博文
藜麦的细胞学研究
||
英国《自然》杂志8日在线发表的一篇植物科学论文,公布了首个藜麦高质量参照基因组。这项新成果将促进藜麦的遗传改良和育种策略,有望提高全球粮食安全。
此次,沙特阿拉伯阿卜杜拉国王科技大学研究人员马克·泰斯特及其同事,检测了智利沿海的藜麦品种基因组序列以及另外的藜属品种基因组序列,以表征藜麦的遗传多样性,理解藜麦基因组的演化。研究团队在进一步分析中,描述了其基因组数据以识别调控皂素形成的基因,皂素是藜麦籽壳中存在的一种苦味分子,必须在人类吸收之前去除。论文作者认为,他们发现的基因标记将可用于开发皂素含量低的无苦味或甜味藜麦商业品种。

藜麦是一种营养丰富、无麸质、血糖指数低的作物,所含人体必需的氨基酸、纤维、脂肪、碳水化合物、维生素能和矿物质达到出色的平衡,是唯一一种单体植物,可基本满足人体基本营养需求的食物。最重要的是,它能够在各种环境条件下生长。
这种植物其实已经有5000年至7000多年的食用和种植历史,在上世纪80年代,藜麦就被美国国家航空航天局用作宇航员太空食物。但迄今为止,藜麦仍属于一种利用不足的作物,为了扩大其在全球范围内的生产,还需要通过育种工作改善其农业性状。
本期小编精选了〔印度〕阿图尔·博汗格瓦,希尔皮·斯利瓦斯塔瓦 著;任贵兴 叶全宝 等译《藜麦生产与应用》[Quinoa Botany,Production and Uses],基于形态学、细胞学和基因组学来论述藜麦的分类学地位。
藜麦的细胞学研究
尽管对藜属植物的染色体数已进行过大量研究,但至今依然缺乏对其染色体核型的详细分析。据报道,藜属植物细胞的基本染色体数为 x = 8,基因组大小为x = 9 (Kawatani and Ohno,1950,1956)。只有Ambrina组的染色体数为x = 8 (Uotila,1973),这个组里的 C. ambrosioides(2n = 2x = 16)染色体数是一个典型的代表(Suzuka,1950;Giusti,1970)。Chenopodia 组中的染色体数为 x = 9,这个组进一步细分成 Cellulata,Leiosperma 和 Undata 3 个亚组(Risi and Galwey,1984)。
最近的细胞学研究确定了 C. quinoa 和 C. berlandieri 的染色体数为 2n = 4x = 36(Harbhajan,1961;Palomino et al.,1990,2008;Bhargava,2005;Bhargava et al.,2006;Kolano et al.,2012a)(图5.1)。Gandarillas(1979)报道,C. quinoa 具有混倍性,染色体数目为 2n = 18,2n = 27,2n = 36 和 2n = 45。Wang 等(1993)对藜属5个种下的9个类群的根尖体细胞染色体数进行了研究,其中C. berlandieri的 2 个栽培种和 C. quinoa 的 3 个栽培种,以及 2 个野生种C. berlandieri ssp.berlandieri 和C. hircinum 为四倍体。同一种下的各亚种染色体数目完全相同。在各种研究材料中频繁发现混倍性染色体。
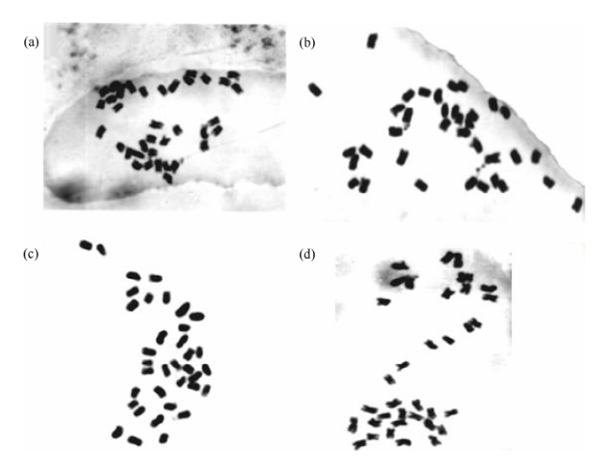
▲图5.1 体染色体 2n = 36,x = 9(转载自 Bhargava,2005)(a)C.quinoa PI 584524;(b)C.quinoa PI 596489;(c)C.quinoa CHEN 58/77;(d)C. quinoa PI 587173
尽管一些研究人员对藜属某些种的特定核型的特征进行了研究(Tanaka andTanaka,1980;Wang et al.,1993;Kolano et al.,2001),但依然缺乏对藜麦及其近缘种的完整细胞发育情形和详细染色体核型分析。
Catacora(1977)对 C. quinoa 的部分材料进行了核型研究,通过测定染色体臂长发现了其种间具有异源多倍性。以染色体长度和染色体长臂/短臂的值作为划分规则,将 C. quinoa(2n = 36)的 36 条染色体分成了 9 个组或 4 对同源染色体。Bhargava 等(2006)首次对藜麦(包括 7个材料)进行了全面的细胞遗传学研究,发现所有 C. quinoa 的材料核型特征均具有高度相似性,基于染色体臂长比值(最长臂/最短臂)将其分为两种核型,即比值小于 2.0 的为 1a 核型,比值大于 2.0 的为 1b 核型(图 5.2,图 5.3,表 5.1)。
基于臂长比值,对称指数(TF%)的变化为 43.9%(PI 584524,大部分非对称)~47.4%(CHEN 58/77,大部分对称)(表 5.1)。据报道,在所有分类群中都发现了双随体,其位置根据其在补体中相对大小的不同而改变(图 5.2,图 5.3,表 5.1)。

▲图 5.2 核型分析(转载自 Bhargava et al.,2006,经 Springer 许可)(a)C. quinoa PI 587173;(b)C. quinoa PI 584524;(c)C. quinoa PI 596498;(d)C. quinoa PI 510537;(e)C. quinoa CHEN 71/78;(f)C. quinoa CHEN 58/77;(g)C. quinoa CHEN 33/84;(h)C. berlandieri

▲图 5.3 染色体组型模式图(转载自 Bhargava et al.,2006,经 Springer 许可)(a)C. quinoa PI 587173;(b)C. quinoa PI 584524;(c)C. quinoa PI 596498;(d)C. quinoa PI 510537;(e)C. quinoa CHEN 71/78;(f)C. quinoa CHEN 58/77;(g)C. quinoa CHEN 33/84;(h)C. berlandierissp. nuttalliae PI 568156
◆表 5.1 藜麦 8 个类群核型分析(转载自 Bhargava et al.,2006,经 Springer 许可)
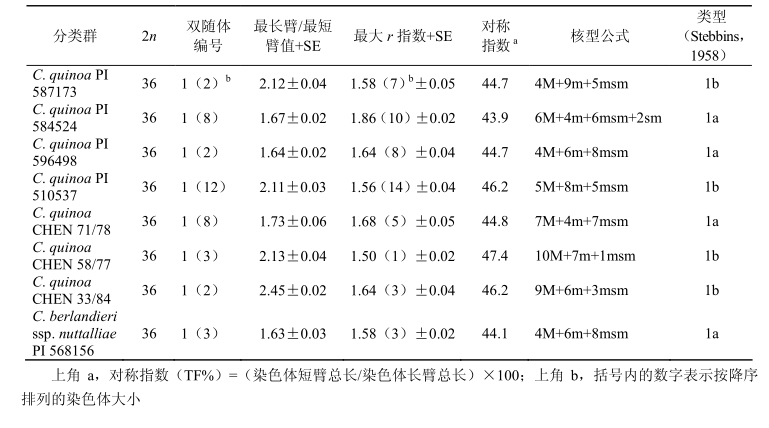

所有材料中的随体都具有相似特征,通常位于染色体中部(m)和近中部(msm),且都位于短臂上。第 1 号染色体的补体基因位置随着臂比率[1.18(PI510537)~1.56(CHEN 71/78)]变化,并不位于染色体中部和近中部,第 4、第9、第 18 号染色体的补体基因稳定地位于染色体中部(M 或 m)(图 5.2,图 5.3,表 5.1)。
第 10 号和第 13 号染色体上臂长比值变化范围为 1.0~1.86 和 1.0~1.78(图 5.2,图 5.3,表 5.1)。C. berlandieri ssp. nuttalliae 同样有随体染色体,随体位于第 3 号染色体短臂上(图 5.2,图 5.3,表 5.1)。
第 1 号染色体着丝点位于近中部,第 18 号染色体着丝点位于中部,与 C. quinoa 的其他材料的补体基因相似,都缺少 sm 基因。对称系数为 44.1%,染色体核型属于 1a 型(表 5.1)。
Bhargava 等(2006)的研究表明藜麦材料的核型存在微小差异,由于藜麦是起源于安第斯作物/杂草系统的单源物种,这样的结果在预料之中(Wilson,1990)。
然而,由自花授粉产生的染色体变异导致的藜麦核型微小差异(Risi and Galwey,1984),与形态特征变化(Risi and Galwey,1984;Wilson,1988;Bhargava et al.,2007)、蛋白质表达(Bhargava et al.,2005)及 RAPD 表达(Ruas et al.,1999)一致。


▲图 5.4 染色体模式图(转载自 Palomino et al.,2008,经 Springer 许可)(a)C. berlandieri ssp. nuttalliae cv. Chia roja 1-99;(b)C. quinoa cv. Barandales;(c)C. berlandieri ssp. nuttalliae cv.Quelite;(d)C. berlandieri ssp. nuttalliae cv. Huauzontle 8-01;(e)C. berlandieri ssp. nuttalliae cv. Huauzontle 7-01;(f)C. berlandieri ssp. nuttalliae cv. Chia roja 2-99;(g)C. berlandieri ssp. nuttalliae cv. Huauzontle 2-01;数字代表具有随体的染色体
通过 45S rDNA 荧光原位杂交法研究,发现了两条同源染色体的两个杂交位点,并只观察到了 1 个双随体(Kolano et al.,2001)。这两个 rDNA 基因座都具有转录活性,这意味着至少丢失了 1 个这样的基因座(Kolano et al.,2001)。Palomino 等(2008)还通过染色体核型分析进行了细胞遗传学方面的类似研究,鉴定了 C. quinoa cv. Barandales 和 C. berlandieri ssp. nuttalliae 的 6 个材料细胞学特征和基因组大小。从 C. quinoa 的核型公式、对称指数和相似的双随体形态都能够清晰的反映出 C. berlandieri ssp. nuttalliae 核型图与 C. quinoa 的其他材料基本相似。所有核型均具有 9 组等臂染色体,支持染色体数 x = 9 的观点(图5.4)。

各品种在染色体大小、基因组长度(GL)和随体位置上都有明显差异。被分析植物表现出两对具有随体的染色体,第一个随体定位在第 1 号染色体上,第二个随体的位置是变化的。C. quinoa cv. Barandales 的不对称指数或 TF=43.80%,与收集的关于 C. quinoa 的材料报道的 TF%值43.9%~47.4%相似(Bhargava et al.,2006)。

本文摘编自[印度] 阿图尔·博汗格瓦、希尔皮·斯利瓦斯塔瓦著;任贵兴 叶全宝 等译《藜麦生产与应用》[Quinoa Botany,Production and Uses]( 责编:刘 畅)第5章,内容有删减。
(本期编辑:安 静)

〔印度〕阿图尔·博汗格瓦,希尔皮·斯利瓦斯塔瓦 著
任贵兴 叶全宝 等译
北京:科学出版社 2014.11
ISBN 978-7-03-042333-7
《藜麦生产与应用》旨在为藜麦的生产和应用做一个全面的介绍。全书分为四个部分,共14章。第一部分对藜麦进行了简介,详细介绍了它的栽培驯化历史和地理分布。第二部分基于形态学、细胞学和基因组学论述了藜麦的分类学地位。第三部分主要讲藜麦的植物学特性和农业生产技术,包括植物学、抗逆能力、生产与管理、病虫害管理、育种及分子研究。第四部分阐述了藜麦的营养价值,并详细介绍了它的营养成分。

一起阅读科学!
科学出版社│微信ID:sciencepress-cspm
专业品质 学术价值
原创好读 科学品味
点击文中书名、作者、封面可购买本书。
https://blog.sciencenet.cn/blog-528739-1033955.html
上一篇:喜讯:SCIENCE CHINA Materials(《中国科学:材料》)被SCI检索
下一篇:大咖云集:“癌症研究前沿论坛”将在清华大学举行