博文
STING Pathway in Cancer Immunology
 精选
精选
|
STING Pathway in Cancer Immunology
之前简单回顾了“cGAS-cGAMP-STING”通路的发现过程,时间过的很快,2008年首次报道STING,2012年底揭示上游通路的关键分子cGAS和第二信使cGAMP。随即在2013年11月,Aduro BioTech(a clinical-stageimmunotherapy company located in Berkeley, California)就在 Societyfor Immunotherapy of Cancer (SITC)的年会上展示了他们筛选到的化合物在临床前研究的数据,突出显示了刺激STING通路后可明显增强机体抗肿瘤的免疫能力。他们基于刺激STING活性的天然小分子cGAMP的结构优化后得到MIW815(2016年2月进入临床Phase I,NCT02675439),in vitro的实验显示该化合物对磷酸二酯酶(phosphodiesterase)不敏感,可以高水平的活化STING、激活PBMC的活性;In vivo实验的结果也显示该分子可明显增强CD4 和CD8 T细胞抗原特异性的攻击效应,抑制肿瘤细胞的生长。2015年三月底,Novartis与Adruo达成7.5亿美元的交易(首付资金2亿美元),借此机会希望增强在免疫疗法市场上的优势。Novartis在免疫疗法中确实是大佬:CAR-T的CTL019已进入临床 phase 2,靶向免疫节点的PD1、LAG3和 TIM3也在同时进行,他们希望联合STING通路上调免疫的能力的获得更好、更广泛的抗癌疗效。
The STING signaling pathway
STING由379个氨基酸构成,有数个跨膜结构域,在多种内皮细胞、上皮细胞中以及造血肝细胞均有表达。这条信号通路简单、干净、有效:细胞质内的DNA与522个氨基酸组成的cGAS结合后被催化成的第二信使cGAMP(此过程需要ATP和GTP);接着二聚化的STING与cGAMP结合,发生构象变化,通过自噬小体(autophagsome)经ER、Golgi再转战到内体,期间被泛素化后募集TBK1蛋白,再被磷酸化,之后与IRF3结合;接着IRF3被磷酸化后入核,诱导表达IFNs,细胞因子、T细胞募集因子等进而增强机体的免疫能力,更大限度的杀伤肿瘤细胞。其实cGAMP还可以透过细胞间的空隙连接(gapjunctions)进入邻居免疫细胞中,活化更多细胞中的STING通路,这也是人们希望得到其类似物的原因之一吧,已拥有较多的Proof-of-concept。Aduro就致力于合成类似cGAMP的环二核苷酸(cyclicdinucleotide, CDN)STING的激动剂,持续活化该信号通路。其实这里面有很多故事,应朋友的建议,在此再稍稍介绍一下STING是如何一步步成为药物研发中热门的靶点,如何让专注免疫疗法的各位如此心动,还请各位不吝指正。
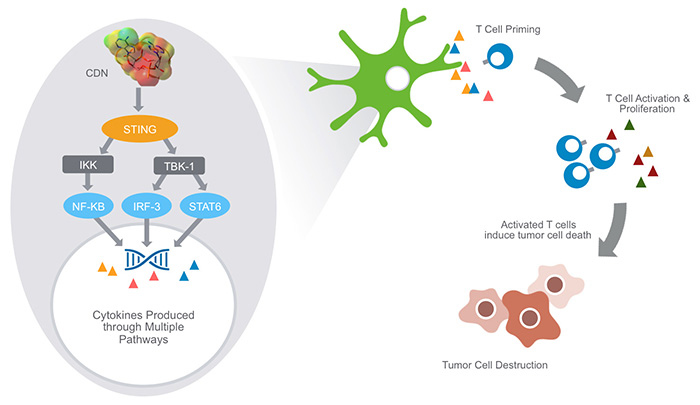
CDNs have shown promising therapeutic activity in cancer(From Aduro)
最近阅读STING的相关文章,有一件事让我觉得很有趣:重磅的发现总是在权威期刊连续刊出,比如这次看到两篇关于STING结构生物学的文章,一篇来自Texas A&M University的Dr.Pingwei Li的团队,解析出 STING C-末端的结构域(155-341aa)与c-di-GMP结合的结构(Page 722-4);[1]一篇来自山东大学的Dr. Lichuan Gu的团队,解出C-末端149-379 aa与c-di-GMP复合体的三维结构(Page 725-7)。[2]双方共同揭示STING可二聚化形成V型结构与c-di-GMP结合的分子机制。我很好奇的是两个独立的团队是如何在同一时间段、利用类似的方法、解析出同一个蛋白的C端结构域?虽说在氨基酸的位置上有1的差异,可两篇文章利用结构上的提示同时发现Y239S和E259A(或E260A)的突变让STING失去结合c-di-GMP的能力。(可能也提示了人们前期临床试验失败的原因,见下文)
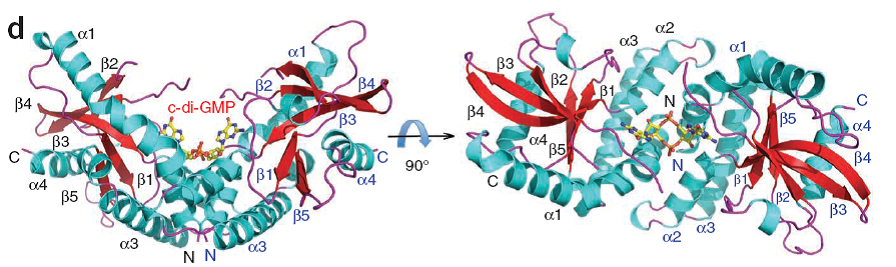
STING forms a dimer and binds c-di-GMP at the dimerinterface [1]
STING and cancer
其实肿瘤细胞存在的地方同时伴有很多的先天免疫相关的细胞,他们时而无视、时而也会互助肿瘤细胞的增殖。有一个研究方向就是如何能激活这些隐藏在肿瘤细胞中的免疫细胞,如何让他们从“良民”变成“军人”,如何在敌人的大本营攻击肿瘤细胞。1991年人们发现FAA(flavonoidsflavone acetic acid)可以调节肿瘤细胞中免疫活性;2005年发现DMXAA(5,6-dimethylxanthenone-4-acetic acid)可以激活NFκB信号通路,后续看到他可以破坏肿瘤血管的发生,2007年发现他还可以激活TBK1-IRF3信号通路。诸多让人激动的实验数据将他们推向临床试验,可惜未能如愿,最终以失败告终,其中DMXAA已走到临床试验Phase III。人源STING的晶体结构为此提供了理论解释,2014年发现DMXAA和FAA可以结合mSTING(mouseSTING),却无法结合hSTING(human STING)。[3]到此我倒是想问:为何人们在分析化合物对STING功能时未曾观察到对hSTING的作用?为何检测IFNβ释放时不选用THP1,而始终在鼠巨噬细胞系Raw264.7分析?为何在早期不观察化合物对hSTING的结合呢?可惜,最终只能说这两个化合物是mSTING特异的激动剂。接下来的任务就是重新来过,设计出特异于hSTING的激动剂,激活肿瘤旁边的机体免疫能力。
在人们都在关注CDNs时,有人留意到细胞质中的RNA和DNA的杂交分子也可以激活cGAS-STING通路。[4]虽说这类分子在病毒感染的情况下经常产生,停下来想一下:该如何设计这样的实验?如何保证RNA:DNA的分子在细胞中接触STING之前保持完整?或如何人工合成一种稳定的杂交分子?其实搜索前人的工作不难发现曾有人研究过杂交分子在细胞中的生物学作用,那么剩下的问题就相对简单些。文中发现该杂交分子可激活细胞表达抗病毒蛋白、并非参与与RIG-I和MDA5依赖的MAVS通路,而是STING通路依赖的过程、可以直接结合cGAS并产生cGAMP。基础研究的不断深入,终能让人们登上一个又一个新的台阶。
谈到免疫还是让我打怵,对抗原性(antigenicity)和免疫原性(immunogenicity)的理解还不够多,有时总是搞混或直接画约等号。现在感觉他们的区别还是比较明显的:抗原性,这与抗体对应,偏重结合力,指结合某种细胞表面的受体或是B细胞活化后分泌的抗体的能力;免疫原型指激活机体免疫的能力,也就是抗原能激活T细胞或B细胞的能力,这个更为有效。换句话讲,抗原结合了T细胞或B细胞表面的受体还不能算有免疫原性,只有能起始后续获得性免疫的才算有免疫原性。
对于肿瘤细胞而言,他们由于自发突变率较高,比如皮肤癌就排在众多癌症的前列,容易合成一些“异样”的蛋白(俗称肿瘤相关抗原,tumor-associated antigens, TAAs),会被细胞列为外源物质而被攻击,因此我们说肿瘤细胞也有免疫原性。2014年11月University of Chicago的团队在Immunity期刊连续发表两篇姊妹文章,提供了新的解释,表明肿瘤细胞免疫原性的来源。结果显示STING在探测肿瘤细胞及诱发机体抗肿瘤效应的过程中扮演重要角色:一方面活化的STING可以诱导细胞因子的生成、活化靶向肿瘤的T细胞;[5]另一方面通过STING活化的免疫系统会提高肿瘤细胞对放疗的敏感性,这对癌症患者来说又是一福音。[6]
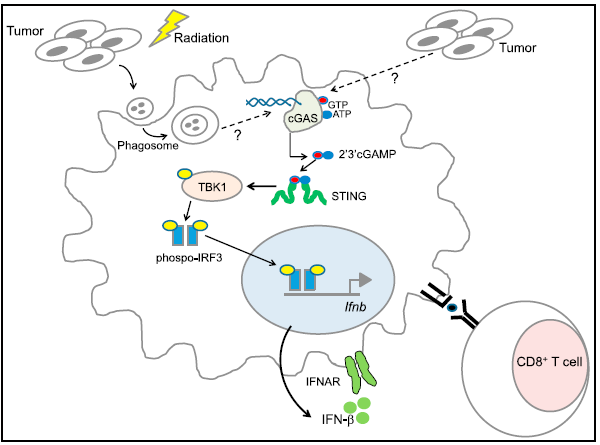
Spontaneous and Iatrogenic Tumor ImmunogenicityRequire STING and Type I IFN[7]
肿瘤细胞内部并非太平,细胞恶性增殖,内部定是险象环生。若是在此时自发地或人为地将肿瘤附近免疫细胞中的STING激活,促使细胞释放IFNβ、细胞因子,反过来增强免疫攻击力,甚至派送T细胞上场,对肿瘤细胞而言是致命的威胁。结合之前火热的immune checkpoint,若是细胞对药物产生耐药性或患者对此不响应,此时的STING是否会惊醒昏沉的免疫系统呢?Thomas的团队为此提供了重要的数据支持,他们发现体内的肿瘤微环境中肿瘤细胞中的DNA会被机体的抗原呈递细胞(antigen-presentingcells, APCs)识别,并激活内部的STING通路,释放IFNβ;而在STING缺失的小鼠体内却无法观察到此现象,这也进一步揭示肿瘤诱发先天免疫的分子机制。
无独有偶,由University of Chicago癌症生物学家Dr.Yang-Xin Fu和放射肿瘤学家Dr. Ralph R. Weichselbaum带领的团队与University of Texas陈志坚教授的团队合作也发现了类似的现象,此外还发现STING的活化可增强肿瘤细胞对射线照射时的敏感性,而非依赖之前报道的MyD88。昨日和领导交流的过程中恍然发现放疗的直接伤害之一就是破坏大部分细胞中的DNA,而这大部分细胞中的大部分就是数目最多、增殖最快的癌细胞。只是细胞中DNA有诸多的修复机制,之前主要抑制放疗后肿瘤细胞内的DNA修复,可明显增强放疗的疗效,如今可以通过STING提升DC攻击肿瘤细胞的能力,是比较新方式。



ThomasGajewski MD, PhD Yang-XinFu MD, PhD RalphR. Weichselbaum MD
STING and immunity
这个时候难免会遐想:若是肿瘤细胞中也有类似STING的功能,若是可以让肿瘤细胞自我分泌杀伤性武器该多好;(后来还真看到一篇今年年初发表的文章提到肿瘤细胞中的STING通路处于被抑制状态[8])是否可以引进一种复制能力更强的先天免疫细胞(比如NK、DC等),稍微有肿瘤倾向的也可以接受,用以毒攻毒的方式攻击肿瘤细胞呢?如此一来稍稍提升STING的活性就会产生强大的免疫反应。话说回来,STING的活化需要细胞质中有外源的DNA才行,那么免疫细胞又是如何获得肿瘤的DNA呢?目前看至少有如下几种可能:1,当实体肿瘤中部分细胞由于缺氧、营养耗尽而死亡时,DNA会与抗微生物的多肽LL37结合进行转运;[9]2,DCs可通过CLEC9A受体介导对死亡肿瘤细胞DNA的摄取;3,通过HMGB1(high-mobility group box 1)与肿瘤DNA结合后再与其他蛋白结合,实现DNA的转运;(Aduro称这个过程可能不依赖TLR)4,通过自噬体转运的方式摄入外源DNA。由此看来,被激活STING的细胞可能主要是DCs!(还有一个疑问:究竟什么样的DNA一定会被cGAS催化?是否像CRISPR那样需要某种特别的模序即可?)

A human dendritic cell (DC), which interacts with Tcells to initiate the immune response to tumor cells (Image: NationalInstitutes of Health)
到了这里,人们的视野更广——哺乳动物细胞中的DNA也可以被免疫细胞感知并激活STING通路,由此很容易让人联想到自身免疫疾病,如Aicardi-Goutieres syndrome(AGS)和全身性红斑狼疮(systemic lupuserythematosus, SLE)。有人基于这类疾病设计出一种合适的动物疾病模型——DNase II敲除的小鼠,这类小鼠无法正常降解溶酶体(lysosome)中冗余的DNA,导致DNA释放到细胞质中,可直接激活cGAS-STING通路,表现出严重的自身免疫疾病病症。有人发现敲除STING基因的小鼠对这类疾病有明显的缓解。这也表明STING通路长期处于活化的水平容易发展成过敏反应。此外,还有人发现在一些带有inflammatoryvascular-pulmonary syndrome 的患者体内的STING基因发生突变,导致组成型活化,过表达IFNs。现在看来,若是能找到一种STING的antagonist也是功德一件啊!(想起了雌激素受体ER,agonist到antagonist,再经down-regulator到modulator,这都是人们对事物不断认识的过程)
体内的实验不断的揭示肿瘤细胞中存在活化的CD8 T细胞,比如卵巢癌,乳腺癌,皮肤癌,胃肠癌 等等,而肿瘤微环境中T细胞的活化又离不开APCs的帮助。假设肿瘤微环境是无菌的(无病原微生物抗原的刺激),APCs的活化就需要内源性物质的刺激。T细胞与肿瘤天生势不两立,只是有时T细胞无法顺利的渗透(infiltration)到实体肿瘤内部。基因表达组学分析显示帮助CD8T细胞渗透到皮肤癌细胞中的蛋白多表达自IFNs诱导表达的基因,这也从侧面显示了IFN抗肿瘤的英雄角色,有材料显示IFN可帮助提升疫苗对癌症的疗效。其中释放IFN较多的一种免疫细胞是胞浆树突细胞(plasmacytoid dendritic cells, pDC),而STING在这类细胞中发挥关键的作用,如小鼠中STING或IRF3缺陷时无法激活T细胞排斥肿瘤的活性,ex vivo的分析显示肿瘤来源的DNA可被渗透入肿瘤的DCs感知,促使IRF3入核、诱导IFNs的表达。
其实还有很多关于STING的新发现,2015对STING而言又是一个丰硕之年,她再先天免疫、炎症、抗病毒、抗癌的过程中都有着无法忽视的作用,这也吸引了许多药物研发的工作者亲睐。一方面IFN直接与肿瘤内T细胞的渗透能力密切相关,利用小分子agonist活化STING通路是一个选择,而IFN的释放可以作为Biomarker;一方面肿瘤微环境中激活STING通路或释放STING活化后诱导产生的细胞因子,在临床免疫(联合)疗法中有着广阔的应用前景(这也是Novartis在这一领域抢先的原因之一)。目前已经开展的试验是将STING的agonist直接注射到肿瘤微环境附近(挺难的!如何找到肿瘤所在地?还是适合癌症晚期的患者?),in vivo的数据显示这种方法确实可以激活体内的STING通路,分泌I型IFN,活化CD8α/CD103 DCs,经淋巴激活抗原特异性的T细胞,经血液循环可在肿瘤之处募集活化后的T细胞,增强机体的免疫攻击能力。

Working model of the innate immune sensing of tumorsleading to spontaneous T-cell responses in vivo[10]
早在发现STING之前,CDNs就作为一种佐剂增强疫苗的免疫活化作用。在认识到STING通路的具体分子机制以后,CDNs和STING的天然配体cGAMP迎来一个春天。人们设计了许多类似结构的小分子,只是这些类似物的临床试验开展时间不久。后来发现STING的氨基酸序列存在多样性,对CDNs的结合能力不同,通路活化程度有所区别。[11]比如hSTING蛋白232位的精氨酸(R232)可以被细菌的CDNs激活,比如c-di-GAP,也可以被cGAS催化产生的cGAMP活化,而突变体R232H对细菌来源的CDNs并不敏感,但可被cGAMP活化,R293Q也是如此;hSTING-R71H-G230A-R293Q(HAQ)目前可以被所有的CDNs活化,然而在HEK293T中过表达时的活性却比野生型低,原因不明;在小鼠中,T596A的突变体也不响应CDNs的激活。诸如此类的单核苷酸多样性(single nucleotide polymorphisms, SNPs)日后临床试验中或许能为精准医疗提供参考。
介于DMXAA的临床试验的失败,人们在筛选初期便同时比较不同变异体的hSTNG和mSTING的活化情况。去年四月,Aduro与Jonhs Hopkins University合作发表一篇文章,介绍了他们设计的STINGagonist联合制剂可治疗对PD-1阻断耐药的肿瘤。[12]这个Lead小分子ML-RR-S2-CDA表现出较好的稳定性和亲脂性,与内源性和病原CDNs相比可明显活化STING信号通路。与DMXAA类似,直接注射入B16皮肤癌肿瘤小鼠模型中,可明显的清除肿瘤细胞,延长小鼠存活时间长达5个月,更重要的是对二次攻瘤有保护作用,在4T-1乳腺癌和MC26结肠癌模型中也观察到类似的现象。这些临床前的数据都显示ML-RR-S2-CDA直接注射的方式会得到最大的疗效,虽说该方法会限制临床上的应用,但试验显示在抑制本底肿瘤细胞的同时,也会抑制远处肿瘤细胞的生长,听说这和临床上使用T-VEC肿瘤病毒的疗法和TLR9 agonist CpG在放疗处理的non-Hodgkinlymphoma中的应用类似。
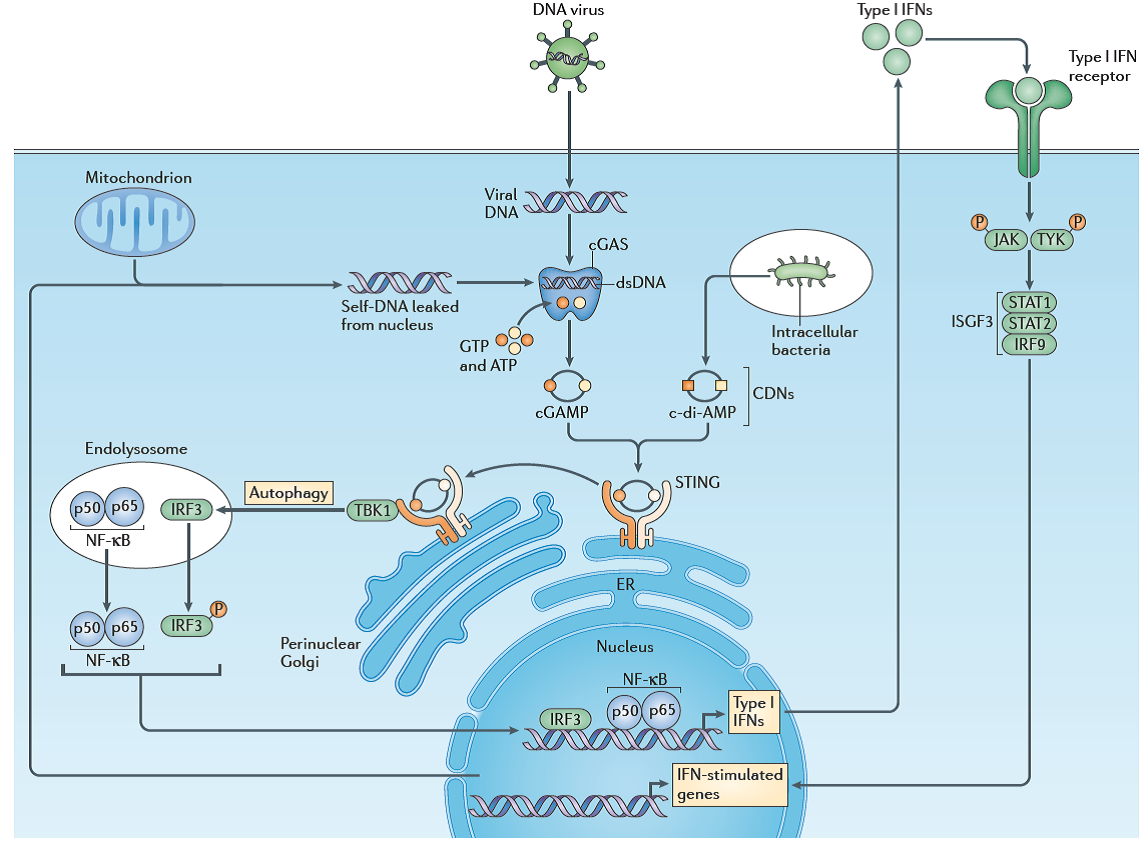
STING-dependent innate immune signaling [13]
越来越多的数据显示STING通路可通过提升免疫系统活性帮助细胞抵抗各种病原甚至癌症的发生,抗炎症、抗肿瘤药物研发由此又获得一次新的机会。磨刀不误砍柴工,详细了解STING相关的网络关联,更利于人们围绕该靶点设计药物,比如研究STING的转运、其他新身份、新功能、转录后修饰与调节等等,有数据显示STING是ER相关的转运复合体相关蛋白(ER-associatedtranslocon-associated protein, TRAP)的成分之一;其C端结构域中有多个Ser可被磷酸化,S366被磷酸化后会抑制对IRF3的激活,减少前炎症因子的释放,但对NFκB通路却无影响;酶组学分析发现自噬相关的激酶ULK1和ULK2可通过磷酸化S366来减缓STING的长期活性(听说TBK1也可以靶向该位点调节STING的活性),而ULK1和2在细胞不同饥饿情况下又可被AMPK和mTOR调节;还有人研究mciroRNA对STING活性的影响(主要在病毒学领域),目前还有很多工作亟待尝试。
STING and autoinflammatory disease
虽说暂时的炎症反应利于机体清理外源病原的入侵,物极必反,若是长时间活化机体的炎症反应便会导致自身免疫疾病。(这也是我们在孩子发烧时不能过早的吃退烧药,也不可以置之不顾的原因之一)有的国家有20%的的人们饱受自身免疫疾病的困扰,比如类风湿性关节炎(rheumatoidarthritis, RA),SLE和炎症性肠病(inflammatorybowel disease)等,只是大多数疾病的致病原因不明,但已知的是患者体内的炎症相关调节失调(inflammation-relateddisorder),抗核抗体(antinuclear antibody, ANA)水平较高,在血液中发现DNA甚至核小体(nucleosome)。这类内源的DNA可能来自凋亡、焦亡的细胞,目前认为与自身免疫疾病有关联,细胞在此难以分清敌我。
正常情况下细胞核外的DNA会被DNase处理掉,人和小鼠体内模型已经发现相关基因缺失易诱发自身免疫疾病,如缺失DNase 2基因的小鼠因无法降解细胞内多余的DNA,诱导IFNs高水平表达,一些细胞被巨噬细胞吞噬,小鼠未能顺利出生。后续有人同时敲除了STING(Tmem173)基因,并未发现凋亡细胞来源的DNA诱导的细胞因子风暴。在Trex1(three-primerepair exonuclease 1, a.k.a. DNase 3)缺失的小鼠中也发现类似现象,细胞因子在体内高表达,而STING在此过程中一样至关重要。(不知是否可以说STING是自身免疫疾病的帮凶?有种伴君如伴虎的感觉!)

STING in autoinflammatory disease [13]
反过来说,是否可以想象DNase可以负调节STING通路呢?有实验显示Trex1-/-的细胞复制过程中无法经G1期进入S期,在细胞质中积累大量的DNA;[14]Trex1-/-小鼠的BDMCs在G2/M期阻滞过程高表达细胞因子,侧面也显示Trex1在细胞内可能扮演一种看家基因的角色,基因组复制后清理细胞质中多余的DNA。[15]此外,还有一项研究显示Trex1无法降解经UV照射被破坏的DNA,而这类DNA却可以激活cGAS-STING通路。如此看来,细胞内应该有多种机制清理外源DNA或内源细胞质DNA,抑制这类分子激活DNA sensor相关的通路,其中也包括依赖STING的通路。理解这些细节,或许多药物研发会有帮助。
STING and Mitochondria DNA
谈到细胞质中的DNA,无法忽略线粒体DNA(mitochondrial DNA, mtDNA)。一般细胞中有数以百计(多达千余)的线粒体,而每个线粒体中约有2~10个拷贝的mtDNA,一种闭合环状的双链DNA(closed circular dsDNA),长约15,000个碱基对。[16]最近有人发现被病毒感染的细胞中由于线粒体压力导致mtDNA渗到细胞质中,激活STING通路,诱导细胞释放细胞因子。[17]咋一看来,这种凋亡的方式利于机体清理病毒的侵袭,可是mtDNA同时也会诱导细胞凋亡的发生。可是,体内每天有多达5百亿的细胞发生凋亡,如此一来,细胞必不可免的需要寻找高效的程序性死亡的方式处理这类事件,减缓对免疫系统的刺激。一些Caspase在这个过程中可以组织细胞因感知mtDNA而引发炎症反应。
简单来说,细胞发生凋亡需要先激活促凋亡分子(pro-death molecules)BAK和BAX,接着诱导线粒体外膜渗透(mitochondrial outer-membranepermeabilization, MOMP)、释放细胞色素c(cytochromec, cyt c)到细胞质中,与APAF1(apoptoticprotease-activating factor 1)和caspae-9 结合形成凋亡小体(apoptosome),激活下游的caspae-3、 caspae-7,靶向细胞蛋白诱发凋亡。尽管这些caspase蛋白在凋亡中有着至关重要的作用,可是对细胞死亡、体内清除细胞而言却并非必须,由此人们认为他们应该有其他身份。前两年人们发现caspae蛋白可以阻止mtDNA激活STING通路,具体分子机理不明,可能是摧毁了STING通路关键蛋白,也可能间接靶向了mtDNA。[18]当然,这种情况需要发生凋亡的细胞中表达STING蛋白,还好肝细胞中的STING表达量很低,要不然将会让人们很头痛,因为肝细胞中有太多的线粒体(每个细胞中多达数千个线粒体)。
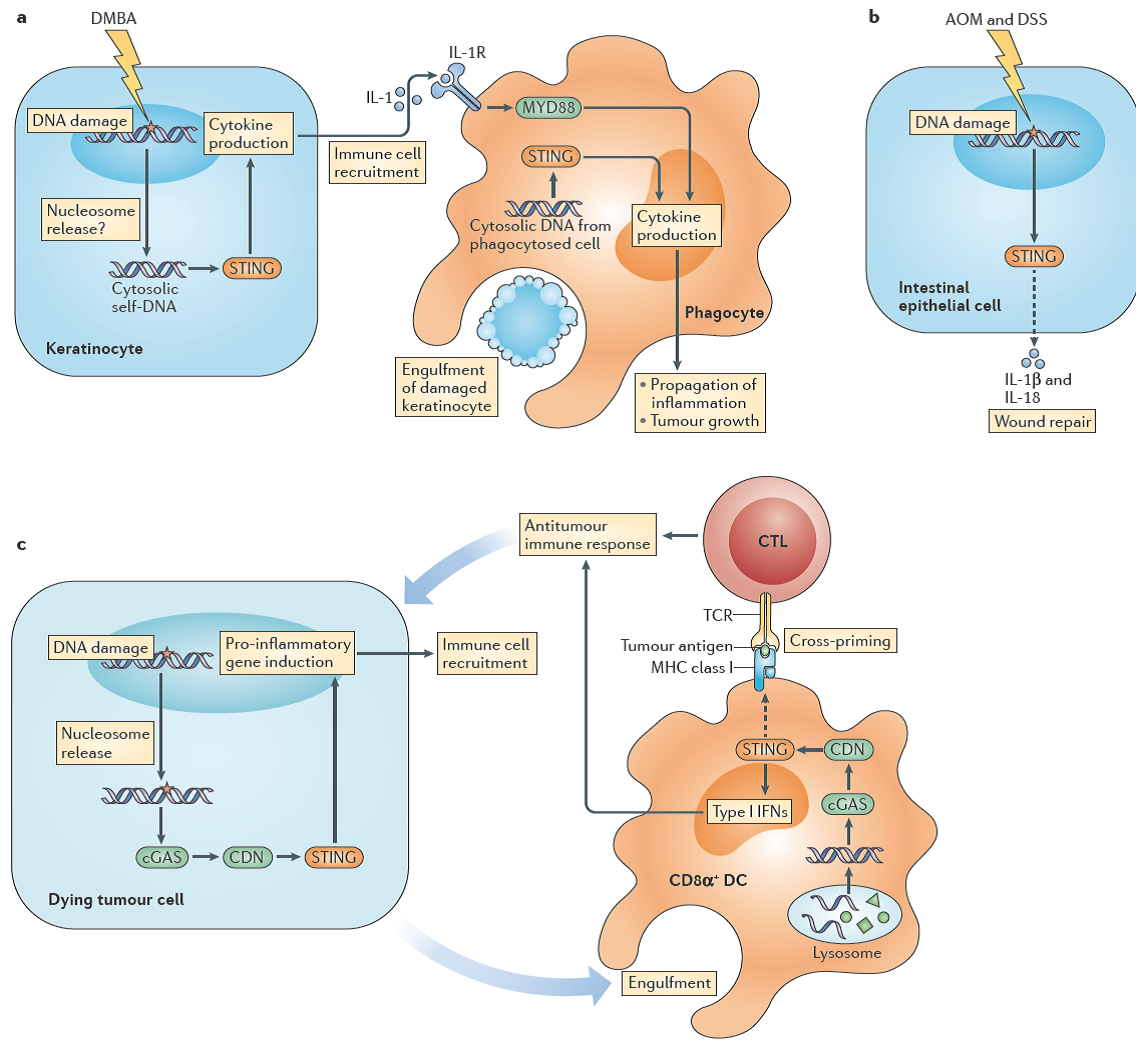
STING control of tumour development [13]
再次回头看cGAS-STING的通路,不再是DNA sensor那么简单,而是therapeutic target令人神往。多亏近五年几篇很有归纳性的综述,要不然向前后了解STING的通路要花费更多的时间。STING可以通过验证诱发癌症,可以通过细胞毒性T细胞提抗癌症;STING可以通过感知细胞质内DNA诱导细胞因子的释放,也可持续活化诱发自身性免疫疾病。这个时候人们会关注更细微的变化,完善STING相关的网络结构,因为越来越多的数据表示:STING通路无论是被诱导,还是被抑制,在癌症、抗病毒、抗炎症反应和疫苗研发等领域中都是一个非常具有前景的医疗靶点。
TJ At MeiHeKou city
Sept. 13, 2016 Tue
Mid-Autumn Day
Reference
7. Bronte,V., Tumors STING adaptive antitumorimmunity. Immunity, 2014. 41(5):p. 679-81.
https://blog.sciencenet.cn/blog-571539-1007532.html
上一篇:奔跑吧,少年!
下一篇:To Fanfan (XI)