїгУКЭЩњЛЏЛљДЁЃЌвдМАетвЛЙ§ГЬдкЩњЬЌЁЂЛЗОГЁЂвНбЇКЭЬьЬхЩњЮябЇЗНУцЕФживЊадЁЃДѓЖрЪ§ЭСШРЯИОњКЭвЛаЉЙХОњПЩвдЭЈЙ§ЪЙгУДѓЦјжаЕФH2КЭвЛбѕЛЏЬМзїЮЊЬцДњФмдДЃЌШчЗжжІИЫОњЯИАћКЭСДУЙОњцпзгЕФвХДЋбаОПЫљЪОЁЃФГаЉзЈвЕЯИОњвВПЩвдНідкПеЦјжаЩњГЄЃЌе§ШчMethylocapsa gorgonaЕФБъжОадЬиеїЫљжЄЪЕЕФФЧбљЃЌMethylocapsa gorgonaЭЈЙ§ЭЌЪБЯћКФДѓЦјCHРДЩњГЄ4ЃЌ H2ЯИОњЪЙгУН№ЪєУИЕФИпЧзКЭСІЦзЯЕЃЌМДЧтЛЏУИЁЂвЛбѕЛЏЬМЭбЧтУИКЭМзЭщЕЅМгбѕУИЃЌРћгУДѓЦјжаЕФКлСПЦјЬхНјаагабѕКєЮќКЭЙЬЬМЁЃИќЙуЗКЕиЫЕЃЌКлСПЦјЬхбѕЛЏОњдіЧПСЫЭСШРКЭКЃбѓЩњЬЌЯЕЭГЕФЩњЮяЖрбљадКЭИДдСІЃЌдкФЯМЋЩГФЎЭСШРЕШМЋЖЫЛЗОГжаЭЦЖЏСЫГѕМЖЩњВњСІЃЌВЂЭЈЙ§МѕЩйЮТЪвЦјЬхКЭгаЖОЮлШОЮяЕФШЫЮЊХХЗХРДЬсЙЉЙиМќЕФМрЙмЗўЮёЁЃ
ДѓЦјзїЮЊбѕЦјЕФжївЊРДдДЖдЩњУќжСЙиживЊЃЈO2ЃЉ гУгкгабѕКєЮќЃЌЕЊЦј ЃЈN2ЃЉ гУгкЙЬЕЊКЭЖўбѕЛЏЬМ ЃЈCO2ЃЉ гУгкГѕМЖЩњВњЁЃШЛЖјЃЌжБЕНЙ§ШЅЪЎФъЃЌДѓЦјВХБЛШЯЮЊЪЧжЇГжВЛЭЌЩњЬЌЯЕЭГЕФживЊФмдДЁЃгЩгкздШЛзЪдДКЭШЫЮЊХХЗХЃЌДѓЦјжаДцдкИїжжКлСПЦјЬхЁЃетаЉЦјЬхжазюЗсИЛЕФЪЧЧтЦјЃЈH2ЃЉЁЂвЛбѕЛЏЬМ ЃЈCOЃЉ КЭМзЭщ ЃЈCH4ЃЉЃЌЦфШЋЧђЦНОљЛьКЯБШР§ЗжБ№ЮЊЪЎвкЗжжЎ 531ЁЂ90 КЭ 1ЃЌ857ЁЃИїжжЭСШРЮЂЩњЮявбОНјЛЏГіИпЧзКЭСІУИРДЯћКФетаЉЦјЬхвджЇГжгабѕКєЮќЃЌгаЪБЛЙжЇГжЬМЙЬЖЈЁЃЫќУЧМгЦ№РДеМДѓдМ 75% ЕФH2ОЛЯћКФСПЃЌ 15% вЛбѕЛЏЬМКЭ 4% CH4УПФъЖМДгДѓЦјжаЯћЪЇЁЃвђДЫЃЌЫќУЧЭЈЙ§МѕЩйЦјКђЛюадКЭгаЖОЦјЬхЕФХХЗХЃЌжДаавЛЯюжївЊЕЋОГЃБЛКіЪгЕФЩњЬЌЯЕЭГЕїНкЗўЮёЁЃБэ1змНсСЫПижЦетШ§жжЦјЬхШЋЧђЩњЮяЕиЧђЛЏбЇбЛЗЕФИїжжРДдДКЭЛуЁЃ
ЫфШЛДѓЦјКлСПЦјЬхбѕЛЏЕФЩњЮяЕиЧђЛЏбЇживЊадвбЕУЕНЙуЗКШЯПЩЃЌЕЋжБЕНзюНќВХжиЪгетвЛЙ§ГЬЕФЩњЬЌживЊадЁЃетЗДгГСЫНщЕМКлСПЦјЬхбѕЛЏЕФЮЂЩњЮяКЭУИвдМАИУЙ§ГЬЕФЩњРэзїгУШдШЛФбвдзНУўЁЃШЫУЧДэЮѓЕиШЯЮЊЮоЯИАћЃЈЁАabionticЁБЃЉУИжївЊНщЕМДѓЦјжаЕФH2бѕЛЏЃЌЖјАБКЭМзЭщЕЅМгбѕУИЕФСЊКЯзїгУЪЧДѓЦјжаCOбѕЛЏЕФдвђЁЃДѓЖрЪ§ДЋЭГЕФбАевКлСПЦјЬхбѕЛЏДњаЛЮЂЩњЮяЃЌвЛАуЪЧЭЈЙ§ЪЙгУКЌгаИпЫЎЦНH2ЃЈФмдДЃЉЕФбЁдёадХрбјЛљРДЗжРыЃЈКЌЖўбѕЛЏЬМЃЉКЭCOЃЈЬМдДЃЉЃЌетжжЗНЗЈжЛФмЛёЕУЕЭЧзКЭСІЕФЧтгЊбјОњКЭєШЫсЬМгЊбјОњЃЌвђЮЊетжжХрбјЛЗОГЪЪКЯетаЉЯИОњЕФЩњГЄЃЌгЩгкЕижЪЛђЩњЮяЙ§ГЬЃЌетаЉЮЂЩњЮяФмдкИпХЈЖШЕФЛЗОГжаЩњГЄЁЃетвЛДэЮѓЕМжТШЫУЧвЛжБШЯЮЊЮЂЩњЮядкздШЛДѓЦјХЈЖШЯТЮоЗЈгааЇЕиЪЙгУетаЉКлСПЦјЬхЁЃ
ВЛЙ§ЃЌНќФъРДбаОПЗЂЯжаэЖрЯИОњПЩвдЯћКФДѓЦјжаЕФКлСПЦјЬхЁЃетжжЯИОњЕФИпЧзКЭСІЮќЪеЖЏСІбЇгыећИіЭСШРдкЬхЭтКЭдЮЛЩЯЯрЕБЃЌетБэУїЫќУЧНтЪЭСЫетаЉЦјЬхЕФЭСШРЯћКФЁЃгыДЫЭЌЪБЃЌдкЪЖБ№КЭБэеїбѕЛЏДѓЦјCH4ЕФЪШМзЭщОњЗНУцвВШЁЕУСЫКмДѓНјеЙЁЃ
ИУСьгђЕФЭЛЦЦЪЧШЯЪЖЕНИпЧзКЭСІКлСПЦјЬхбѕЛЏОњВЩгУЛьКЯгЊбјЃЈдгЪГадЮЂЩњЮяЃПЃЉЕФЩњЛюЗНЪНЁЃДѓЖрЪ§ВЛЪЧдкЕЅвЛЦјЬхЩЯЩњГЄЃЌЖјЪЧдкЩњГЄЛђЩњДцЙ§ГЬжаНЋДѓЦјКлСПЦјЬхгыЦфЫћгаЛњКЭ/ЛђЮоЛњФмдДЙВЭЌбѕЛЏЁЃжївЊбаОПБэУїЃЌОЙ§ГфЗжбаОПЕФгаЛњвьбјЗжРыжъЃЌШчАќЦЄЗжжІИЫОњЃЌ мПЯуЪєЃЌ АЂЮЌУзЕйРћСДУЙОњКЭШШЮЂЩњЮяУЕЙхЃЌЖМФмЪЙгУДѓЦјH2КЭ/Лђ CO ДцЛюЕБНіЯогкгаЛњЩњГЄЛљжЪЪБЁЃШчЧАЫљЪіЃЌетаЉКлСПЦјЬхЪЧЮЂЩњЮяГЄЦкЩњДцЕФИпЖШПЩППЕФФмдДЃЌвђЮЊЫќУЧдкДѓЦјЛђИпХЈЖШЯТЮоДІВЛдкЃЌЮоашжїЖЏдЫЪфМДПЩЧсЫЩРЉЩЂЕНЯИАћжаЃЌПЩвдБЛзЈУХЕФН№ЪєУИбѕЛЏЃЌЙ§ЕчЮЛзюаЁЃЌЕБгУзїЕчзгЙЉЬхЪБЛсВњЩњДѓСПФмСПгУгкгабѕКєЮќЁЃзюНќЃЌФГаЉЯИОњвбБЛжЄУїЪЧЭЈЙ§гыЦфЫћЛЏКЯЮяЙВЭЌбѕЛЏКлСПЦјЬхЖјЩњГЄЕФЁЃетдкаТЕФЗжРыЮяMethylocapsa gorgonaжаЕУЕНСЫзюКУЕФЗДгГЃЌЫќЭЈЙ§ЭЌЪБЙВбѕЛЏДѓЦјCH4ЃЌ H2КЭвЛбѕЛЏЬМдкПеЦјжаЩњГЄЁЃ
ЮЂЩњЮяПЩвдЁАЩњЛюдкПеЦјжаЁБЕФЗЂЯжжиаТЖЈвхСЫЖдЩњУќЕФзюЕЭвЊЧѓЃЌВЂОпгаЙуЗКЕФгАЯьЁЃдНРДдНЖрЕФжЄОнБэУїЃЌЮЂСПЦјЬхбѕЛЏОњдкШЋЧђЭСШРжаЗсИЛЁЂЖрбљЧвЛюдОЃЌВЂЧвЪЧМЋЕиКЭЩГФЎЭСШРЕШМЋЖЫЛЗОГжаЕФжївЊГѕМЖЩњВњепЁЃДЫЭтЃЌетаЉЯИОњДцдкгкЦиЦјЫЎЩњЩњЬЌЯЕЭГжаЁЂгыЖЏжВЮяЯрЙиЕФЩчЧјКЭНЈжўЛЗОГЁЃБОЮФзлЪіСЫФПЧАЖдДѓЦјЮЂСПЦјЬхбѕЛЏЮЂЩњЮяНщжЪЁЂЩњЛЏЛљДЁЁЂЩњРэзїгУКЭЩњЬЌЯЕЭГКѓЙћЕФШЯЪЖЃЌВЂШЗЖЈСЫетвЛаТСьгђЕФЧАеАадбаОПЗНЯђЁЃЮвУЧзЈзЂгкШ§жжзюЗсИЛЕФЛЙдДѓЦјЦјЬхЃЌH2ЁЂCO КЭ CH4ЃЌОЁЙмШчЗНПђ1ЫљЪіЃЌЮЂЩњЮявВПЩвдбѕЛЏИїжжЦфЫћДѓЦјжаЕФКлСПЦјЬхКЭЛгЗЂадгаЛњЛЏКЯЮяЃЈР§ШчвьЮьЖўЯЉКЭєЪЛљСђЛЏЮяЃЈCOSЃЉЃЉЁЃ
зЈРИ1 ЦфЫћБЛЮЂЩњЮябѕЛЏЕФКлСПЦјЬх
БОЮФжївЊбаОПДѓЦјЧтЃЈH2ЃЉЁЂвЛбѕЛЏЬМ ЃЈCOЃЉ КЭМзЭщ ЃЈCH4ЃЉЃЌвђЮЊетаЉЪЧЦљНёЮЊжЙДѓЦјжазюЗсИЛЕФЛЙдЦјЬхЁЃШЛЖјЃЌЮЂЩњЮяПЩвдЪЙгУИїжжЦфЫћДѓЦјКлСПЦјЬхКЭЛгЗЂадгаЛњЛЏКЯЮязїЮЊЕчзгЙЉЬхЁЃетжжЦјЬхЭЈГЃжЛгаЮЂСПЃЈЭЈГЃЮЊЭђвкЗжжЎвЛЃЉЃЌОЁЙмгЩгкЩњЮяРДдДЃЈР§ШчЃЌбиКЃКЃЫЎжаЕФгаЛњСђЛЏКЯЮяЃЌШчЖўМзЛљСђУбЃЉЃЌЫќУЧПЩФмЛсОжВПЩ§ИпЁЃЛђШЫЮЊЩњВњЃЈР§ШчЃЌГЧЪаЛЗОГжаЕФТБЬўЃЌШчТШМзЭщЃЉЁЃвьЮьЖўЯЉзїЮЊзюЗсИЛЕФЩњЮяЛгЗЂадЛЏКЯЮяКЭгыЦјКђЯрЙиЕФЦјЬхгШЦфв§ШЫзЂФПЁЃЫќУПФъжївЊЭЈЙ§жВБЛДѓСПХХЗХЕНДѓЦјжаЃЈдМ500 TgЃЉЃЌВЂЧвЭЈГЃДяЕНЪЎвкЗжжЎвЛЕФЬхЛ§ЫЎЦНЃЌОЁЙмЦфИпЗДгІадвтЮЖзХЫќБЛбИЫйЛЏбЇбѕЛЏЁЃЭСШРЮЂЩњЮяЛЙГфЕБвьЮьЖўЯЉЕФОЛЛуЃЌУПФъЪМжеШчвЛЕиЯћКФДѓЦјжаЫ№ЪЇЕФвьЮьЖўЯЉЕФдМ4%ЁЃвьЮьЖўЯЉНЕНтЯИОњЫЦКѕЙуЗКДцдкгкЭСШРЁЂКгПкКЭвЖВужаЁЃвбОЗжРыГіИїжжЗХЯпИЫОњКЭБфаЮИЫОњЃЌЫќУЧвдИпХЈЖШЕФвьЮьЖўЯЉзїЮЊЦфЮЈвЛЕФФмСПКЭЬМдДЩњГЄЁЃетШЁОігквьЮьЖўЯЉЕЅМгбѕУИЕФЛюадЃЌЦфМЄЛювьЮьЖўЯЉЮЊ1ЃЌ2-ЛЗбѕвьЮьЖўЯЉЃЌвдМАМИжжЯТгЮУИЁЃШЛЖјЃЌЦљНёЮЊжЙЃЌЛЙУЛгаЮЂЩњЮяБЈИцЪЙгУДѓЦјжаетжжЦјЬхЕФЫЎЦНЁЃзюНќЕФвЛЦЊзлЪіЯъЯИНщЩмСЫФПЧАЖдвьЮьЖўЯЉбЛЗЕФЩњЮяЛЏбЇЁЂЩњРэбЇКЭЩњЬЌбЇЕФРэНтЁЃЮЂЩњЮявВЕїНкЦфЫћЦјЬхЕФЫЎЦНЁЃР§ШчЃЌЭСШРЪЧєЪЛљСђЛЏЮяЃЈCOSЃЉЕФжївЊЛуЃЌВЂЧввбОЗжРыГіЗжжІИЫОњЃЌМДЪЙдкЛЗОГХЈЖШЃЈЬхЛ§ЮЊЭђвкЗжжЎ530ЃЉЯТвВЛсЯћКФетжжЦјЬхЁЃЭЌбљЃЌдНРДдНЖрЕФжЄОнБэУїТШМзЭщЕФЭСШРЛуЁЃШЛЖјЃЌЮвУЧШБЗІЖдФФаЉЮЂЩњЮяЪЙгУетаЉЦјЬхвдМАЫќУЧШчКЮвдМАЮЊЪВУДетбљзіЕФЯЕЭГСЫНтЁЃМјгкетаЉХЈЖШШчДЫжЎЕЭЃЌетаЉКлСПЦјЬхПЩФмгыаэЖрЦфЫћЛЏКЯЮявЛЦ№ЯћКФЃЌзїЮЊЛьКЯгЊбјЩњГЄКЭЩњДцВпТдЕФвЛВПЗжЁЃ
вЛЁЂЮЂСПЦјЬхбѕЛЏОњ
КлСПЦјЬхбѕЛЏОњдкЗжРрбЇЁЂЩњРэбЇКЭЩњЬЌбЇЩЯОпгаИпЖШЖрбљадЁЃДѓЦјH2КЭCOбѕЛЏЕФвХДЋОіЖЈвђЫиЗжБ№гЩжСЩй22ИіКЭ17ИіВЛЭЌЕФУХБрТыЃЌАќРЈШЋЧђЭСШРКЭКЃбѓжаЕФжївЊУХЁЃШчВЙГфБэ1ЫљЪОЃЌЪЕбщжЄУїРДзд9УХКЭ17ЪєЕФЯИОњЗжРыжъЯћКФДѓЦјH2ЁЃЫќУЧАќРЈдјОБЛШЯЮЊНіЯогкбѕЛЏбЧЯѕЫсбЮЁЂCH4ЛђИДдггаЛњЛЏКЯЮяЕФИїжжЯИОњЁЃДѓЦјжаЕФвЛбѕЛЏЬМбѕЛЏОњжСЩйСЅЪєгк4ИіУХКЭ18ИіЪєЃЌАќРЈРДздЭСШРЁЂКЃбѓКЭЕиШШЕиЧјЕФИїжжЯИОњЗжРыЮяЃЈМћВЙГфБэ1ЃЉЁЃживЊЕФЪЧЃЌетжжадзДЛЙбгЩьЕНДгбЮКўжаЗжРыГіЕФЖржжЪШбЮЙХОњЁЃЯрБШжЎЯТЃЌДѓЦјCH4бѕЛЏЫЦКѕОпгаЯСеЕФЗжРрЗжВМЁЃетжжадзДвбОдкМИИіБфаЮИЫОњЪєжаЪЕбщЙлВьЕНЃЌЬиБ№ЪЧMethylocapsaЃЈИпЕиЭСШРДи-ІСЃЈUSCІСЃЉЃЉЃЌЮДддХрЕФйЄТэЯИОњЦзЯЕUBA7966ЃЈUSCІУЃЉКЭМзЛљцпзгГцЃЌвВгЩМзЛљЧђдхНщЕМКЭМзЛљЮЂЩњЮядкИпМзДМХЈЖШЯТЁЃзюНќШЗЖЈЕФКђбЁЪєMethylotropicumЃЈШШДјИпЕиЭСШРДиЃЈTUSCЃЉЃЉдкGemmatimonadotaУХФквВПЩФмЪЙгУДѓЦјCH4ЁЃЭЈЙ§ЕїЖЏЯрЖдХХЫћадЕФЛљжЪЃЌДѓЦјCH4бѕЛЏОњЪЧЙуЗКЕФЩчЧјГЩдБЃЌЗЂЛгзХЙиМќЕФЩњЮяЕиЧђЛЏбЇКЭЩњЬЌзїгУЁЃШчЗНПђ2ЫљЪіЃЌвЛаЉШЫРрЁЂЖЏЮяКЭжВЮяВЁдЬхвВПЩЯћКФH2вдМАЧБдкЕФДѓЦјХЈЖШЯТЕФвЛбѕЛЏЬМЁЃ
Ш§жжУИЃЈ[NiFe]ЧтЛЏУИЁЂютвРРЕадвЛбѕЛЏЬМЭбЧтУИЃЈMo-CODHЃЉКЭПХСЃМзЭщЕЅМгбѕУИЃЈpMMOЃЉЕФИпЧзКЭСІЦзЯЕДпЛЏДѓЦјH2ЁЂCO КЭ CH4ЕФбѕЛЏЁЃЖрЯюЛљвђзщЁЂвХДЋКЭЩњЛЏжЄОнБэУїЃЌетаЉУИИКд№ВЛЭЌЗжРыЮяжаЕФКлСПЦјЬхбѕЛЏЃЌЙВЭЌЮЌГжЭСШРдЊЫиЕФГСЛ§ЁЃетШ§жжУИЖМЪЧЖрбЧЛљН№ЪєЕААзЃЌашвЊМИИіНсЙЙЁЂИЈжњКЭГЩЪьЛљвђВХФмзщзАКЭГЩЪьЁЃетаЉИпЧзКЭСІУИЃЈKM гІгУГЬађ< 150 nMЃЉвВгаЕЭЧзКЭСІЧзЪєЃЈKM гІгУГЬађ> 500 nMЃЉЃЌдкЮЂСПЦјЬхЫЎЦНЩ§ИпЪБЮЌГжЧтгЊбјОњЃЌєШбјОњКЭЪШМзЭщОњЕФЩњГЄЁЃетаЉУИЕФИпЧзКЭСІКЭЕЭЧзКЭСІЦзЯЕжЎМфЕФЯЕЭГЗЂг§ЙиЯЕдкЭМжаПЩЪгЛЏЁЃЫфШЛетжжУшЪіКмгаАяжњЃЌЕЋгІзЂвтСНИізЂвтЪТЯюЁЃЪзЯШЃЌЦљНёЮЊжЙЛЙУЛгаГЩЙІДПЛЏГіПЩвдбѕЛЏДѓЦјХЈЖШМЖБ№H2ЁЂCO Лђ CH4ЕФУИЁЃЯрЗДЃЌДѓЖрЪ§ЖЏСІбЇВтСПРДздДПХрбјЮяЛђећИіЛЗОГбљЦЗЃЈФмЙЛВтСПБэЙлЕЋВЛЪЧеце§ЕФУИЧзКЭСІЃЉЁЃЦфДЮЃЌЛљгкХрбјКЭЩњЬЌЯЕЭГЙцФЃЕФбаОПдНРДдНЖрЕФжЄОнБэУїЃЌЪТЪЕЩЯЃЌЖдДѓЦјКлСПЦјЬхЕФЧзКЭСІЪЧСЌајЕФЁЃ
 ЭМ1ЃКИКд№ЮЂСПЦјЬхбѕЛЏЕФУИЦзЯЕЁЃ
ЭМ1ЃКИКд№ЮЂСПЦјЬхбѕЛЏЕФУИЦзЯЕЁЃ
ДпЛЏЕк1зщКЭЕк2зщ[NiFe]-ЧтЛЏУИЃЈАќРЈHhyLЃЌHucLЃЌHyoLКЭHylLЃЉЕФДѓбЧЛљЕФЯЕЭГЗЂг§ЪїЃЈaВПЗжЃЉЃЌютвРРЕадвЛбѕЛЏЬМЭбЧтУИЕФДпЛЏДѓбЧЛљЃЈMo-CODH;CoxLЃЉЃЈbВПЗжЃЉКЭПХСЃМзЭщЕЅМгбѕУИЃЈpMMO;PmoAЃЉЃЈcВПЗжЃЉЁЃЭЦЖЯвѕгАЗжжЇПЩбѕЛЏДѓЦјжаЕФКлСПЦјЬхвдЛёШЁЧтЦјЃЈH2ЃЉЁЂвЛбѕЛЏЬМ ЃЈCOЃЉ КЭМзЭщ ЃЈCH4ЃЉЃЈМДЃЌЭЈГЃЮЊИпЧзКЭСІЛђжаЕШЧзКЭСІЃЉЃЌБОЮФжаЮДзХЩЋЕФНјЛЏжІЪЪгкЪЙгУРДздЩњЮяЁЂЕижЪЛђШЫЮЊРДдДЕФетаЉЦјЬхЕФЩ§ИпЫЎЦНЃЈМДЃЌЭЈГЃЮЊЕЭЧзКЭСІЃЉЁЃЭЈЙ§зюДѓЫЦШЛЗЈЙЙНЈЕФЪїЪЧжаЕуИљЕФЃЌВЂЧвЭЈЙ§ 50 ИіжиИДНјааздОйЁЃАДбЧзщЃЈ1a-1kЃЌ2aЃЌ2eЃЉБъМЧЕФЧтЛЏУИЃЌбЧзщ1fЃЈHyoЃЉЃЌ1hЃЈHhyЃЉЃЌ1lЃЈHylЃЉКЭ2aЃЈHucЃЉЯдЪОНщЕМДѓЦјH2бѕЛЏЁЃОЁЙметаЉЧтЛЏУИЦзЯЕЭЈГЃОпгаИпЧзКЭСІЃЌЕЋЖдгкCupravidus necatorжаБэеїСМКУЕФHhyУИРДЫЕЃЌЧщПіВЂЗЧШчДЫЁЃMo-CODHsАДЗжжЇЛЎЗжЃЌМДЗХЯпОњЃЈAЃЉЁЂбЮИЫОњЃЈHЃЉЁЂБфаЮИЫОњЃЈPЃЉЁЂЛьКЯ1ЃЈM1ЃЉКЭЛьКЯ2ЃЈM2ЃЉЗжжЇЁЃpMMOЕФИпЧзКЭСІИпЧзКЭСІИпЕиЭСШРДиІСЃЈUSCІСЃЉЁЂUSCІУКЭМзЛљцпзгГцЦзЯЕвВЕУЕНСЫЧПЕїЁЃTUSCЃЌШШДјИпЕиЭСШРДиЁЃ
ДѓЦјH2бѕЛЏЫЦКѕдкМИИіГЁКЯЖРСЂНјЛЏЁЃЯЕЭГЗЂг§ЗжЮівЛжТЕиБэУїЃЌЪЪгІЛЗОГЬѕМўЕФ [NiFe]-ЧтЛЏУИЕФжСЩйЫФИіИпЧзКЭСІЁЂЖдбѕВЛУєИаЕФЧтЛЏУИбЧШКЪЧДгНщЕМШБбѕЩњГЄЕФИќЩюЗжжЇЕФЕЭЧзКЭСІЁЂбѕУєИаЧтЛЏУИзцЯШНјЛЏЖјРДЕФЃЌH 2-ЗсИЛЕФЛЗОГЃЈЭМЁЃетаЉЪЧ1hЃЌ1aЃЌ2fКЭ1l [NiFe]-ЧтЛЏУИзщЃЌЗжБ№УќУћЮЊHhyЃЌHucЃЌHyoКЭHylЃЌУПИіЖМЗЧГЃвЃдЖЃЈДѓдМ1-25%ЕФађСаЭЌвЛадЃЉЃЌВЂЧвдквХДЋзщжЏЩЯДцдкЯдзХВювьЁЃЫфШЛЮЊЪВУДИпЧзКЭСІЧтЛЏУИЖрДЮНјЛЏЛЙгаД§НтОіЃЌЕЋетаЉУИЫЦКѕдкЯИОњЩњГЄКЭДцЛюЗНУцОпгаВЛЭЌЕФЙІФмЁЃР§ШчЃЌШчЯТЫљЪіЃЌАќЦЄЗжжІИЫОњБрТыЕФHhyКЭHucУИдкЖЏСІбЇЬиеїЃЌБэДяЦзКЭећКЯЕНКєЮќСДжагаЫљВЛЭЌЁЃИпЧзКЭСІЧтЛЏУИЕФЫФИібЧШКЫЦКѕвбЙуЗКЫЎЦНВЅЩЂЕНВЛЭЌЕФЯИОњУХЁЃЯрБШжЎЯТЃЌДѓЦјжаЕФвЛбѕЛЏЬМЫЦКѕЪЧMo-CODHЕФвЛАуКЭзцЯШЬиеїЁЃБрТыMo-CODHsЕФДѓЦјCOбѕЛЏОњПчдНЫљгаЮхИіжївЊЕФЯЕЭГЗЂг§ЗжжЇЃЈЭМ1b;МћВЙГфБэ1ЃЉЁЃДЫЭтЃЌМДЪЙЪЧєШЫсОњЃЌШчAfipia carboxidovoransЃЈвдЧАГЦЮЊOligotropha carboxidovoransЃЉКЭCarbonactinospora thermoautoautotrophicaЃЈвдЧАГЦЮЊStreptomyces thermoautotrophicusЃЉвВПЩвдбѕЛЏДѓЦјжаЕФCOЁЃШЛЖјЃЌЯИОњЫЦКѕвбОЭЈЙ§ЩаЮДНтОіЕФЛњжЦЕїНкСЫЦфMo-CODHЕФЖЏСІбЇЃЌетШЁОігкЫќУЧЫљНгДЅЕФвЛбѕЛЏЬМЫЎЦНЃЛР§ШчЃЌЭСШРвьбјОњAminobacter sp. COX1БэДяИпЕУЖрЕФЧзКЭСІУИЃЈKM гІгУГЬађ= 12 nMЃЉБШЗЯЫЎздбјОњA.ЬМбѕЛЏЮяЃЈKM гІгУГЬађ= 640 nMЃЉЁЃЙигкДѓЦјCH4бѕЛЏЁЂИпЧзКЭСІЕФpMMOЦзЯЕвбОдкІСБфаЮИЫОњКЭІУТэЯИОњЪШМзЭщОњжаНјЛЏЃЌПЩФмРДздЕЭЧзКЭСІЕФзцЯШЁЃгыMo-CODHРрЫЦЃЌpMMOЕФИпЧзКЭСІКЭЕЭЧзКЭСІБфЬхжЎМфДцдкЯрЖдНЯаЁЕФађСаВювьЁЃ
зЈРИ2 ВЁдЬхжаЕФКлСПЦјЬхбѕЛЏ
ашвЊЩюШыбаОПВХФмГфЗжСЫНтзюНќЗЂЯжЕФЮЂЩњЮяЧхГ§ДѓЦјКлСПЦјЬхЕФвНбЇживЊадЁЃЫфШЛДѓЖрЪ§КлСПЦјЬхбѕЛЏОњНщЕМживЊЕФЩњЬЌЯЕЭГЗўЮёЃЌЕЋвЛаЉШЫРрЁЂЖЏЮяКЭжВЮяВЁдЬхвВПЩФмдкЦфЩњУќжмЦкжаЪЙгУКлСПЦјЬхЁЃР§ШчЃЌЩќаѓВЁдЬхЛєИёКьЧђОњЃЈвдЧАГЦЮЊТэДЬКьЧђОњЃЉКЭжВЮяВЁдЬхНъДЏСДУЙОњЃЈТэСхЪэКкаЧВЁЃЉЖМЯћКФЧтЦјЁЃетвЛЙ§ГЬПЩФмгажњгкЗЂВЁЛњжЦЃЌЕЋИќгаПЩФмЪЙЛњЛсадВЁдЬхдкЛЗОГПтжаГжајДцдкЁЃе§ШчзюНќЩѓВщЕФФЧбљЃЌаэЖрЦфЫћВЁдЬхашвЊ H2ЮЌГжЖОСІЃЌР§ШчгФУХТнИЫОњКЭГІЕРЩГУХЪЯОњбЊЧхЩГУХЪЯОњЪѓЩЫКЎбЧжжЃЌОЁЙмЫќУЧЪЙгУСЫЪЪгІЮИГІЕРЗЂНЭЙ§ГЬжаВњЩњИпЫЎЦН H2 ЕФЕЭЧзКЭСІЧтЛЏУИЁЃ
ЛйУ№адЕФШЫРрВЁдЬхНсКЫЗжжІИЫОњБЃСєСЫЦфЛЗОГзцЯШЕФютвРРЕадвЛбѕЛЏЬМЭбЧтУИЃЈMo-CODHЃЉЃЌЕЋЪЇШЅСЫЧтЛЏУИЃЈетРяЧПЖШЪЇШЅЫЕУїНсКЫИЫОњБОРДгаЧтЛЏУИЃЉЁЃШЫУЧКмШнвзЭЦВтЃЌвЛбѕЛЏЬМЃЈCOЃЉПЩФмЪЧвЛжжЙиМќЕФФмСПРДдДЃЌЪЙетжжГєУћебжјЕФГжОУадЯИОњФмЙЛЬгБмгЊбјУтвпЁЃетжжВЁдЬхГжајБЉТЖгкРДздШ§ИіжївЊРДдДЕФвЛбѕЛЏЬМЃКЛЗОГКЭЮлШОЕФПеЦјЁЂЯубЬбЬЮэКЭЫожїОоЪЩЯИАћбЊКьЫиМгбѕУИ 1ЁЃгавЛаЉжЄОнБэУїетжжЯИОњЯћКФвЛбѕЛЏЬМЃЌАќРЈдкКлСПХЈЖШЯТЃЌОЁЙметжжУИЕФЩњЛЏЛюадКЭЩњРэзїгУШдгаД§УїШЗжЄУїЁЃ
ЖўЁЂЩњРэзїгУКЭећКЯ
2.1 дкЩњДцжаЕФзїгУ
дкздШЛНчжаЃЌДѓЦјH2вЛбѕЛЏЬМЫЦКѕжївЊгЩанУпЯИОњЧхГ§ЃЌвдТњзуГЄЦкЩњДцЦкМфЕФЮЌГжФмСПашЧѓЁЃЕБЯожЦЦфЩњГЄЛљжЪЪБЃЌИїжжашбѕгаЛњвьбјЯИОњЕФХрбјЮяБэДяКЭРћгУЦф[NiFe]-ЧтЛЏУИКЭMo-CODHЁЃетБэУїетаЉЯИОњДгдкгаЛњЛЏКЯЮяЩЯЩњГЄзЊБфЮЊЪЙгУКлСПЦјЬхДцЛюЃЌЭЈГЃгыЦфЫћЭтдДЛљжЪКЭЬМДЂБИвЛЦ№ЩњДцЁЃгЋЙтЯдЮЂОЕЯдЪОЃЌ S. avermitilisбѕЛЏОњЕФHhyзЊТМБОНідкДѓЦјH2ЕФцпзгжаБэДяЁЃШБЗІБрТыетжжЧтЛЏУИЕФНсЙЙЛљвђЕФЭЛБфОњжъЕФцпзгЛюСІНЕЕЭСЫ76%ЁЃдкЗЧцпзгЗХЯпИЫОњАќЦЄЗжжІИЫОњжавВШЁЕУСЫРрЫЦЕФЗЂЯжЃЈЭМ2aЃЉ. дкМЂЖіЦкМфЃЌетжжЯИОњБэДяБрТыHhyЃЌHucКЭMo-CODHЕФЛљвђвдЯћКФДѓЦјжаЕФH2КЭ CO зїЮЊгабѕКєЮќЕФЕчзгЙЉЬхЁЃШБЗІетаЉЛљвђЕФОњжъдкХњСПХрбјКЭСЌајХрбјжаЕФЛюСІЖМбЯжиНЕЕЭЁЃРэТлМЦЫувЛжТБэУїЃЌДѓЦјH2КЭCOбѕЛЏПЩвдвЛЦ№ЮЌГжУППЫЭСШР107ЈC108ЯИАћЪ§ЕФЮЌЛЄвЊЧѓЃЛМјгкЮЌГжФмСПМЦЫуЦЋЯђгкЪЪгІИДжЦгЊбјЛЗОГЕФПьЫйЩњГЄЯИОњЃЌетПЩФмБЛЕЭЙРЁЃ
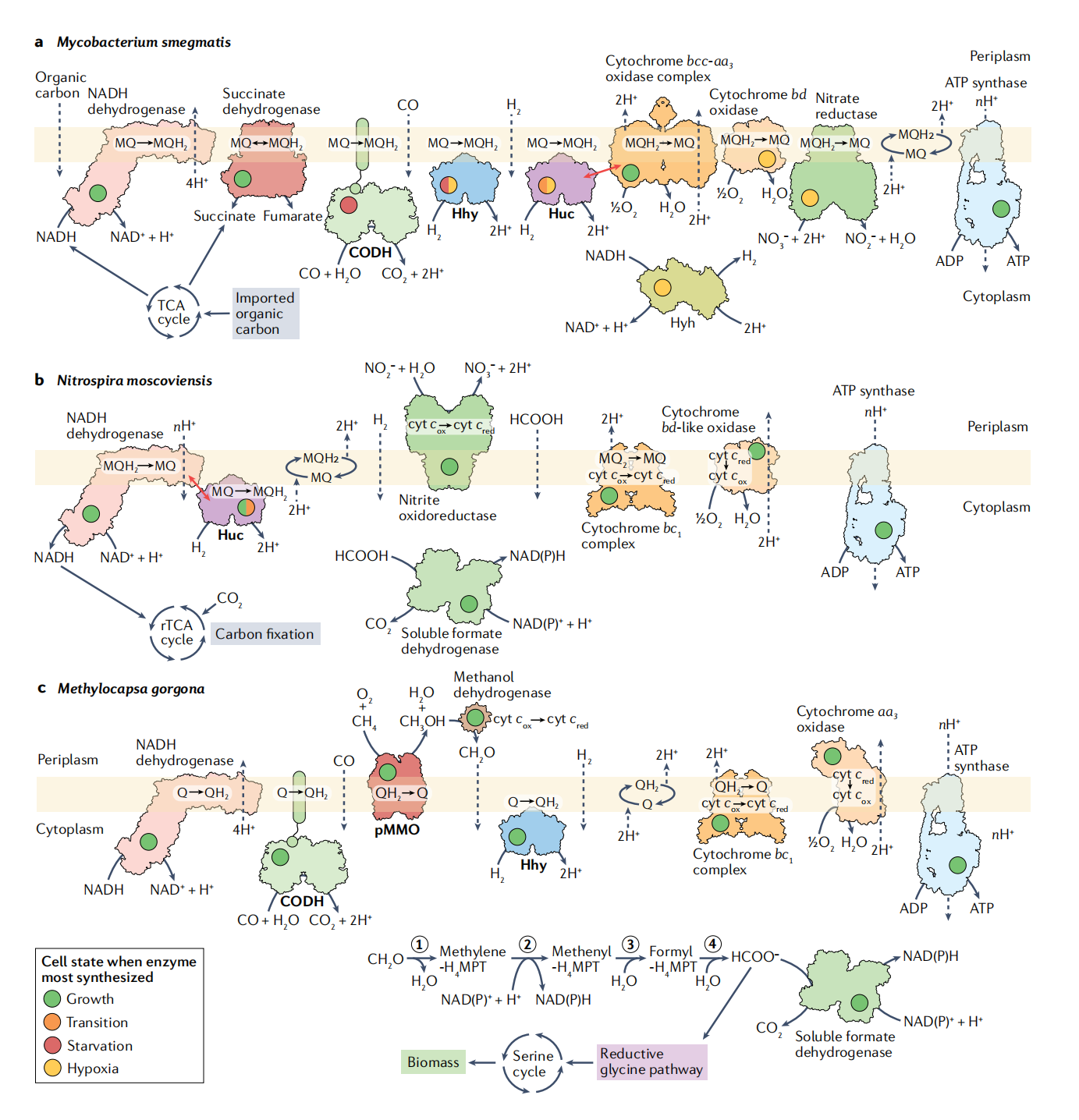
ЭМ2ЃКЮЂСПЦјЬхбѕЛЏЖдЯИОњЩњГЄКЭДцЛюЙ§ГЬжаНкФмКЭЬМЭЌЛЏЙ§ГЬЕФЙБЯзЁЃ
АќЦЄЗжжІИЫОњЕФМђЛЏКєЮќСДЃЌЯдЪОДѓЦјЧтЃЈH2ЃЉКЭHucЃЌHhyКЭютвРРЕадвЛбѕЛЏЬМЭбЧтУИЃЈMo-CODHЃЉЕФвЛбѕЛЏЬМЃЈCOЃЉжЇГжМЂЖіКЭШБбѕЦкМфЕФДцЛюЁЃдкДгЩњГЄЕНГжОУадЕФЙ§ЖЩЙ§ГЬжаЃЌHucВњЩњМзнСрДМЃЌЦфНіБЛЯИАћЩЋЫиbcc-aaбѕЛЏ3бѕЛЏУИИДКЯЮяЃЌВњЩњжЪзгЖЏСІЁЃдкГЄЪБМфЕФгаЛњЬММЂЖіЦкМфЃЌHhyКЭMo-CODHВњЩњрДМЃЌЦфБЛЯИАћЩЋЫиbcc-aaбѕЛЏ3бѕЛЏУИИДКЯЮяЛђЯИАћЩЋЫиBDбѕЛЏУИЁЃдкШБбѕЦкМфЃЌHyhЭЈЙ§ВњЩњH2ЭЈЙ§бѕЛЏNADHЁЃШБбѕЪБЃЌФкдДадЛђЭтдДадВњЩњ H2ПЩБЛHucЛђHhyбѕЛЏЃЌЩњГЩМзнСрДМЃЌМзнСрДМБЛИпЧзКЭСІЯИАћЩЋЫиbdбѕЛЏУИРћгУКлСПбѕбѕЛЏЃЈO2ЃЉЁЂЪЙгУИЛТэЫсбЮЕФчњчъЫсЭбЧтУИ/ИЛТэЫсЛЙдУИЃЌвдМАЪЙгУNOЕФЯѕЫсбЮЛЙдУИ3−ШЁОігкУПИіЕчзгЪмЬхЕФПЩгУадЁЃЮЊМђЕЅЦ№МћЃЌЮДЯдЪОМИжжКєЮќЛЙдУИЁЃФЊЫЙПЦбЧЯѕЛљТна§ЬхКєЮќСДЕФМђЛЏБэЪОЃЌЯдЪОСЫДѓЦјHЕФбѕЛЏЙ§ГЬ2Huc жЇГжбЧЯѕЫсбЮГфТњЬѕМўЯТЕФЩњГЄКЭбЧЯѕЫсбЮЯћКФЬѕМўЯТЕФДцЛюЁЃБЛHucЛЙдЕФМзнСѕЋБЛIаЭNADHЭбЧтУИРћгУЃЌЭЈЙ§ЗДЯђЕчзгСїВњЩњNADHЃЌДгЖјЭЈЙ§ЗДЯђШ§єШЫсЃЈrTCAЃЉбЛЗЪЕЯжЬМЙЬЖЈЁЃc |Methylocapsa gorgonaКєЮќСДЕФМђЛЏБэЪОЃЌЯдЪОСЫДѓЦјHШчКЮЭЌЪБбѕЛЏ2ЁЂвЛбѕЛЏЬМКЭМзЭщЃЈCH4ЃЉ жЇГждіГЄЁЃДѓЦјHЕФбѕЛЏ2HhyКЭMo-CODHЕФвЛбѕЛЏЬМПЩФмЛсМѕЩйѕЋЃЌШЛКѓБЛжЪзгвзЮЛЯИАћЩЋЫиbcбѕЛЏ1ИДКЯЮяКЭЯИАћЩЋЫи AA3бѕЛЏУИЁЃO2-CHЕФвРРЕадКЭрХЕвРРЕадбѕЛЏ4ЭЈЙ§ПХСЃМзЭщЕЅМгбѕУИЃЈpMMOЃЉВњЩњМзДМЁЃМзДМБЛжмжЪМзДМЭбЧтУИбѕЛЏЮЊМзШЉЃЌЛЙдПЩБЛЯИАћЩЋЫиaaдйбѕЛЏЕФЯИАћЩЋЫиc3бѕЛЏУИЁЃМзШЉБЛзЊдЫЕНЯИАћжаВЂвРДЮбѕЛЏГЩМзЫсбЮЃЌШЛКѓбѕЛЏГЩЖўбѕЛЏЬМЃЌВњЩњСНИіNADЃЈPЃЉHЗжзгЁЃ ЛђепЃЌМзЫсбЮЭЈЙ§ЛЙдИЪАБЫсЭООЖКЭЫПАБЫсбЛЗВєШыЩњЮяжЪжаЁЃМзШЉЛюЛЏУИЃЈ1ЃЉЁЂNADЃЈPЃЉвРРЕадбЧМзЛљЫФЧтМзЕћпЪЭбЧтУИЃЈ2ЃЉЁЂбЧМзЛљЫФЧтМзЕћпЪЛЗЫЎНтУИЃЈ3ЃЉКЭМзѕЃзЊвЦУИ/ЫЎНтУИЃЈ4ЃЉгЩгкЦЊЗљЯожЦЖјЮДГщГіЁЃH4MPTЃЌЫФЧтМзЕћпЪЁЃTCAЃЌШ§єШЫсЁЃ
ЪЙгУБфаЮИЫОњЖдГжОУадЯрЙиЕФКлСПЦјЬхбѕЛЏНјааСЫРрЫЦЕФЙлВьЃЌ ЫсИЫОњЃЌ ТЬНцЪєЃЌ ФтИЫОњКЭЯѕЛљТна§ЬхДгИїжжЛЗОГжаЗжРыГіРДЁЃР§ШчЃЌдкЗЧцпзгЯИОњУЕЙхзЖГцКЭдЖЧзцпзгГцThermogemmatispora sp. T2жаЃЌHhyКЭMo-CODHНідкгаЛњЬММЂЖіЦкМфБэДяКЭЛюдОЁЃДЫЭтЃЌОЁЙметСНжжЯИОњЖМСЅЪєгкChloroflexotaУХЃЌЕЋЯЕЭГЗЂг§ЗжЮіБэУїЃЌЫќУЧИїздЕФРрБ№ЖРСЂЕиЫЎЦНЛёЕУСЫКлСПЦјЬхбѕЛЏЕФЛљвђЁЃЃЈЭМ1ЃЉ ОЁЙмМзЭщЕЅМгбѕУИЫЦКѕНіЯогкПЩвддкCH 4ЩЯЩњГЄЕФЯИОњЃЈМДЪШМзЭщОњЃЉЃЌДѓЦјCH4ПЩФмЮЌГжОпгазуЙЛИпЧзКЭСІЕФМзЭщЕЅМгбѕУИЕФЯИОњЕФДцЛювдЯћКФЫќЁЃР§ШчЃЌЛљгкШЋЯИАћЖЏСІбЇЃЌMethylocystis sp. SC2 зщГЩаЭБэДяУїЯдЕФИпЧзКЭСІ pMMO вдбѕЛЏДѓЦј CH4гУгкЮЌГжВЂгеЕМУїЯдЕФЕЭЧзКЭСІ pMMO дк CH 4Щ§ИпЕФЛЗОГжаЩњГЄПЩгУадЃЈЭМ1cЃЉ. РрЫЦЕиЃЌєШЫсєШМЄЫиЃЈШчA. carboxidovoransЃЉбѕЛЏДѓЦјCOЕФФмСІвтЮЖзХМДЪЙЕзЮяЕФПЩгУадВЛзувдЮЌГжЩњГЄЃЌЫќУЧвВОпгаКуЖЈЕФФмСПЙЉгІЁЃзмЬхЖјбдЃЌетаЉвЛжТЕФЙлВьНсЙћКЭКлСПЦјЬхбѕЛЏЛљвђЕФПьЫйДЋВЅБэУїЃЌКлСПЦјЬхбѕЛЏЪЧвЛжжЭЈгУЧвИпЖШПЩППЕФГЄЦкЩњДцВпТдЁЃ
ПижЦДѓЦјКлСПЦјЬхбѕЛЏЕФЕїНкМСНідкАќЦЄЗжжІИЫОњжаНјааСЫбаОПЁЃетжжЯИОњдкЯожЦЩњГЄЫљашЕФгаЛњЬМЛђбѕЪББэДяHucЃЌHhyКЭMo-CODH;HucдкЩњГЄКЭГжОУаджЎМфЕФЙ§ЖЩЦкМфВњЩњВЂЛюдОЃЌЖјHhyЛюаддкГЄЦкГжОУадЦкМфДяЕНЗхжЕ.етШЁОігкДэзлИДдгЕФМрЙмЛњЙЙЭјТчЃЌЦфжаСНИівбОШЗЖЈЁЃАќЦЄЗжжІИЫОњЪЙгУДЋИаЦїЃЈР§ШчИЪгЭгеЕМЕФGylRЃЉжБНгИагІгаЛњЬМЫЎЦНЃЌЕМжТЗжНтДњаЛвжжЦБрТыHucЕФВйзнзгдкЩњГЄЙ§ГЬжагЩЩаЮДШЗЖЈЕФЯьгІЕїНкМСвжжЦЁЃДЫЭтЃЌЯИОњИажЊO2вдМАЪЙгУзщАБЫсМЄУИDosSКЭDosTЕФбѕЛЏЛЙдЫЎЦН;ЕБЕЭЕчЦНЪБЃЌЯьгІЕїНквђзгDosRБЛМЄЛюЃЌЫќЩЯЕїДѓдМ50ИіЛљвђЕФБэДяЃЌАќРЈБрТыHhyКЭH2ЕФВйзнзгВњЩњЗЂНЭЧтЛЏУИЁЃЭЈЙ§етаЉгХбХЕФЛњжЦЃЌЗжжІИЫОњНЋЩњГЄЕзЮяЕФПЩгУадгыГЄЦкЩњДцЫљашЕФУИЕФЩњВњжБНгСЊЯЕЦ№РДЃЈЭМ2aЃЉЁЃЦфЫћЗжНтДњаЛЮяДЋИаКЭбѕЛЏЛЙдДЋИаЯЕЭГПЩФмПижЦЦфЫћЮяжжжаЕФКлСПЦјЬхбѕЛЏЃЌВЂЧввбжЊПижЦЕЭЧзКЭСІЧтгЊбјЩњЮяЃЈР§ШчЃЌCupravidus necator;вдЧАГЦЮЊRalstonia eutropha)ЁЃ
2.2 дкЛьКЯгЊбјЩњГЄжаЕФзїгУ
КмУїЯдЃЌКмЖрЯИОњПЩвдЪЙгУКлСПЦјЬхЛьКЯгЊбјЩњГЄЁЃКлСПЦјЬхбѕЛЏЕФЯИАћЬивьадЫйТЪЭЈГЃЬЋЕЭЃЌЯИОњЮоЗЈдк H2 КЭвЛбѕЛЏЬМCO Лђ CH4зїЮЊЮЈвЛЕФФмдДКЭЬМРДдДЩЯЩњГЄЁЃРэТлЩЯНВЃЌЖдгкЦмЯЂдкЩСжЭСШРжаЕФИпЛюадВњМзЭщОњвдМАМЋЕиЩГФЎжавЛаЉФмСПашЧѓЕЭЕФЯИОњРДЫЕЃЌДѓЦјКлСПЦјЬхЕФЩњГЄЪЧПЩФмЗЧГЃЛКТ§ЕФЁЃШЛЖјЃЌетжжЭЦЖЯЪЧЛљгкЩњЬЌЯЕЭГЙцФЃЕФЙлВьЃЌЦљНёЮЊжЙЛЙУЛгаЙигкПЩвддкДПХрбјЮяжаДѓЦјХЈЖШЕФЕЅвЛКлСПЦјЬхЩЯЩњГЄЕФЯИОњЕФБЈИцЁЃ2019ФъЃЌФмЙЛЁАдкПеЦјжаЩњГЄЁБЕФЯИОњБЛЪзДЮБЈЕРЁЃMethylocapsaЪєжаЕФШ§жжЯИОњЃЌАќРЈРДздUSCІСЗжжЇЕФаТаЭЭСШРЗжРыжъM. gorgonaЃЌдкБЉТЖдкПеЦјжаЕФОлЬМЫсѕЅЙ§ТЫЦїЩЯЩњГЄСЫвЛФъвдЩЯЁЃM. gorgonaЯдШЛЪЧЭЈЙ§ЭЌЪБбѕЛЏДѓЦјCH4ЁЂH2КЭвЛбѕЛЏЬМРДЪЕЯжетвЛЕуЕФ ЃЌвдТњзуЦфЯрЖдНЯЕЭЕФдіГЄФмдДашЧѓЃЈЭМ2cЃЉЁЃШЛЖјЃЌгІИУзЂвтЕФЪЧЃЌЫљгаШ§жжОњжъзюГѕЖМЪЧдкЩ§ИпЕФCH4ЩЯЗжРыЕНЕФЁЃЦљНёЮЊжЙЃЌЛЙУЛгаШЮКЮНіЪЙгУДѓЦјХЈЖШФмСПдДЗжРыГіЕФЯИОњЁЃЪЪгІЭСШРЛЗОГЦфЫћИїжжЪШМзЭщОњКмПЩФмЭЈЙ§ЪЙгУИДдггаЛњЮяЃЈШчДзЫсбЮЃЉКЭДѓЦјКлСПЦјЬхЛьКЯгЊбјЁЃ
ДѓЦјH2ПЩвдВЙГфИїжжЦфЫћгаЛњвьбјЩњЮяКЭбвЪЏздбјЩњЮяЕФЩњГЄЁЃН№ЛЦЩЋЕЅАћОњЁЂТЬЕЏадОлМЏЬхКЭАЂРЫЙМгЧЪФЄЕЅАћОњЗжБ№БэДяHucВЂбѕЛЏДѓЦјH2дкгаЛњвьбјЩњГЄЦкМфЁЃетжжВпТдПЩФмЛсЬсИпЫќУЧдкЬМПнНпЛЗОГжаЕФЬМРћгУаЇТЪЃКЪЙгУH2КЯГЩATPПЩвдНЋИќЖрЕФгаЛњЬМЗжХфИјКЯГЩДњаЛЃЈЩњЮяжЪаЮГЩЃЉЖјВЛЪЧЗжНтДњаЛЃЈATPВњЩњЃЉЁЃШБЗІHucЕФАќЦЄЗжжІИЫОњЭЛБфЬхдкбѕЛЏЬМдДввЫсбЮКЭБћЫсбЮЩЯЕФЩњГЄЫйЖШБШвАЩњаЭТ§ЃЌетСНжжЬМдДЖМашвЊФмСПЮќЪеЁЃетжжЬиадвВбгЩьЕНдкИпЧБСІЛљжЪЃЈШчбЧЯѕЫсбЮЁЂСђЛЏЮяЛђбЧЬњЃЉЩЯбвЪЏздбјЩњГЄЕФЯИОњЁЃР§ШчЃЌдкГфТњбЧЯѕЫсбЮЕФЬѕМўЯТNitrospira moscoviensisзщГЩадЕибѕЛЏДѓЦјжаЕФH2ДйНјЩњГЄЃЌВЂдкбЧЯѕЫсбЮЯћКФЬѕМўЯТМЬајДцЛюЁЃЭЈЙ§ЪЙгУетжжЮоДІВЛдкЕФЕЭЕчЮЛЦјЬхЃЌетжжЯИОњПЩвджївЊЪЙгУбЧЯѕЫсбЮбмЩњЕчзгРДжЇГжЗжНтДњаЛЖјВЛЪЧКЯГЩДњаЛЃЌгЩгкашвЊЗДЯђЕчзгСїЃЌКЯГЩДњаЛГЩБОКмИпЃЈЭМ2bЃЉЁЃ ДгЗЯЫЎДІРэГЇЁЂЮТШЊЁЂБББљбѓКЭЫсадЮВПѓЕШВЛЭЌЛЗОГжаЗжРыГіЩЯЪіЯИОњЃЌБэУїДѓЦјКлСПЦјЬхЩЯЕФЛьКЯгЊбјЩњГЄЪЧвЛжжЛЗОГЙуЗКЕФВпТдЁЃ
2.3НкФмгыЬМЭЌЛЏЙ§ГЬ
дДздДѓЦјH2ЁЂCO КЭ CH4бѕЛЏВњЩњЕФЕчзгжївЊгУгкгабѕКєЮќЁЃгЩH2бѕЛЏВњЩњЕФЕЭЕчЮЛЕчзгЃЈH2Ёњ 2H + 2e+ЈC; EЁфo= ЈC414 mVЃЉ ZКЭвЛбѕЛЏЬМ ЃЈCO + H2O ЁњЖўбѕЛЏЬМ+ 2H + 2e+ЈC; EЁфo= ЈC524 mVЃЉБЛШЯЮЊжБНгзЊвЦЕНКєЮќСДжаЁЃвЛжТЕиЃЌЗжСѓЯИАћЩЯЕФЛюадВтЖЈКЭЕААзжЪгЁМЃЯдЪОHucКЭHhyгыФЄЕФАћжЪВрЯрЙиЃЌMo-CODHвВгыФЄЕФАћжЪВрЫЩЩЂЕиНсКЯЃЌПЩФмЪЧЭЈЙ§дЄВтЕФПчФЄУЊCoxGЁЃЪЙгУВЛЭЌвжжЦМСЕФКєЮќВтЖЈбаОПБэУїЃЌH2-бмЩњКЭCOбмЩњЕФЕчзгБЛзЊвЦЕНМзнСѕЋГиЃЌШЛКѓзЊвЦЕНO2;ШЛЖјЃЌЕчзгЪЧШчКЮДгH2жаМЬЕФЩаЮДНтОіЕФКЭМзнСѕЋЕФвЛбѕЛЏЬМЃЌвдМАЪЧЗёЩцМАЦфЫћбѕЛЏЛЙдЕААзЁЃетаЉЙ§ГЬЖдКєЮќНтёюЦїУєИаЃЌгыВњЩњФЄЕчЮЛЕФЕчзгзЊвЦвЛжТЃЌДгЖјЧ§ЖЏATPКЯГЩЁЃ
ГЃбЙКлСПЦјЬхбѕЛЏЭЈГЃашвЊO2зїЮЊФЉЖЫЕчзгЪмЬх.дкЗжжІИЫОњжаЃЌHucНігыжЪзгвзЮЛХМСЊЯИАћЩЋЫиЛљЕзЯИАћАЉ-AA3бѕЛЏУИИДКЯЮяЃЌПЩФмЭЈЙ§аЮГЩГЌХфКЯЮяЃЌДгЖјзюДѓЛЏУПИіH2ЗжзгвзЮЛЕФжЪзгЪ§СПбѕЛЏ.ЯрБШжЎЯТЃЌHhyКЭMo-CODHгыЯИАћЩЋЫиbcc-aa3ХМСЊбѕЛЏУИИДКЯЮяКЭЗЧжЪзгвзЮЛЯИАћЩЋЫиBDбѕЛЏУИ.ШЛЖјЃЌОнБЈЕРЯИОњЛсбѕЛЏH2дкбЧДѓЦјбЙЫЎЦНЩЯЃЌЪЙгУЯѕЫсбЮКЭИЛТэЫсбЮзїЮЊЕчзгЪмЬхЁЃАќЦЄЗжжІИЫОњдкШБбѕЯТЮЌГжбѕЛЏЛЙдЦНКтЕФжївЊЗНЗЈЪЧЦНКтЗЂНЭH2ЕФЙ§ГЬЩњВњЃЌбсбѕ H2бѕЛЏКЭКУбѕH2бѕЛЏШЁОігкЕчзгЪмЬхЕФПЩгУадЃЈЭМЃЉ2aЃЉЁЃ COбѕЛЏвВПЩвдгыЯѕЫсбЮКЭИпТШЫсбЮЕШЕчзгЪмЬхЕФЛЙдёюКЯЃЌОЁЙмМјгкбѕЛЏЗЂЩњдкНЯИпЕФуажЕХЈЖШЯТЃЌЕЋДѓЦјжаЕФCOЪЧЗёПЩФмгыбсбѕЕчзгЪмЬхбѕЛЏШдгаД§ШЗЖЈЁЃ
вЛаЉДѓЦјH2дЄМЦCOбѕЛЏОњЛсВњЩњCO2ЛЙдМСЙЬЖЈЁЃетШЁОігкдкЯћКФжЪзгЖЏСІЕФЙ§ГЬжаЯђИДКЯЮяIЕФЗДЯђЕчзгСїзЊвЦЃЈЭМ2bЃЉ. гЩДЫВњЩњЕФNADHвбБЛжЄУїжЇГжЖўбѕЛЏЬМЭЈЙ§МгЖћЮФ-БОЩ-АЭЩсФЗбЛЗЛђЗДЯђШ§єШЫсбЛЗЙЬЖЈЁЃЫфШЛH2жаЕФвЛаЉЯИОњ-ЗсИЛЕФБкэшПЩвджБНгёюКЯH2ЭЈЙ§ПЩШмадАћжЪЧтЛЏУИбѕЛЏЮЊNADЃЈPЃЉЛЙд+109ЃЌетаЉУИдкДѓЖрЪ§ДѓЦјH2жаВЛДцдкбѕЛЏОњ51ЃЌ63ЁЊ ПЩФмЗДгГСЫИУЗДгІдкЕЭ H2ЯТдкШШСІбЇЩЯВЛПЩааХЈЖШЁЃШЛЖјЃЌгІИУжИГіЕФЪЧЃЌH2ЛђДѓЦјЫЎЦНЯТЕФвЛбѕЛЏЬМПЩвдЧ§ЖЏЖўбѕЛЏЬМЙЬЖЈдкХрбјЮяЛђЛЗОГбљЦЗжаЁЃДњаЛЫЎЪЧДѓЦјH2ЕФАћжЪИБВњЮябѕЛЏЃЈH2+ 1/2O2Ёњ H2OЃЉ ВЂЧвПЩвддкЕЭЫЎЪЦЕФЛЗОГжаЮЌГжЫЎКЯзїгУЁЃЭЈЙ§етжжМђдМЕФЙ§ГЬЃЌЯИОњПЩвдЭЌЪБДгЁАЯЁБЁЕФПеЦјЁБжажЦдьЯИАћФмСПЃЌЬМКЭЫЎЁЃ
ЖрВНЭООЖПЩЭЈЙ§ДѓЦјCH4ЪЕЯжНкФмКЭЬМЭЌЛЏзїЮЊЛљВФЃЈЭМ2cЃЉ. ЖјЯИОњжБНгЪЙгУДѓЦјжаЕФ H2КЭCOзїЮЊЕчзгЙЉЬхЃЌЯрЖдЖшадЕФCH4ЪзЯШдкашвЊЛЙдМСКЭO2ЕФЗДгІжагЩpMMOМЄЛюЁЃШЛКѓЃЌбмЩњЕФМзДМж№НЅбѕЛЏГЩМзШЉЁЂМзЫсбЮКЭЖўбѕЛЏЬМЃЌВњЩњСљИіПЩгУгкгабѕКєЮќЁЂЙЬЬМЛђ pMMO ЗДгІЕФЕчзгЁЃДѓЦјCH4дЄМЦбѕЛЏОњЛЙЛсЭЈЙ§ІСБфаЮИЫОњЫПАБЫсбЛЗЛђІУТэЯИОњКЫЭЊЬЧЕЅСзЫсЭООЖДгМзШЉжаЮќЪеЩњЮяжЪЁЃЛьКЯгЊбјЕФЩњЛюЗНЪНПЩФмЖдДѓЦјCH4гавцбѕЛЏОњвдЦНКтCH4ЕФЙ§ГЬЛљжЪКЭбѕЛЏОњЙЉгІБфЛЏжаЕФЛюЛЏЁЂНкФмКЭЬМЭЌЛЏЁЃ
Ш§ЁЂЩњЛЏЛљДЁ
3.1ДѓЦјH2бѕЛЏОњ
бѕЛЏH2ЕФ[NiFe]ЧтЛЏУИгаЫФжжИпЧзКЭСІЧтЛЏУИЃЈHhyЃЌHucЃЌHylЃЌHyoЃЉЕФДпЛЏФЃПщАќРЈСНИіЕААзжЪбЧЛљЃЈЭМ 3a-iЃЉЁЃДѓбЧЛљАќКЌ[NiFeЃЈCNЃЉ2CO]ИКд№H2ЕФДпЛЏДибѕЛЏКЭаЁбЧЛљАќКЌШ§ИіЬњСђЃЈFeSЃЉДиЃЌЫќУЧвРДЮзЊвЦH2-ЕчзгЪмЬхЕФбмЩњЕчзгЁЃЮЈвЛОпгаНсЙЙЬиеїДњБэЦзЯЕЕФHhyжаЃЌЦфжаСНИідаЭаЮГЩЖўОлЬхЃЌВЂЧвУПИіаЁбЧЛљЕФФЉЖЫFeSДиНєУмЯрЙиЃЌЪЙЕУУПИідаЭжЎМфЕФЕчзгзЊвЦдкРэТлЩЯЪЧПЩФмЕФЃЈЭМ3a-iiЃЌa-iiiЃЉЁЃДпЛЏДиТёдкДѓаЭбЧЕЅдЊЕФФкВПЃЌгЩH2ЭЈЙ§СЌНгЕНЕААзжЪБэУцЕФЪшЫЎЦјЬхЭЈЕРЁЃ H2НсКЯдкЬњКЭФјдзгжЎМфЃЌНјаадгНтДпЛЏбЛЗЃЌЦфжажЪзгПЩФмзЊвЦЕНФјХфЮЛАыызАБЫсжЎвЛЕФСђжаЃЌЪЃгрЕФЧтЛЏЮядкН№ЪєдзгжЎМфаЮГЩЧХСКЃЈЭМ 3a-IV ЃЉЁЃетжжвьНтбЛЗПЩвдгЩОљНтбЛЗВЙГфЁЃ
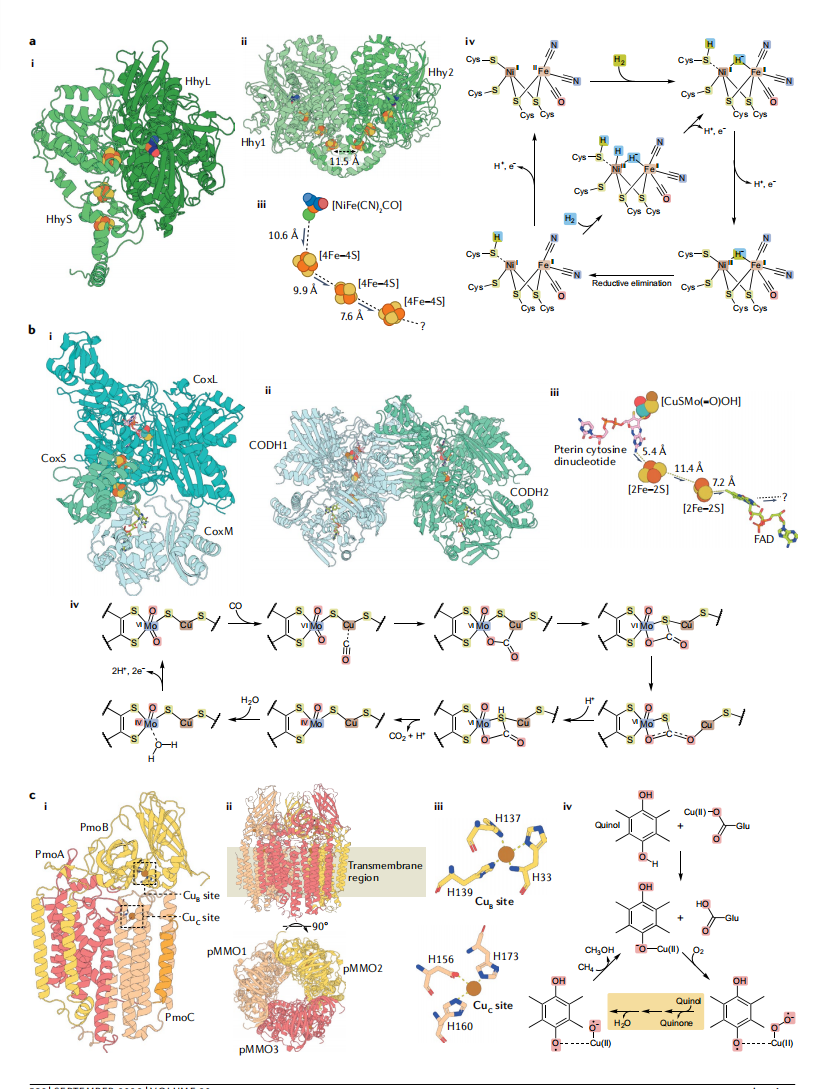
ЭМ3ЃКДѓЦјЧтЦјЁЂвЛбѕЛЏЬМКЭМзЭщбѕЛЏЕФHЩњЛЏЛљДЁЁЃ
A-I |РДздCupriavidus necatorЕФбѕВЛУєИаЛљЭХ1h [NiFe]-ЧтЛЏУИHhyЕФДпЛЏдаЭНсЙЙЃЌЯдЪОИЈвђзгКЭДѓЃЈHhyLЃЉКЭаЁЃЈHhySЃЉбЧЛљ[PDBЃК5AA5].A-II |гЩРДздC. necatorЕФСНИіHhyдаЭЬхаЮГЩЕФЩњРэЖўОлЬхЁЃжИЪОдЖЖЫЬњСђ ЃЈFeSЃЉ дЙЙЬхДижЎМфЕФОрРыЁЃA-III |[NiFe]ДпЛЏДиКЭHhyЕФFeSДигыC. necatorЕФЯрЖдЮЛжУКЭОрРыЁЃЧтЦкМфЕФЕчзгСїЃЈH2ЃЉбѕЛЏгУМ§ЭЗБэЪОЃЌЮДжЊЕчзгЪмЬхЕзЮяБэЪОЃЈЃПA-IV |HЕФДпЛЏЛњРэ2SiegbahnКЭLiaoЬсГіЕФ[NiFe]-ЧтЛЏУИЕФбѕЛЏЁЃH2ЭЈЙ§вьжЪбЛЗКЭОљНтбЛЗНјШыЃЌЗжБ№вдТЬЩЋКЭРЖЩЋЭЛГіЯдЪОЁЃB-I |РДздAfipia carboxidovoransЕФютвРРЕадвЛбѕЛЏЬМЭбЧтУИЃЈMo-CODHЃЉЕФДпЛЏдаЭНсЙЙЃЌЯдЪОИЈвђзгКЭCoxLЃЌCoxMКЭCoxLбЧЛљ[PDBЃК1N63]ЁЃB-II |гЩРДздA. carboxidovoransЕФСНжжMo-CODHдаЭЬхаЮГЩЕФЩњРэЖўОлЬхЁЃB-III |Э-ютЕћпЪДпЛЏИЈвђзгЁЂСНИі[2Fe-2S]ДиКЭРДздЬМмйоЃЕФMo-CODHЗжзгЕФЯрЖдЮЛжУКЭОрРыЁЃвЛбѕЛЏЬМЃЈCOЃЉбѕЛЏЙ§ГЬжаЕФЕчзгСїгУМ§ЭЗБэЪОЃЌЮДжЊЕчзгЪмЬхЕзЮягУМ§ЭЗБэЪОЃЈЃПB-IV |аьКЭЦНалЬсГіЕФMo-CODHДпЛЏЛњРэ.C-I |РДздMethylocystis sp. ATCC 49242ЕФПХСЃМзЭщЕЅМгбѕУИЃЈpMMOЃЉЕФНсЙЙЃЌЯдЪОPmoAЃЌPmoBКЭPmoCбЧЛљвдМАЭНсКЯCuЕФЮЛжУBКЭЭCЭјТч [PDBЃК4PI0]ЁЃC-II |ЩњРэШ§ОлЬхгЩРДздМзЛљцпзгГцЪєATCC 49242ЕФШ§ИіpMMOдаЭЬхаЮГЩЁЃC-III |ЭДІЕЅИіЭРызгЕФХфЮЛBКЭЭCРДздМзЛљцпзгГцЪєATCC 49242ЕФЭјеОЁЃC-IV |ХэЕШШЫЬсГіЕФpMMOДпЛЏЛњРэзлЪіЁЃЛЦЩЋПђМ§ЭЗЫѕаДСЫМИИіДпЛЏВНжшЃЈгаЙиЫљЬсГіЕФЛњжЦЕФЭъећУшЪіЃЌЧыВЮдФPengЕШШЫЃЉЁЃ
дкДпЛЏЖОЮяO2ЕФЛЗОГЫЎЦНЯТЃЌетаЉУИЕФЫФИіИпЧзКЭСІЦзЯЕШчКЮЗЂЛгзїгУЃЌФПЧАвЛЮоЫљжЊЁЃДѓЖрЪ§[NiFe]-ЧтЛЏУИЖдбѕИпЖШУєИаЃЌвђЮЊH2гыДпЛЏжааФНсКЯВЂЛЙдЮЊНєУмНсКЯЕФЛЏбЇЮяжЪЃЈOH−ЛђOOHЈC)ЁЃФЭбѕЧтЛЏУИЕФвЛИібЧзщЃЌМД1dзщ[NiFe]-ЧтЛЏУИЃЈHyaЃЉЃЌПЩвддкЮЂбѕЬѕМўЯТЗЂЛгзїгУЃЌДгЖјжЇГжЕЭЧзКЭСІЧтгЊбјОњЕФгабѕЩњГЄЁЃЫфШЛБЛO2вжжЦЃЌетаЉУИвбОНјЛЏГіЖРЬиЕФНќЖЫ6Cys[4Fe-3S]ДиЃЌЭЈЙ§ЫФИіЕчзгЕФЗДЯђзЊвЦНЋжївЊгыЫЎНсКЯЕФЮяжЪМѕЩйЃЌДгЖјНЋЧтЛЏУИЛжИДЕНДпЛЏОЭаїзДЬЌЁЃЯрБШжЎЯТЃЌHhyЃЌHucЃЌHylКЭHyoЫЦКѕЖдбѕЦјВЛУєИаЃЌвђЮЊЫќУЧдквЛЯЕСаЮяжжЕФЛЗОГЬѕМўЯТЗЂЛгзюМбЙІФмЁЃКЭ O2зюаЁвжжЦДПЛЏЕФHhyгыHyaЕШаЇЕФдйМЄЛюЛњжЦВЛЬЋПЩФмЃЌвђЮЊЫќЬЋТ§ЧвФмСПАКЙѓЃЌЮоЗЈдкЛЗОГЬѕМўЯТгааЇЃЌВЂЧвЫљгаЫФжжУИЦзЯЕЖМШБЗІНсКЯ6Cys[4Fe-3S]ДиЫљашЕФАыызАБЫсВаЛљЁЃвЛжжПЩФмадЪЧетаЉУИКЌгаНЯеЕФЦјЬхЭЈЕРЃЌзшжЙO2НјШыЃЌОЁЙмвВЛсНЕЕЭ H 2ЕФРЉЩЂЫйТЪЃЌНјЖјМѕЩйДпЛЏЛюадЁЃгыДЫвЛжТЃЌбѕУєИаЧтЛЏУИЕФЦјЬхЭЈЕРЫѕаЁвВЛсНЕЕЭ O2СщУєЖШКЭ H2ЕФбѕЛЏЫйТЪЁЃ
ЧјЗжИпЧзКЭСІКЭЕЭЧзКЭСІ[NiFe]-ЧтЛЏУИЕФНсЙЙЬиеївВШдЮДНтОіЁЃетЗДгГСЫЩаЮДЖдИпЧзКЭСІЧтЛЏУИНјааНсЙЙЛђЩњЛЏБэеїЁЃОЁЙмРДздC. necatorЕФHhyвбдкНсЙЙЩЯБэеїЃЌЕЋИУУИЫЦКѕЖдH2ЮоТлЪЧдкећИіЯИАћжаЛЙЪЧДПЛЏЪБЃЌгыЫљгаЦфЫћЬиеїЮяжжжаЕФHhyЯрЗДЃЈБэ1ЃЉЁЃРДздИЛТэРяРћЫсМзЫсОњЕФHhyЪЧЮЈвЛФмЙЛбѕЛЏДѓЦјH2ЕФУИвбЗжРыЕНЁЃетжжУИдкШЋЯИАћКЭФЄЬсШЁЮяжаФмбѕЛЏбЧДѓЦјбЙХЈЖШЕФH2ЃЌЕЋбаОПБэУїЫќашвЊёюКЯЕНЕчзгДЋЕнСДВХФмвдИпЧзКЭСІдЫааЁЃЦфЫћПЩвддіМгЧзКЭСІЕФаоЪЮАќРЈжиЫмДпЛЏжааФЃЌFeSДиЛђЦфХфЮЛЛЗОГвддіЧПH2РІАѓЁЃ
3.2ДѓЦјвЛбѕЛЏЬМбѕЛЏЃЈЪЁТдЃЉ
3.3ДѓЦјCH4бѕЛЏЃЈЪЁТдЃЉ
ЫФЁЂКлСПДѓЦјбѕЛЏЮЂЩњЮяЕФЩњЬЌживЊад
4.1ЭСШРЩњЬЌЯЕЭГ
ОЁЙмЮЂЩњЮяЮЂСПЦјЬхбѕЛЏЕФЩњЮяЕиЧђЛЏбЇзїгУвбЕУЕНГфЗжШЯПЩЃЌЕЋетвЛЙ§ГЬЕФЩњЬЌживЊадВХИеИеПЊЪМЕУЕННтОіЁЃетЗДгГСЫКлСПЦјЬхбѕЛЏГЄЦквдРДвЛжББЛШЯЮЊЪЧЗЧЩњВњадЙ§ГЬЃЌЛђОжЯогкЯЁгаЩњЮяШІЁЃШЛЖјЃЌзюНќЖдВЛЭЌЭСШРЕФКъЛљвђзщЕїВщБэУїЃЌДѓЖрЪ§ЯИОњПЩвдЧхГ§КлСПЦјЬхЃКЩуШЁЧтЦјЕФЧтЛЏУИжївЊЪЧHhyЁЂMo-CODHsКЭpMMOЕФЛљвђБрТыЁЃетаЉЛљвђДцдкгкУПИіжївЊЭСШРУХЕФКъЛљвђзщзщзАЛљвђзщжаЃЌЕЋвВДцдкгкЮДХрбјЕФКђбЁУХжаЃЌР§ШчDormibacterotaЃЌEremiobacterotaКЭMethylomirabilotaЃЈЭМ4ЃЉ. ЭЌбљЃЌЫќУЧКИЧСЫЗсИЛЕФЭЈВХЃЌШчЛКИљСіОњЪєКЭЗжжІИЫОњЪєЃЌвдМАЯЁгаЩњЮяШІЕФжкЖрГЩдБЁЃетБэУїДѓЦјH2КЭCOбѕЛЏЪЧвЛжжЙуЗКЕФЬиеїЃЌЪЙЯИОњФмЙЛеМОнЙуЗКЕФЩњЬЌЮЛЃЌЖјДѓЦјCH4бѕЛЏЪЧвЛжжЯСеЕФбѕЛЏЃЌЪЙЯИОњФмЙЛдкИќЗсИЛЕЋЯрЖдЖшадЕФЕзЮяЩЯзТзГГЩГЄЁЃетжжЭЦТлЕУЕНСЫЦфЫћзюНќЕФКъЛљвђзщбаОПКЭХрбјЯИОњЕФЛљвђзщЕїВщЕФжЇГжЃЈЭМ1ЃЉЁЃ
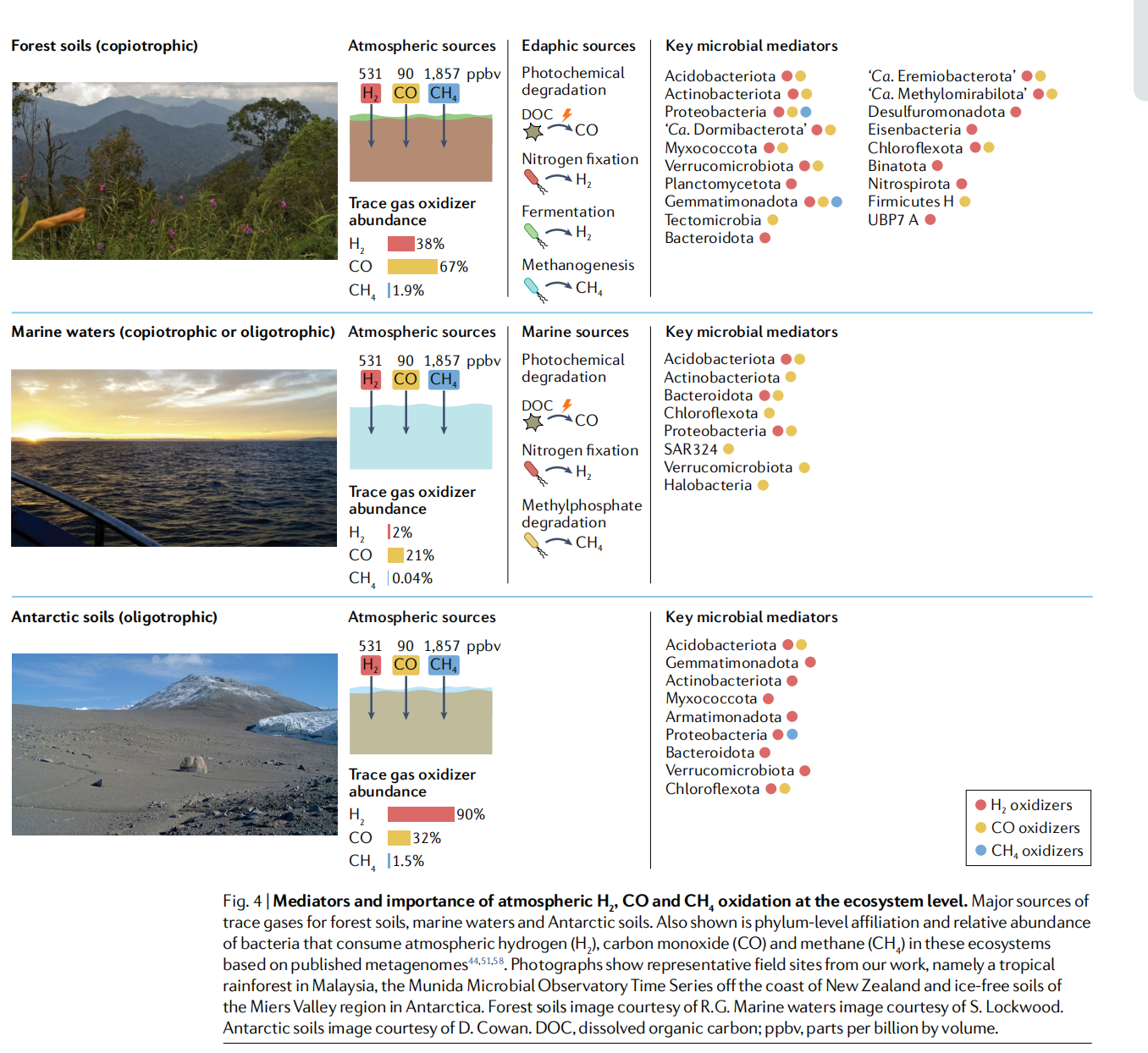
ЭМ4ЃКДѓЦјH2CO КЭ CH4бѕЛЏЕФжаНщвђЫиКЭЩњЬЌЯЕЭГживЊадЁЃ
ЮЂСПЦјЬхЕФжївЊРДдДгкЩСжЭСШРЁЂКЃЫЎКЭФЯМЋЭСШРЁЃЛЙЯдЪОСЫУХЫЎЦНЕФСЅЪєЙиЯЕКЭЯћКФДѓЦјЧтЕФЯИОњЕФЯрЖдЗсЖШЃЈH2ЃЉЁЂвЛбѕЛЏЬМ ЃЈCOЃЉ КЭМзЭщ ЃЈCH4ЃЉдкетаЉЛљгквбЗЂБэЕФКъЛљвђзщЕФЩњЬЌЯЕЭГжаЁЃееЦЌЯдЪОСЫЮвУЧЙЄзїжаОпгаДњБэадЕФвАЭтЕиЕуЃЌМДТэРДЮїбЧЕФШШДјгъСжЃЌаТЮїРМКЃАЖЕФMunidaЮЂЩњЮяЙлВтеОЪБМфађСаКЭФЯМЋжоУзЖћЫЙКгЙШЕиЧјЕФЮоБљЭСШРЁЃЩСжЭСШРЭМЦЌгЩR.G.КЃбѓЫЎгђЭМЦЌЬсЙЉЃЌгЩS. LockwoodЬсЙЉЁЃФЯМЋЭСШРЭМЦЌгЩD. CowanЬсЙЉЁЃDOCЃЌШмНтгаЛњЬМ;ppbvЃЌЪЎвкЗжжЎвЛЃЈАДЬхЛ§МЦЫуЃЉЁЃ
ФЧУДЃЌКлСПЦјЬхбѕЛЏЬсЙЉСЫФФаЉбЁдёадгХЪЦФиЃПЭСШРжаЕФДѓЖрЪ§ЯИОњЩњЛюдквЛЯЕСаанУпзДЬЌжаЃЌгаЛњЬМЯожЦБЛШЯЮЊЪЧЯожЦЫќУЧЕФжївЊвђЫиЁЃДгРэТлЩЯНВЃЌДѓЖрЪ§ПЩвдЭЈЙ§ГжајбѕЛЏДѓЦјжаЕФКлСПЦјЬхРДЮоЯоЦкЕиЩњДцЁЃШШСІбЇФЃаЭБэУїЃЌДѓЦјH2ЭСШРжаЗЂЩњЕФвЛбѕЛЏЬМбѕЛЏЃЌДг0ЕН30РхУзЕФЩюЖШЃЌзувдЮЌГжЫљгааЏДјЛљвђЕФЯИОњРДбѕЛЏетаЉЦјЬхЁЃвЛжТЕиЃЌДѓЦјH2ЕФЯрЖдЗсЖШКЭЯИАћЬивьадЫйТЪбѕЛЏОњЫцзХбјЗжПЩгУадЕФНЕЕЭЖјдіМгЃЌР§ШчдкИЩКЕЬнЖШЩЯЁЃгІИУзЂвтЕФЪЧЃЌКлСПЦјЬхбѕЛЏЫйТЪПЩФмвђЯИАћЖјвьЃЌОЁЙмЕЅЯИАћЗНЗЈВЛФмЧсвзЧјЗжЛюадЃЌвђЮЊH2ВЂЧвЃЌЭЈГЃЃЌCOВЛжБНгБЛЭЌЛЏЕНЩњЮяжЪжаЃЈгыCH4).ДѓЖрЪ§анУпЯИОњПЩФмЭЈЙ§ЯћКФКлСПЦјЬхвдМАФмСПДЂБИКЭЦфЫћЭтдДадЛЏКЯЮяРДЛьКЯЩњЛюЃЌетвВЕУЕНСЫЛљгкХрбјЕФбаОПЕФжЇГжЁЃШЛЖјЃЌКлСПЦјЬхбѕЛЏЪЧГЄЦкЮЌГжЯИОњЕФзюПЩППЛњжЦЁЃЯИОњдканУпзДЬЌЯТЩњДцЕФФмСІБЛШЯЮЊЭЈЙ§діМгЭСШРШКТфЕФЗсИЛЖШЕЋМѕЩйИќЬцРДЫмдьЭСШРШКТфЕФЩњЮяЖрбљадЃЌДгЖјдіЧПЙиМќЩњЬЌЯЕЭГЙІФмКЭЗўЮёЕФИДдСІЁЃР§ШчЃЌбЧЯѕЫсбЮбѕЛЏЯИОњКЭЬМЁЂЕЊКЭСђбЛЗжаЕФаэЖрЦфЫћЙиМќНщжЪдкКлСПЦјЬхжаДцЛюЕФФмСІПЩФмЪЙЫќУЧФмЙЛдкЛЗОГБфЛЏЕФЧщПіЯТжДааЙиМќЕФЕїНкЙІФмЁЃвђДЫЃЌКлСПЦјЬхбѕЛЏОњЕФживЊаддЖдЖГЌГіСЫЫќУЧдкЕїНтЭСШРЛужаЕФзїгУЁЃ
ЮЂСПЦјЬхбѕЛЏвВгажњгкЭСШРЩњЬЌЯЕЭГжаЕФЯИОњЩњГЄКЭГѕМЖЩњВњЁЃетдкДѓЦјCH4жаЕУЕНСЫзюКУЕФЗДгГбѕЛЏОњЁЃЭЈЙ§ЪЙгУЖРЬиЕФДѓЦјЛљжЪВЂВЩгУЛьКЯгЊбјЩњГЄВпТдЃЌАќКЌM. gorgonaЕФUSCІСЗжжЇвбГЩЮЊЩњОГЖрУцЪж;ЫќвбОжГУёСЫЪРНчИїЕиЖрбљЛЏЕФТНЕиЩњЬЌЯЕЭГЃЌДгЮТДјВнЕщЕНШШДјАзвЯЧ№дйЕНгРОУЖГЭСЁЃЦфЫћЭСШРЯИОњПЩФмЭЈЙ§ЯћКФДѓЦјжаЕФH2РДНщЕМЛьКЯгЊбјЩњГЄКЭгаЛњЬМЕФвЛбѕЛЏЬМЁЃИљОнКъЛљвђзщбЇЕїВщЃЌДѓдМ12%ЕФЭСШРЯИОњПЩвдЪЙгУH2-бмЩњЛђCOбмЩњЕФЕчзгвдЭЌЛЏCO2ЪЙгУRuBisCOЕФЛЏбЇКЯГЩЗжжЇЁЃ.етаЉЯИОњЕФвЛИізгМЏПЩФмЪЧбвЪЏздбјЩњЮяЃЌЫќУЧЪЙгУЕЭЧзКЭСІЧтЛЏУИЛђMo-CODHsЃЌгЩгкЕЊЙЬЖЈКЭЙтЛЏбЇЗДгІЬсЙЉH2ЃЌЪЙгУФГаЉЛЗОГЃЈР§ШчИљМЪЃЉжаПЩгУЕФИпЦјЬхЙЉгІРДЩњГЄКЭвЛбѕЛЏЬМЃЈЭМ4;Бэ 1ЃЉЁЃЕЭЧзКЭСІКЭИпЧзКЭСІH2ЕФЙВДцЭСШРжаЕФбѕЛЏОњДгКъЛљвђзщЭМЦзжаКмУїЯдЃЌH2ЕФЫЋЯрЖЏСІбЇдкгаЛњЭСШРжаЙлВьЕНбѕЛЏвдМАБШНЯЩ§ИпКЭЛЗОГ H2гАЯьЕФЮЂЙлбаОПЕФЙлВьНсЙћЁЃашвЊНјвЛВНЕФбаОПРДНтОіH2ЕФФкдДадВњЩњКЭФкВПбЛЗСПвЛбѕЛЏЬМЗЂЩњдкЭСШРЩњЬЌЯЕЭГжаЁЃ
4.2ЫЎЩњЩњЬЌЯЕЭГ
ЮЂСПЦјЬхбѕЛЏОњвВДцдкгкКЃбѓЫЎгђЁЂКЃбѓГСЛ§ЮяКЭЕЫЎЩњЬЌЯЕЭГжаВЂЛюдОЁЃдкКЃбѓБэВуЫЎжаЃЌДѓдМЫФЗжжЎвЛЕФЯИОњЯИАћКЌгабѕЛЏДѓЦјвЛбѕЛЏЬМЕФЛљвђЃЌАќРЈКьИЫОњПЦЃЈКЃбѓУЕЙхИЫОњНјЛЏжІЃЉжаЕФЗсИЛЖрУцЪжЃЈЭМ4ЃЉ. етаЉЯИОњЭЈГЃБЉТЖгкИпХЈЖШЕФ COЃЌвђЮЊгаЛњЮяЕФЙтЛЏбЇНЕНтЛсдк DIEL бЛЗжаВњЩњ COЃЌОЁЙмЫќУЧвВПЩвдЯћКФЕЭгкДѓЦјЫЎЦНЕФвЛбѕЛЏЬМЁЃгыЭСШРЩњЬЌЯЕЭГвЛбљЃЌвЛбѕЛЏЬМбѕЛЏПЩФмжївЊжЇГжЯИОњдкЕЭгЊбјПЊРЋКЃбѓжаЕФГЄЦкЩњДцЁЃЛљгкХрбјЕФбаОПБэУїЃЌCOВЛгАЯьR. pomeroyiЕФЩњГЄЃЌОЁЙметжжЯИОњКЭНќЧздкМЂЖіЦкМфЖМЧПСвЩЯЕїMo-CODHЁЃКЃбѓвЛбѕЛЏЬМЫЎЦНЫцЩюЖШбИЫйЯТНЕЃЌДгФЩФІЖћХЈЖШЕНЦЄФІЖћХЈЖШЃЌетЪЧгЩгкЙтВњСПМѕЩйКЭЧБдкЕФЯћКФдіМгЁЃШЛЖјЃЌMo-CODHЛљвђдкЩюКЃКЭБљМмвдЯТЫЎгђЕШЙтадЫЎгђжаКЌСПЗсИЛВЂзЊТМЃЌетБэУїCOШдШЛЪЧвЛжжживЊЕФФмдДРДдДЁЃ
ЯрБШжЎЯТЃЌИпЧзКЭСІ H2бѕЛЏЪЧКЃбѓЯИОњЕФЯожЦадзДЁЃвбОБЈЕРСЫБрТыгыЛьКЯгЊбјЩњГЄЃЈжївЊЪЧHucЃЉКЭГЄЦкГжајадЃЈжївЊЪЧHylЃЉЯрЙиЕФЧтЛЏУИЕФЯИОњЃЌОЁЙмгЩгкЮДжЊдвђЃЌЫќУЧНіеМЕиБэЫЎШКТфЕФ1%ЁЃШЛЖјЃЌетаЉЯИОњНщЕМИпЯИАћЬивьадЕФH2бѕЛЏЃЌБэУїЫќУЧЪмвцгкЯрЖдХХЫћадЕФЕзЮяЁЃРрЫЦгкЭСШРЩњЬЌЯЕЭГЕФЗЂЯжЃЌH2дкИќЙбгЊбјЕФЕиБэЫЎжаЃЌбѕЛЏТЪИќИпЃЌР§ШчдкбиКЃЕНбЧФЯМЋбљДјЩЯдіМгЁЃЗДЙ§РДЃЌетаЉЯИОњПЩФмЪЧH2дкМЋЕиЫЎгђгыДѓЦјЕФБШНЯЁЃЙбгЊбјМЋадЯИОњАЂРЫЙМгСДЧђОњЕФДПХрбјЮяЙВбѕЛЏДѓЦјH2ЩњГЄЙ§ГЬжаЕФгаЛњЛЏКЯЮяЁЃЭЌбљЃЌвЛЯюКъЛљвђзщбаОПЗЂЯжЃЌдкЩюКЃH2жаЃЌЯИОњаЏДјЕЭЧзКЭСІЩњГЄжЇГжЧтЛЏУИЕФзЊБфЁЃИЛКЌШШвКг№СїЃЌдкЯрЖдЕЭгЊбјЕФжмЮЇЫЎгђжаОпгаИпЧзКЭСІЕФГжОУаджЇГжУИЁЃ
4.3ДѓЦјЧ§ЖЏЕФЩњЬЌЯЕЭГ
дкДѓЖрЪ§ЩњЬЌЯЕЭГжаЃЌЭЈЙ§КлСПЦјЬхбѕЛЏЙЬЖЈЕФЬМПЩФмжЛеМжВЮяКЭЦфЫћЙтздбјЩњЮяВњЩњЕФЬМЕФвЛаЁВПЗжЁЃШЛЖјЃЌФГаЉМЋЖЫЛЗОГПЩФмжївЊгЩДѓЦјФмСПЧ§ЖЏЃЌЬиБ№ЪЧФЧаЉЮяРэЛЏбЇЬѕМўХХГ§ЙтздбјЩњЮяКЭЕижЪФмдДЯЁШБЕФЛЗОГЁЃдЄМЦФЯМЋжоРфЖГЃЌИЩдяЃЌМОНкадКкАЕЭСШРжаЕФжївЊГѕМЖЩњВњепЪЧЮЂСПЦјЬхбѕЛЏОњЁЃ.дкТѓЛљБљДЈЕиЧјЃЌ90%ЁЂ32%КЭ27%ЕФЯИОњЗжБ№БрТыЧтЛЏУИЃЈжївЊЪЧHylЃЉЃЌMo-CODHКЭRuBisCOЁЃЃЈЭМ4ЃЉ. зюЗсИЛЕФЕЅвЛЯИОњЪЧРДздUBA7966ЗжжЇЃЈUSCІУЃЉЕФдЄВтЪШМзЭщОњЃЌНјвЛВНЧПЕїСЫCHЕФвцДІ4зїЮЊзЈгУЛљВФ51.ЪМжеШчвЛЕиЃЌетаЉЭСШРжаЕФЯИОњШКТфЛсбИЫйЯћКФДѓЦјжаЕФH2ЁЂCO КЭ CH4КЭЛЏбЇКЯГЩЙЬЖЈЖўбѕЛЏЬМдкгыЛЗОГЯрЙиЕФЮТЖШЯТЃЌвдзувдЮЌГжЕФЫйЖШЃЌвдМАвЛаЉЩчЧјГЩдБЕФЧБдкЛКТ§діГЄЁЃШЛЖјЃЌашвЊИќСщУєЕФЗНЗЈРДШЗЖЈЯИОњЪЧЗёЙЬЖЈЖўбѕЛЏЬМдкЛЗОГЦјЬхХЈЖШЯТЃЌНјЖјНщЕМЯИАћЗжСбЁЃжЕЕУзЂвтЕФЪЧЃЌДѓЦјжаВњЩњЕФДњаЛЫЎH2бѕЛЏзувддк 15 ЬьФкЬцЛЛЫљгаЯИАћЫЎ.етаЉМйЖЈЕФГѕМЖЩњВњепПЩФмгыДцдкЕФЦфЫћЩњЮяЬхНЈСЂЙВЩњКЭгЊбјЙиЯЕЁЃ
ЛљгкетвЛИХФюЃЌПЩФмДцдкИїжжЦфЫћДѓЦјЧ§ЖЏЕФЩњЬЌЯЕЭГЁЃгЩгкЮяРэЛЏбЇЬѕМўЯожЦЃЌНЋДѓЖрЪ§ЙтКЯГѕМЖЩњВњепХХГ§дкГЌИЩКЕЩГФЎЁЂСзШмЖДбЈЁЂИпбЮКўВДКЭИпКЃАЮЩНЧјжЎЭтЃЌЕЋПЩФмЮЌГжЪЙгУДѓЦјКлСПЦјЬхЕФГѕМЖЩњВњепЁЃгыетвЛЙлЕуЯрвЛжТЃЌАЂЫўПЈТэЁЂИъБкЁЂФкИЧЗђКЭФЩУзВМЩГФЎвдМАЧрВиИпдЕШГЌИЩКЕЕиЧјЕФДѓЖрЪ§ЯИОњвВФмЙЛЧхГ§ДѓЦјжаЕФКлСПЦјЬхЁЃЭЌбљЃЌбЮЬяжагааэЖрДѓЦјжаЕФвЛбѕЛЏЬМбѕЛЏЯИОњКЭЙХЯИОњЃЌЖјUSCІУМзЭщгЊбјОњдкПІЫЙЬиЖДбЈЯЕЭГжазТзГГЩГЄ.ШЛЖјЃЌашвЊНсКЯКъЛљвђзщбЇЁЂЩњЮяЕиЧђЛЏбЇКЭНЈФЃЗНЗЈЕФЙуЗКВЩбљЛюЖЏЃЌвдЯЕЭГЕиСЫНтКлСПЦјЬхбѕЛЏгыетаЉЩњЬЌЯЕЭГжаЦфЫћНкФмКЭГѕМЖЩњВњЙ§ГЬЯрБШЕФЯрЖдЙБЯзЁЃ
зюКѓЃЌКлСПЦјЬхбѕЛЏОњдкЩњЬЌЯЕЭГРЉЩЂКЭЖЈжВжаЕФзїгУжЕЕУЬНЬжЁЃМјгкЫќУЧЖдЮоДІВЛдкЕФЛљжЪЕФвРРЕЃЌетаЉЯИОњПЩФмЛсдкаТЛЗОГжабИЫйЖЈжВВЂДйНјГѕМЖбнЬцЃЌР§ШчЛ№ЩНГСЛ§ЮяЁЂБљДЈЧАГЁЁЂЮВПѓЩѕжСдЩЪЏЁЃбаОПБэУїЃЌЯФЭўвФЕФЛ№ЩНГСЛ§ЮяЯрЖдНЯПьЕиБЛCOКЭH2бѕЛЏЕФИїжжЮЂЩњЮяШКТфЖЈжГЁЃЪШМзЭщОњКЭвЛбѕЛЏЬМбѕЛЏОњЖМПЩвджЇГжжВЮяШКТфЕФЗЂеЙЁЃДЫЭтЃЌЯИОњдкПеЦјжаЩњДцЕФФмСІПЩФмдкДѓЦјжаДЋВЅЙ§ГЬжаЗЂЛгвдЧАЮДБЛШЯЪЖЕНЕФзїгУЁЃ
4.4ЬьЬхЩњЮябЇвтвх
ЮЂЩњЮявВПЩвдбѕЛЏЯЕЭтаааЧЩЯЕФДѓЦјКлСПЦјЬхЃЌетЪЧКЯРэЕФЁЃЙигкаааЧБЃЛЄЃЌПЩвдЯыЯѓЃЌШчЙћНЋФГаЉКлСПЦјЬхбѕЛЏОњв§ШыЯЕЭтаааЧЃЌР§ШчЛ№аЧЃЌетРягаЯЁБЁЕФИЛКЌвЛбѕЛЏЬМЕФДѓЦјВуКЭИЛКЌИпТШЫсбЮЕФЭСШРЃЌЮЂЩњЮявВПЩФмавДцЯТРДЁЃДЫЭтЃЌаэЖрЯЕЭтаааЧЩЯвбМьВтЕНгаВЛЭЌХЈЖШЕФH2ЃЌЩњУќОЭгаПЩФмДцдкетаЉаЧЧђЁЃH2ПЩФмЭЦЖЏСЫЕиЧђЩЯЕФЩњЮяЗЂЩњВЂЧвЪЧвЛжжРэЯыЕФФмдДЃЌдкЦфЫћаЧЧђЩЯЧтЦјвВПЩФмКмЗсИЛЁЃетаЉИХФюдкзюНќвЛаЉЮФЯззлЪіжаЕУЕНСЫОпЬхЬНЬжЁЃ
ЮхЁЂНсТлКЭеЙЭћ
ДѓЦјКлСПЦјЬхЕФЮЂЩњЮябѕЛЏЪЧЮЂЩњЮябЇКЭЩњЮяЕиЧђЛЏбЇСьгђЕФживЊаТЧАбиЁЃОЁЙмзЪдДЯЁШБЃЌаэЖрЯИОњКЭвЛаЉЙХОњвбОНјЛЏГівЛжжМђЕЅЖјгХбХЕФЩњЛюЗНЪНЃКРћгУПеЦјжаЮЂСПЦјЬхЮЊФмСПЮЌГжЩњДцЁЃбаОПШЫдБЯждкЖдетвЛЙ§ГЬЕФЮЂЩњЮяНщжЪЁЂУИОіЖЈвђЫиКЭЩњРэзїгУОљгаСЫЯрЖдЩюПЬЕФРэНтЁЃ
ДѓЦјКлСПЦјЬхЕФЮЂЩњЮябѕЛЏЕФЩњЬЌживЊадвВдНРДдНБЛРэНтЃККлСПЦјЬхбѕЛЏЮЌГжСЫЖрЪ§анУпЕФЮЂЩњЮяЃЌЪЙМЋЖЫЩњЬЌЯЕЭГЕФЩњЮяЩњВњСІГЩЮЊПЩФмЃЌетаЉЛЗОГЯТЙтздбјЩњЮяФбвдВЮгыЃЌВЂМѕЧсСЫДѓЦјЮлШОЮяЕФШЫЮЊХХЗХЁЃ
етвЛСьгђШдШЛДцдкаэЖржЊЪЖашвЊНјвЛВНЬНЫїЭъЩЦЁЃдкЛњРэВуУцЃЌгаБивЊШЗЖЈЧјЗжЕЭЧзКЭСІКЭИпЧзКЭСІЦјЬхбѕЛЏУИЕФНсЙЙВювьЃЌВЂИќКУЕиНтОіетаЉУИЪЭЗХЕФЕчзгШчКЮгУгкгабѕКєЮќКЭЙЬЬМЁЃЭЌбљЃЌШчКЮДгЯИАћКЭЩњЬЌЯЕЭГЫЎЦНЩЯЕїНкКлСПЦјЬхбѕЛЏЃЌвдМАЛЗОГвђЫиШчКЮгАЯьетаЉЙ§ГЬЃЌЛЙгаД§ШЋУцСЫНтЁЃЛЙашвЊИДдгЕФЗНЗЈРДЛёЕУдЮЛКлСПЦјЬхбѕЛЏЕФЕЅЯИАћЪгНЧЃЌвдМАдкЭСШРКЭКЃбѓЩњЬЌЯЕЭГжаЧјЗжФкдДКЭЭтдДЕФКлСПЦјЬхЁЃЛЙашвЊНјвЛВНЕФбаОПРДИќКУЕиСЫНтДѓЦјКлСПЦјЬхШчКЮПижЦжВЮяЁЂЖЏЮяКЭЮЂЩњЮяжЎМфЕФНЛЛЅзїгУЃЌАќРЈЦфХЉвЕКЭвНбЇживЊадЁЃЮЂЩњЮяПЩвдЁАЩњЛюдкПеЦјжаЁБЕФживЊЗЂЯжЃЌгаПЩФмИФБфЮвУЧЖдЕиЧђЩѕжСгюжцЩњУќЕФРэНтЁЃ
https://blog.sciencenet.cn/blog-41174-1379996.html
ЩЯвЛЦЊЃКЕААзЫЎНтвЉЮяПЊЗЂаТШќЕР
ЯТвЛЦЊЃКЧтЦјвНбЇШ§ДѓжЇжљ
