博文
能流进化论——生物进化
||
从能量这一角度我们认为,生物是由非生命态能量传递介体演化而来,是非生命态能量传递介体能量传递能力不断升级的结果,其在自然界中的价值是驱动自然界中的能量流动,促进自然界中的能量分布由非均衡状态向均衡状态转变。既然如此,那么,生物的进化历程也应该是生物的能量传递能力不断升级的过程。生物的能量传递能力是由生物及其能量供体和能量受体组成的能量传递单元的能量平衡能力所体现,因而生物的进化历程,也可以认为是生物所属能量传递单元的进化历程。从生物的角度而言,生物的能量传递能力会影响能量供体和能量受体在自然界中的分布和数量。从能量供体和能量受体的角度而言,能量供体和能量受体的性质及其在自然界中的分布和数量决定了它们所需要的生物的能量传递能力。不同种类的生物之间往往呈现不同的能量传递能力,由此可推知,能量供体和能量受体决定了它们所对应的生物的种类、数量和分布。总之,能量供体、生物和能量受体三者之间相互影响、相互制约、共同进化。
(一)能量供体的演变
1.1能量供体的演变规律
我们假设在一个特定的环境中,几种生物和其“共享能量供体”之间已经达到了一种相对稳定的状态,即这些物种的能量传递能力刚好适合其“共享能量供体”对能量平衡能力的需求。若“共享能量供体”的丰度不再增加,随着“共享能量供体”的消耗,“共享能量供体”对能量传递能力的需求降低,物种之间对“共享能量供体”的竞争就会加剧。第二章已述,能量供体中的能量在被释放和传递过程中,若同时存在多种能量传递介体,那么能量传递能力相对较强的能量传递介体介导传递的能量要多于能量传递能力相对较弱的能量传递介体。因此,对于“共享能量供体”而言,能量传递能力相对较强的物种介导释放和传递的能量要多于能量传递能力相对较弱的物种。对于能量传递能力相对较强的物种,我们称之为“共享能量供体”的优势物种,对于能量传递能力相对较弱的物种,我们称之为“共享能量供体”的劣势物种。
相对于劣势物种,优势物种对“共享能量供体”中的能量表现出更强的竞争力,因而能够获得相对更多的能量来维持和提升自身的能量传递能力。对于有限的“共享能量供体”,优势物种释放和传递的能量越多,意味着劣势物种释放和传递的能量就越少,进而用于提升自身能量传递能力的能量也就越少。当劣势物种从“共享能量供体”中获取不到足以维持自身正常能量传递能力的能量时,其能量传递能力将会减弱。随着时间的推移,优势物种和劣势物种在自然界中所呈现的结果便是,优势物种作为适合“共享能量供体”的物种存活了下来,而劣势物种作为不适合“共享能量供体”的物种被淘汰(图1)。
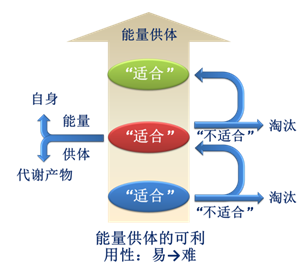
图1 物种与能量供体之间的相互作用。对于有限的能量供体,能量传递能力相对较强的物种,作为“适合”的物种会继续以此能量供体为能量供体;能力传递能力相对较弱的物种,作为“不适合”的物种,要么被淘汰,要么去介导竞争能力相对较弱的能量供体的能量的释放和传递。这个过程会导致新物种的产生。新物种自身及其代谢产物也可作为能量供体驱动新物种的出现。
现在假设:1)在对“共享能量供体”的竞争过程中,有的劣势物种逐渐适应并可利用另外一种物质作为能量供体,在此姑且称之为“新能量供体”;2)以“新能量供体”作为“共享能量供体”的物种之间对“新能量供体”的竞争程度相对缓和。这种情况下,可以以“新能量供体”作为能量供体的劣势物种将不会被淘汰,因为它们能够从“新能量供体”中获取能量来维持自身的能量传递能力。在适应“新能量供体”的过程中,有些物种会通过调整自身的能量传递能力,逐渐成为适合“新能量供体”的新物种,甚至有可能进化为“新能量供体”的优势物种。
此外,对于优势物种而言,其个体之间也会对有限的“共享能量供体”进行竞争。在竞争过程中,也可能会导致物种能量传递能力的改变,进而导致新物种的出现。总之,适合“共享能量供体”的优势物种一般会继续以该能量供体作为自己的能量供体,而不适合“共享能量供体”的劣势物种,要么被淘汰,要么选择另外一种物质作为能量供体。
劣势物种在介导“新能量供体”中的能量释放和传递过程中,也可能会导致新化学物质(代谢产物)的出现。对于新出现的化学物质以及由劣势物种演变而来的新物种而言,其能量也需要释放。也就是说,新出现的化学物质和新物种都有可能作为能量供体驱动新物种的出现。伴随着这一过程的发生和发展,能量供体的种类越来越多,生物的种类也越来越多。
随着生物种类的增多,物种之间的关系也越来越复杂,一些新的关系,如合作、捕食、共生、寄生等,也应运而生。例如,蚜虫以植物的汁液为食。它们把获取的汁液消化后,最终转变成一种叫做蜜汁的含糖物,然后排出体外。这种蜜汁又是蚂蚁喜欢吃的食物,也就是说,蚜虫的代谢产物是蚂蚁的能量供体。作为回报,蚂蚁会保护蚜虫,使其免遭瓢虫等对蚜虫有害的入侵者的攻击。对于这个例子,我们可以认为,蚜虫与植物之间是取食关系,而蚂蚁与蚜虫之间则是一种互惠互利的合作关系。
在合作关系中,有时候,一种物种对合作物种过于依赖以至于无法单独生存。例如,大栌榄树是生长在毛里求斯一个岛上的植物,其果实有坚硬的壳,一般动物无法将其打开,但是每个坚果必须被打开后才可以萌发。岛上的渡渡鸟可以将这种坚果整个吞下,然后,依靠它肌肉强健的、满是砾石的砂囊将果壳挤碎。种子随渡渡鸟的粪便排泄到地上后,便可以发芽,进而长成大树。大栌榄树对渡渡鸟的过于依赖的关系,使得渡渡鸟灭绝后,岛上再也没有长出新的大栌榄树。
七鳃鳗是一种吸附在鱼类身上以吸血为生的生物。它利用多排锋利的、向内弯曲的尖牙咬穿鱼类的皮肤,利用牙齿周围一圈圈的吸管紧紧地吸附在鱼的身体上,再利用舌头刺进鱼的伤口并吸食鱼的血液。对于这个例子,我们可以认为,七鳃鳗与鱼类之间是一种寄生关系,拥有血液系统的鱼的出现,驱动了七鳃鳗等以血液为能量供体的物种的出现。
此外,越容易被生物释放和传递能量的物质,往往越早以生物作为能量传递介体。相反,越不容易被生物释放和传递能量的物质,往往越晚以生物作为能量传递介体。对于能量被生物释放和传递难易情况相似的物质,越早在自然界中出现的往往就越早以生物作为能量传递介体,越晚在自然界中出现的往往越晚以生物作为能量传递介体。
1.2能量供体的演变
生物结构和功能的升级均需要消耗能量,且需要的能量主要来自于能量供体。既然如此,越容易被生物利用的能量,就越有可能成为生物演变阶段及诞生初期的能量供体。热能、光能等相对于储存在化学物质中的化学能而言,不但更容易释放和传递,还更容易获取和利用。光合细菌是最为祥知的在自然界中普遍存在的以光能作为能量供体的生物,是在厌氧条件下进行不放氧光合作用的细菌的总称,被认为是地球上出现最早的具有原始光能合成体系的原核生物。
热能、光能等容易释放和传递的性质,表明这类能量具有难以储存性,也就意味着一旦这类能量得不到持续性的供应,以其作为能量供体的生物的生物活动将难以得到保障。光合细菌结构和功能相对稳定的特性,说明它们具有储存光能的功能。这类生物采取的措施便是将光能转化为储存在化学物质中的化学能,该过程发生在生物内由光合作用和呼吸作用组成的“能量生产系统”中。光能、热能等经“能量生产系统”流入“能量可用系统”,进而转化为生物维持其生物活动所需要的自由能。
前文说过,在推动自然界中的能量由不均衡状态向均衡状态转变的过程中,生物的主要贡献在于将自然界中难以释放和传递的化学能转化为热能、光能等容易释放和传递的能量。既然如此,为什么光合细菌等生物又将这种容易释放和传递的热能或光能等转换成难以释放和传递的化学能呢?这是因为:
1)生物利用光能或热能将二氧化碳等物质转化为能量相对较高的有机物是为了更好的维持生物自身结构和功能的稳定,进而利于生物更好的呈现其能量传递能力。
2)生物的组装和增殖过程,也是将环境中分布不均衡的物质在生物内重新聚集和再分配的过程。物质都具有能量,因而物质在生物内重新聚集和再分配的过程,既是能量在生物内重新聚集和再分配的过程,又是环境中的能量由不均衡状态向均衡状态转变的过程。
热能、光能等可以作为一些生物的能量供体,然而,其分布的局限性也限制了以其作为能量供体的生物的分布。例如,以热能作为能量供体的生物主要分布于火山口及其周围区域、温泉等环境中,而以光能作为能量供体的生物只能分布在太阳光可以照射到的地方。再者,即使生物分布在这类能量供体附近,能量供体的供应也不一定会得到永久的保障。例如,生活在火山口附近以热能为能量供体的生物,当火山休眠或变成死火山后,这类生物将失去其能量供体。此外,环境的变动也会影响生物与能量供体之间的联系。例如,以光能作为能量供体的生物,在刮风、降雨、地震等因素影响下,可能会被从有光照的地方转移至光照较弱,甚至无光照地方。这种情况下,生物将难以获取足够的能量供体,进而难以维持自身结构和功能的稳定及施展能量传递能力。
从生物的角度看,当生物的结构和功能受到影响时,就会面临失去能量传递能力的风险。从自然界的角度看,只有具有合适能量传递能力的生物才能作为其需要的能量传递介体。在因能量供体匮乏致使能量传递能力受到影响的生物中,有的生物通过从周边环境中的物质分子中获取能量逐渐存活了下来,也就是说,作为其它种类物质的能量传递介体。例如,亚硝酸细菌、硝酸细菌、硫细菌、氢细菌及铁细菌等。它们通过氧化简单的无机化合物获取化学能,借助此能量同化二氧化碳进行细胞合成。这类以无机物作为能量供体的生物,被称作化能无机营养菌。化能无机营养菌在环境中普遍存在,可以作为其能量供体的物质主要有还原态的硫和氮化合物、二价铁离子、氢及甲烷、甲醇等一碳化合物。在介导物质分子中的能量释放和传递过程中,有些生物对其能量传递能力做了调整,以便更好的介导特定物质分子的能量释放和传递,进而更好的行使其在自然界中的价值。此时,这类生物对于自然界而言,仍然具有能量传递能力,仍然有存在的必要。
当物质分子作为能量供体时,储存能量越多的物质分子,生物在介导其能量释放和传递过程中,就越有可能获取更多的能量来提升自身的能量传递能力,进而提升自己在自然界中的价值。然而,储存能量越多的物质分子,其对能量传递能力的要求往往越高。对于生物而言,能量传递能力提升的过程也是自身结构升级的过程。换句话说,生物的能量传递能力与自身结构相辅相成。这便意味着,能量供体在驱动生物对其能量传递能力提升过程中,生物同样需要利于完成自身的构建及结构的升级,即在进化过程中,能量供体与生物相互影响、相互制约。前文说过,生物在介导能量供体中的能量释放和传递过程中,还需要从中获取维持自身结构和功能需要的能量。然而,生物的自身构建不仅需要能量,还需要物质分子。这些物质分子主要有两种来源:1)能量供体的代谢产物;2)环境。环境的不稳定性,一方面驱动了既可从能量供体中获取能量,又可从能量供体中获取自身构建所需物质的生物的发展;一方面又抑制了从能量供体中难以获取足够自身构建所需物质的生物的诞生和发展。结果便是,以生物的代谢产物、降解产物甚至生物作为能量供体的物种的出现。例如,以光能作为能量供体的光合细菌,其能量供体中缺乏生物自身构建所需要的物质分子,致使它们只能从周边环境中获取,这就使得它们对环境的依赖性更强,对环境的要求也更加苛刻;相反,以羚羊作为能量供体的狮子,狮子在取食羚羊的过程中,既可从中获取能量,又可从中获取维持生物自身构建所需要的物质分子,因而狮子在自身构建过程中,对周边环境中的物质分子的依赖性没有光合细菌那么强烈。
从以上分析中可以推知,越能同时满足生物能量和物质需求的物质,越适合作为生物的能量供体,也越容易驱动生物的进化。这也解释了为什么物质都具有能量,但不是所有的物质都可以选择生物作为能量传递介体,也不是任何地方都会有生物。
从能量供体的角度看,越适合作为能量供体的物质,以其作为能量供体的生物的种类和数量也就越多,生物之间的竞争也就越激烈。在生物之间对“共享能量供体”的竞争过程中,有的生物可以通过对自身结构组成的升级来提升自身的能量传递能力。例如,一些细菌长出了鞭毛,这使得其运动能力增强。运动能力越强的生物,接触能量供体的几率就越高,所呈现出来的能量传递能力也就越强。
这里需要说明的是,生物结构和功能的升级,不仅仅是自然界对生物能量传递能力需求驱动下的被动过程,还包含着生物的主动因素。随着生物的演变,生物结构和功能升级过程中所表现出来的主动性越来越显著。对于没有运动能力的生物而言,如果周边环境缺乏能量供体,那么这类生物的能量传递能力将会丧失,生物要么进入滞育状态,要么死亡。通过结构和功能的升级,有的生物具有一定的运动能力,能够去主动的获取能量供体,以维持自身的能量传递能力。在这个过程中,生物所表现出来的在能量供体获取方面的主动性,实质上是生物呈现其自然价值一种表现形式。这一表现形式随着生物的进化,越来越显著,特别是当生物的意识能动性在生物的生物活动中发挥的作用越来越大时,表现尤为显著。例如,我们人类不仅可以主动去获取能量供体(食物),还会主动培育和改良能量供体。
物质的结构决定其功能,生物也是如此。对于一条河流而言,如果河水的流量超过了河道的疏通能力,将会发生两种情况:一是出现一条新的河道;一是原来的河道变宽。河道变宽的区域一般是质地相对疏松的区域,这区域在河流的持续冲击下,更容易受到磨损。对于生物而言,当能量供体过量时,即生物可以持续从能量供体中获取充足的能量时,生物就像“河道”,要么形成一条新的“河道”,也就是驱动生物的增殖,要么增强“河道”的疏导力,也就是增强生物个体的能量传递能力。对于生物的这两种“策略”,从上一章对生物起源的推论中可以看出,这是生物与生俱有的属性。组成生物的各个组分就像河道的组成一样,有些系统“质地坚硬”,不能轻易更改。例如“能量生产系统”和“能量可用系统”,这类系统是生物维持其结构和功能所需要的最基本的组成,在几乎所有的生物中都存在高度的同源性。有些系统“质地疏松”,相对比较容易受环境的影响,在不同的物种中表现出显著的差异。例如,有的细菌有鞭毛,有的细菌没有鞭毛,有的细菌的细胞壁含大量的肽聚糖,有的细菌的细胞壁则含有极少的肽聚糖。
总之,生物在能量供体获取中所表现出来的主观性,本质上是生物各个组成部分在自然界的影响下被动改变的结果,是一种被动性中的主观性。
1.3能量供体的演变对生物进化的影响
有些生物,如蓝藻、硝化细菌等,能够利用热能、光能或化学能等将无机物转化成自身需要的糖类等有机物,这类生物称之为自养型生物。按照上文所述,能量供体的演变过程是,从最初的单纯的仅为生物提供能量的能量供体逐渐演变为既可提供能量又可提供生物自身构建所需物质的能量供体,可推知,自养型生物应该是地球上最早出现的生物。
《隐藏的动力:生物在自然界中的价值》一书认为,自养型生物在增殖过程中,由于一些原因,如所在环境中缺乏自养过程必需的某种物质,致使其自养过程不再是一个完整的过程,也就失去了将热能、光能或化学能等作为能量供体的能力。这类生物称之为“自养缺陷型”生物。该缺陷性致使此类生物在原能量供体的存在下难以维持正常的能量传递能力,也就面临着被自然界淘汰的风险。这类生物需要通过利用其它生物的降解产物或者代谢产物来弥补自身的缺陷。也就是说,这类生物需要依赖其它种类的生物才能存活。正如一个丧失劳动能力的乞丐,自身难以获取足够的能量供体维持自身的生存,但他可通过乞讨等方式获取维持自身能量传递能力所需要的能量供体。对于“自养缺陷型”生物而言,其演变方向主要为以下两种:
(1)向“自养-代谢双缺陷型”方向转变
如果“自养缺陷型”生物所在环境不止使生物的自养功能难以维持,而且其代谢功能也无法正常运行,这种情况下,有的生物为了尽可能的减少自身能耗,舍弃了自身组成中与“自养”和“代谢”两个过程相关的大量物质,只保留储存自身遗传信息的遗传物质,有的可能还会保留一些用以保护自身遗传物质的物质,以备“东山再起”。当这类生物遇到正常的生物后,自身遗传物质会借助正常生物组装出新的生物。这类生物称之为“自养-代谢双缺陷型”生物。假设正常生物需要从能量供体中获取总量为“E”的能量来维持自身的能量传递能力。当“自养-代谢双缺陷型”生物侵入正常生物后,它们会与正常生物展开对物质和能量的竞争。假设在竞争过程中,正常生物(寄主)获取的能量为“E1”,“自养-代谢双缺陷型”生物获取的能量为“E2”。当“E1”略小于“E”且远大于“E2”时,“自养-代谢双缺陷型”生物要么寄生在正常生物内,二者共存,要么被寄主抑制,甚至消灭掉(如抗性作物与病原菌之间的关系)。当“E1”远小于“E”和“E2”时,寄主获取的能量不足以维持自身的结构和功能,其能量传递能力就会降低,甚至丧失。“自养-代谢双缺陷型”生物也就是我们现在所说的病毒,需要依靠寄主才能维持其结构和功能的稳定性,进而维持其能量传递能力。总之,“自养-代谢双缺陷型”生物与寄主之间是一种既“合作”又“竞争”的关系,“合作”体现在二者均参与了寄主能量供体中能量的传递,“竞争”体现在二者对流经生物的能量的获取,当“合作”强于“竞争”时,二者共存,当“竞争”强于“合作”时,二者存一。
为了便于理解,我们可以把“自养型”生物、“自养缺陷型”生物和“自养-代谢双缺陷型”生物看作是一个传统的三口之家,“收入”是这个家庭的“能量供体”。“自养型”生物就像是一个家庭的“父亲”,既要获取“能量供体”,又要维持自身的“支出”。“自养缺陷型”生物就像是一个家庭的“母亲”,通过“父亲”获取的“收入”维持自身的“支出”,自己不需要主动去获取“收入”。“自养-代谢双缺陷型”生物就像是一个家庭的“孩子”,既需要依靠“父亲”或“母亲”帮其获取“收入”,又需要“父亲”或“母亲”帮其“支出”(如购买零食、玩具、衣服等)。对于父亲和母亲之间的关系,当父亲一个人的收入在满足自身支出后还有充足的结余时,那么父亲和母亲都会过得很好;如果结余不多,那么父亲和母亲的生活将会受到影响。对于父母和孩子之间的关系,当父母的收入在满足自身支出后还有充足的结余时,那么孩子也会过得很好;如果结余不多,那么孩子的生活将会受到影响。
(2)向“异养型”方向转变
有的“自养缺陷型”生物可能会通过利用其它生物的降解产物或代谢产物完成自我构建,这类生物称之为“异养型”生物。如果“自养缺陷型”生物所在环境中有足够的可以用于完成自我构建的物质,那么,有的生物可能会遗失自养过程所需要的物质分子,或由这些物质分子组成的系统,逐渐演变为彻底的“异养型”生物。对于这类生物而言,自养途径的失去,一方面减少了自养系统维护所需要的能耗,另一方面可以有更多的能量用于改进自身在其它功能上的结构组成,进而提升能量传递能力,包括运动能力、对能量供体的获取能力以及对能量供体中的能量进行释放和传递的能力等。
“异养型”生物在以化学物质作为能量供体的过程中,有的逐渐拥有了利用生物作为能量供体的能力。生物作为能量供体的优势在于,不止含有更多的可以作为能量供体的有机物,还含有可以被直接用来作为自身构建的物质分子。从生物的角度看,“自养缺陷型”生物若要增强自身的能量传递能力,就需要获取更多的能量,也就需要获取更多量的有机物,甚至更多种类的有机物。要实现这个要求,最有效的措施便是以现成的生物作为能量供体。有人可能认为生物的降解产物才能算能量供体,生物自身不能算。为了便于理解,这里对能量供体的界定再次说明一下,对于生物而言,能量供体和能量受体是独立于生物之外的物质。因此,当一种生物是另外一种生物的食物时,被食用的生物便是食用者的能量供体,而被食用生物在食用者体内的降解产物(非细胞内代谢,如在牙齿的咀嚼产物、胃液的消化产物等)则是食用者细胞的能量供体。
一般情况下,以生物作为能量供体的物种,其将化学能转化为热能的转化效率更高。这是因为,以生物作为能量供体的物种的运动能力往往比较强,运动量越大,消耗的能量就越多,能量供体的代谢就会越快,产生的热量也就越多。此外,这一过程也是被取食个体中的物质分子在取食者体内重新分配的过程,是自然界向能量均衡状态转变的需要,也是自然选择的结果。因此,以生物作为能量供体是生物的内部因素和自然因素共同驱动的结果。
通常情况下,出现越晚的或者能量越难以被利用的能量供体,其对应的生物类能量传递介体出现的时间就越晚。当生物作为能量供体时,生物自身的运动能力与能量释放难易性密切相关。运动能力越强的生物,往往越难以被捕获,进而以其活体形式作为能量供体的生物出现的时间也就越晚,数量和种类也随之越少。
对于不容易作为能量供体的生物而言,虽然以其活体形式作为能量供体的物种很少,但当该生物死亡之后,便会有很多物种以其尸体或尸体降解物作为能量供体。例如,对于草原上的狮子而言,很少有动物可以以其活体作为食物,然而,对于一头死亡的狮子,大至豺狼、食肉鸟,小至蚂蚁,甚至很多微生物都可以以这头狮子的尸体或尸体降解物作为能量供体。
综上所述,生物的能量供体在历史长河中的演变过程,被认为主要分为以下三个阶段(图2):
1)光能、热能等能量形式的能量作为能量供体;
2)化学物质作为能量供体,包括无机物(如还原态的硫和氮化合物)和有机物(如葡萄糖和乙酸);
3)生物作为能量供体,从单细胞生物(如大肠杆菌等)到多细胞生物(如草、兔子等)都可作为能量供体。
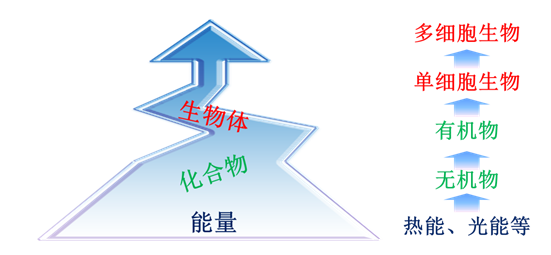
图2 以生物作为能量传递介体的能量供体的演变历程
(二)能量受体的演变
2.1能量受体的特征
为了便于对该部分的理解,我们先河水受体为例,分析一下河水受体的特性。从河流的实际情况不难得知:
1)河水受体与河水供体之间的地势差,既驱动了河水的流动,又限定了河水的流动方向,即自然条件下只能从地势高的地方流向地势低的地方;
2)在河水的流动过程中,从河水供体流出的水并没有全部流入大海,例如,有的河水以水蒸气的形式进入空气中,有的河水则会被河流两岸的生物利用等;
3)在河道坡度相同的情况下,河水受体与河水供体之间的地势差越大,河流流经的路线就可能越远,受益的生物就有可能越多;
4)湖泊和大海都是河水受体,但大海的水容量远远大于湖泊,一般情况下,河流中的水在进入湖泊后,还会继续流出,直至大海。
5)因为河水受体的存在,所以才会有地势差介于河水受体和河水供体之间的其它的河流汇入该河流,致使主河流的水不容易干涸,即主河流越能稳定的存在。
由此可见,河水受体自身的性质以及其与河水供体之间地势差的大小会影响河流的一些特征,如水流速、水流路径的长短等。对于能量受体而言,是否也有与河水受体相似的性质呢?
我们知道,化学反应的本质是电子的得失或转移。对于生物细胞而言,能量供体(如葡萄糖)在细胞内的代谢过程是由一些列的氧化还原反应组成。因此,能量供体中的能量在细胞内释放和传递的过程,也是能量供体中的电子在细胞内不断向氧化物物质传递的过程。基于此,能量受体也被称作“电子受体”,能量供体也被称作“电子供体”。例如,对于人体细胞而言,葡萄糖是电子供体,氧气是电子受体,葡萄糖中的能量经细胞释放和传递的过程,也是葡萄糖中的电子向氧气传递的过程。这里需要说明的是,葡萄糖中的电子向氧气传递的过程并不是指葡萄糖中的电子只传递给氧气,还有可能传递给其它与葡萄糖之间的电势差介于葡萄糖与氧气之间的氧化性物质。氧气因其相对较强的氧化性(接受电子的能力)限定了葡萄糖中的电子在细胞内的流动方向及传递途径。
本部分将重点探讨生物作为电子受体时的能量受体在历史长河中随生物进化而呈现的变化情况。对于生物而言,能量受体主要用于接受电子,为了便于理解,本部分中的“能量受体”以“电子受体”的表述形式呈现,“能量供体”以“电子供体”的表述形式呈现。在这里有一点需要说明的是,尽管很多化学物质具有接受电子的能力,但并不是所有可接受电子的物质都适合作为生物的电子受体。这是因为,生物自我构建及功能维持所需要的能量通常储存在ATP、GTP等“能量可用系统”中,而这些“能量可用系统”的生成并不是任何一个能够释放能量的化学反应都可以驱动的。也就是说,只有释放出足够能量的化学反应才有可能用以驱动ATP等“能量可用系统”的生成。一般情况下,电子供体和电子受体之间的能势差(电势差)越大,电子供体释放出的能量就会越多,细胞从中获取的能量也就越多,进而越有可能使自身的能量传递能力得到优化和升级。
此外,存在能势差的两种化学物质之间也不一定就会发生能量传递。正如上一章所举的例子,葡萄糖和1,3-二磷酸甘油酸两种物质分子均与ADP存在能势差,然而,一般情况下,葡萄糖不会直接将能量传递给ADP,而是先经过一系列的转换变成1,3-二磷酸甘油酸后,才将释放出的能量储存入ADP,使其转变成“能量可用系统”ATP。由此可见,只有存在合适能势差的两种物质之间才会发生电子的传递或转移,也才会发生能量的传递。此外,正如河水受体的存在使得其它河流得以汇入主河流一样,电子受体的存在同样使得生物中的其它能流得以围绕在电子供体和电子受体之间主能流左右,进而使得介导能量流动的物质分子得以聚集在一起,形成一个稳定的利于能量传递的体系。
通过上述分析可见,电子受体的功能在于:1)保证电子供体在细胞代谢过程中所释放的电子向电子受体方向传递,即规定了代谢电子的流动方向;2)驱动生物的各个组成系统的聚集,即保障生物结构和功能的稳定性和完整性;3)保证生物在介导能量传递过程中,可以从中获取充足的能量,产生足够多的“能量可用系统”。
2.2能量受体的演变
众所周知,氧气是几乎所有高等生物(如人类)唯一的电子受体。有意思的是,很多微生物,如地杆菌和希瓦氏菌,可以选择多种氧化物作为电子受体,如Fe(III)、Co(III)、U(VI)、Tc(VII)、延胡索酸、腐殖质等。不同的电子受体接受生物代谢产生的电子的方式往往不同。例如,当以水溶性的柠檬酸铁作为电子受体时(三价铁负责接受电子),有的地杆菌因细胞膜上缺少铁离子运输通道,致使铁离子主要在细胞壁和细胞膜之间的周质接受菌体代谢产生的电子;当以水难溶性的水铁矿(三价铁负责接受电子)作为电子受体时,由于水铁矿的分子量比较大而难以进入细胞,致使其主要在细胞外接受菌体代谢产生的电子。也有一些水溶性的氧化物(如延胡索酸等)或者小分子氧化物(如氧气),可以进入细胞内,因而主要在细胞内接受菌体代谢产生的电子。
一般情况下,电子受体越适合接受生物代谢产生的电子,就越容易驱动电子的流动,也就越有利于电子供体释放能量和传递电子,进而越适合环境向能量均衡状态转变的需求。此外,电子受体越有利于驱动电子在细胞内的流动,细胞中的“能量可用系统”就越充足,细胞也就越有充足的能量来驱动自身能量传递能力的提升。总之,对于生物而言,电子受体越合适,就越有利于生物能量传递能力的发挥,也就越有利于生物自然价值的呈现。对于电子受体而言,越容易接受生物代谢产生的电子,以其作为电子受体的生物的数量和种类就会越多,进而表现为各生物之间对电子受体的竞争越激烈。
水难溶性的电子受体主要存在于地球表面的土壤或淤泥中。这类电子受体的空间流动性(扩散能力)相对比较差,但在一个特定环境中的位置却相对比较稳定。水溶性的电子受体或分子量较小的电子受体主要分布在水环境或大气环境中。这类电子受体的空间流动性相对比较强,这使得其在一个特定环境中的分布相对更容易实现均衡。以难溶于水的铁矿石和分子量相对较小的氧气为例,铁矿石在地球表面的分布是不均匀的,而且自然因素(如风)难以影响其在地球表面的分布,这使得其在特地区域的分布又是相对稳定的。相比较而言,氧气分子的流动性就比较强,而且自然因素(如风)很容易带动氧气分子的流动,不过由于氧气分子的分子量相对比较小,氧气分子的布朗运动又使得其在空气中的分布很容易的再次实现均衡。
细胞复杂的结构组成说明,生物的起源过程需要在一个相对稳定的环境中发生。环境越稳定,组成生物的各种组分之间的联系就越不容易被破坏,也就越有利于生物的形成。由此可进一步推测,生物的起源过程及形成初期,所在特定环境内的电子受体也应该是稳定存在的。难溶性电子受体在特定环境中分布的均衡性和稳定性,使得其更利于非生命体系演变成生物。相比较而言,空间流动性更好的电子受体,因其在大空间尺度上分布更加均匀且更容易利用,因而更利于生物的增殖和传播。
可能有人会问,为什么现在大多数的生物,特别是陆生生物,会选择氧气作为电子受体,而不是其它氧化物?为什么这些生物只能以氧气作为唯一的电子受体?既然这是已经发生的事实,推测这一事实出现的原因主要有以下几点:
1)氧气可以通过自由扩散的方式进入细胞,使得其可以相对更容易的接受生物代谢产生的电子;
2)氧气接受电子的能力相对于其它氧化物而言更强,同时,对细胞造成的氧化毒害又相对较小;
3)“自养型”生物在二氧化碳等物质的固定过程中,实现了氧元素以分子形式从化合物中的分离(如植物的光合作用等),导致自然界中氧气含量的增加;
4)自然因素(如火山、雷电等)致使氧元素以分子形式从其化合物中分离,同样导致自然界中氧气含量的增加;
5)氧元素以分子的形式从化合物中脱离出来,也是降低其原来所在化合物能量的过程,符合自然界在能量流动方面的需求;
6)氧元素是地球表面含量最多的元素之一,为氧气的大量出现及更多的生物选择氧气作为电子受体提供了可能。
总之,氧气含量高、分布广、流动性好及其易于接受电子的属性,是越来越多的生物选择以其作为电子受体的根本原因,这同样也为生物的进化提供了驱动力。
2.3电子受体驱动的生物进化
如果一个环境不够稳定,生物各个部分之间的相互联系和相互作用就难以维持,也就不可能有生物的出现。水难溶性电子受体的空间稳定性使得组成生物的各个组分或系统之间更容易相互合作,生物的能量传递能力也能得以稳定的发挥。由此可见,稳定的电子受体环境是生物起源的必要条件,而这一环境便是富含水难溶性电子受体的环境。
然而,由于水难溶性电子受体的空间流动性比较差,随着生物的增殖,生物之间对电子受体的竞争必然越来越激烈。此外,随着电子的不断接受,特定环境中的电子受体对电子的接受能力越来越弱。空间流动性差这一特点,致使电子受体难以在该特定区域得到有效补充,这将进一步加剧生物之间对电子受体的竞争。在这个过程中,特别是当电子受体的电子容纳量快达到饱和时,如果环境中的电子供体含量充足,那么生物就不得不选择其它可以利用的氧化性物质作为电子受体。
在自然因素(如火山喷发、雷电等)及生物因素(如生物的代谢产物、降解产物等)的影响下,水溶性氧化物的数量和种类逐渐增多。这类氧化物空间流动性比较好,因而不适合非生命体系向生物的演变。然而,流动性好这一特点,使得可溶性氧化物得以在更大的空间尺度上分布,进而使得生物在特定空间里对电子受体的竞争相对缓和。这是因为,以水难溶性氧化物作为电子受体的生物,其只能分布在这些氧化物的表面,而对于具有相同电子接受能力的水溶性氧化物而言,电子受体的分布范围则是整片水域,因而生物的分布范围在理论上也是整片水域。就像一个开口的装满白砂糖的瓶子,假设瓶口的位置最多只能供100只蚂蚁取食,如果将瓶子里的白砂糖全部倒出来,并平摊在一张比瓶口面积大100倍的桌面上,对于相同数量的蚂蚁而言,其取食空间也就增大了100倍,也就意味着可以有更多的蚂蚁同时享用白砂糖。
再者,有的水溶性氧化物,特别是能进入细胞内的氧化物,相对于水难溶性的氧化物而言,更容易接受生物代谢产生的电子,这就使得一部分生长在水难溶性电子受体表面上的生物逐渐适应并能够利用这类水溶性氧化物作为电子受体。水溶性电子受体提高了生物代谢产生的电子在生物中的传递效率,进而使得更多的能量被生物释放和传递,这适合自然界对生物所属能量传递单元的能量平衡能力的需求,相应的生物的能量传递能力也就有存在的可能。此外,经生物释放和传递的能量越多,生物可获取的用以提升自身能量传递能力的能量也就越多。这些能量不仅用于生物自身结构组成的升级优化,还可用于提高增殖速度。生物数量的增加,表现为单位空间内生物对电子供体和电子受体的竞争加剧。在竞争压力下,有些生物通过对自身结构的升级,如产生鞭毛等运动器官,提升了自身的空间移动能力,即运动性,因而可以主动的移动至竞争环境相对缓和的空间生存。此外,也有些竞争能力相对较强的生物,可以利用自身的攻击系统去消灭或抑制竞争对手,以获取自身相对更大的生存空间。总之,水溶性氧化物作为电子受体,驱动了生物从地表或地下环境进入湖泊或海洋。在这个过程中,生物的增殖能力和扩散能力也都得到提升。此外,水溶性电子受体的多样性和更广的空间分布性还推动了生物在结构和功能方面的多样化升级,最终表现为物种的多样化。
在新出现的物种中,有些还保留着自养系统,只是因为周边环境的不合适,致使其自养系统一直处于休眠状态。当这类生物重新处于适合其自养系统正常运行的环境中时,自养系统有可能会被重新激活。例如,一些“光能自养型”生物因环境变化而进入了没有光的环境,在对新环境的适应过程中,一直保留着原来的光能自养系统,只是为了节约能量而将其光能自养系统处于休眠状态。在与电子受体的共进化过程中,有的生物重新回到了可以接受光能的环境中,如水面。充足的光照使得这些生物中的光能自养系统再次被激活,甚至有可能得到进一步的完善和升级。以光能作为能量供体的生物的种类和数量的增加,加剧了光能自养型生物之间对光能的竞争。有的生物可能通过提高光能利用率来减缓自身在光能获取方面的竞争弱势,有的生物则可能通过增强自身组织细胞的增殖来增加光接收面积,进而提升自身在光能获取方面的竞争优势。
除了自养系统,生物内也有可能存在一些对光照敏感的组分或系统。这类系统在受到光照后,有的功能可能会受到影响,有的则可能会呈现出新的功能。对于功能受到影响的系统,生物为了维持其正常的能量传递能力,就需要对其防御系统进行改造升级,以尽可能的降低或摒除光照对这些光敏感系统的影响。例如,有些化合物在光照条件下会发生共价键的均裂(共用电子均等地分配给成键的两个原子的过程),均裂后的两个部分各带有一个未成对电子,是一种活泼中间体。这类中间体又被称为自由基或游离基。如果自由基的含量比较高,就会对生物的健康造成影响。自由基的种类很多,比较常见有氢自由基、氯自由基、甲基自由基、氧自由基等。以超氧自由基为例,生物为了尽可能的降低超氧自由基对自身系统的伤害,会产生一种叫做超氧化物歧化酶的蛋白,该蛋白可将有害的超氧自由基转化为过氧化氢。过氧化氢在过氧化氢酶的催化作用下立即分解为无害的水。
对于激发出新功能的系统,如果该功能有利于提高生物的能量传递能力,将有可能会被生物加以优化利用。以视觉系统为例,视觉系统可以让生物更容易的探寻到能量供体或能量受体,以及躲避来自竞争对手的伤害等。鉴于视觉系统的这些优点,很多物种在进化过程中也在不断的对自身的视觉系统进行升级改造,使其更好的为自己服务。例如,乌贼的眼睛能够很好地处理海洋中浑浊的光线,使其在海洋中可以看得更清楚;比目鱼的两只眼睛位于同一侧,使其在侧躺在海床上时,更容易发现和捕捉到猎物,同时又可避免沙子进入眼睛;蜻蜓和苍蝇的眼睛是由数万只单眼组成的复眼,这种眼睛可以有效的计算自身与所观察物体的方位,从而有利于作出更快速的判断和反应。除了对与这类新功能直接相关的系统进行结构和功能的升级外,有的生物为了让这个新功能得以更充分的施展,还会驱动一些相关系统结构和功能的升级。例如,很多植物能够通过调节生长激素在自身不同组织器官的分布量,来改变组织器官相对太阳的朝向,进而使得自身可以获取更多的光照,该进化结果便是植物等生物所呈现出来的趋光性。
在水溶性氧化物可以作为生物电子受体的初期,水溶性氧化物的种类和数量繁多,但却还未出现一种占有主导地位的电子受体。此时,充足的电子供体和种类繁多的电子受体驱动了生物种类的爆发。这可能是寒武纪生物大爆发的原因之一。
在这些可被生物用作电子受体的水溶性氧化物中,有的丰度越来越高(如铁离子),同时,以其作为电子受体的生物的种类和数量也越来越多,该电子受体参与的能量传递单元的能量平衡能力也越来越强。这类电子受体逐渐成为生物的主导电子受体。与此同时,以主导电子受体作为电子受体的生物,也逐渐成为一个特定环境中共享“电子供体”的物种中的优势物种。在优势物种与劣势物种对“共享电子供体”的竞争中,劣势物种的劣势越来越明显,表现为能量传递能力越来越弱。共享“电子供体”的物种中的劣势物种,如果不能适应并利用其它物质作为能量供体,那么,随着时间的推移,该物种可能会被自然界所淘汰。反之,如果该劣势物种可以利用其它种类的能量供体,根据自然界会以当前最高效的方式使自身的能量向均衡状态转变可推知,这种能够适应其它电子供体的劣势物种有很高的几率会存活下来。随着利用主导电子受体的生物的数量和种类的增加,共享主导电子受体的物种之间对主导电子受体的竞争也日益激烈。
水溶性电子受体驱动着生物在地球表面更广的范围内分布和传播。在这个过程中,光能自养型生物的增殖和传播,使得空气中氧气的含量不断地增加。由于氧气能够通过自由扩散的形式进入细胞内,且接受电子的能力相对更强等优势,越来越多的生物能够适应并逐渐选择氧气作为电子受体。然而,氧气难溶于水的特性,致使其在空气中的含量比在水中的含量更高。总之,氧气作为生物的电子受体后,一方面使得生物的能量传递能力得到进一步提升,一方面其水难溶性也驱动着生物逐渐从海洋走向陆地。
基于电子受体的特征及其与生物之间关系的讨论,本书构建了一种电子受体驱动的生物进化模型(图3),即电子受体在生物进化过程中的演变历程被认为是:首先,难溶于水的、在特定区域内稳定存在的、相对难以被利用的氧化物(如水铁矿、磁铁矿等)是生物最初阶段的主要电子受体;随着水溶性氧化物的含量及种类越来越多,因其相对更容易接受生物代谢产生的电子,且生物对其竞争相对缓和等原因,越来越多的生物逐渐适应并选择这类氧化物作为电子受体;随着氧气的大量产生,因其氧化性更强、分子量更小、分布更均匀、更容易接受生物代谢产生的电子等特性,致使越来越多的生物逐渐适应并选择氧气作为电子受体。
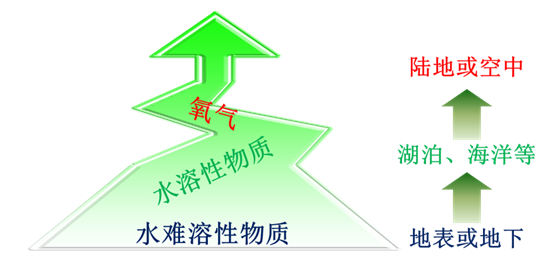
图3 驱动生物进化的电子受体的演变
该进化模型的构建之所以选择电子受体作为参照而非电子供体,主要是基于以下两件事实:
1)对于任一生物而言,电子供体(食物)的多样性往往多于电子受体的多样性。例如,人可以吃很多种食物,如肉类、蔬菜、水果等,但只能以氧气作为电子受体。当然,自然界中也存在着一些专食性的物种,如大熊猫只吃竹子、考拉只食用桉树叶子、蜗鸢专吃蜗牛等。然而,对于专食者而言,其专食性也是相对的,在偏好的食物匮乏时,它们也会选择其它食物,例如,野生的大熊猫也会吃草、野果、昆虫、竹鼠、牛羚、甚至附近村落里养的羊和垃圾堆的剩饭菜等。
2)电子受体的缺乏对生物造成的影响一般要比电子供体缺乏造成的影响更严重。例如,人可以三天不吃东西,但不能1小时不呼吸;冬眠中的青蛙可以几个月不吃不喝,但冬眠的环境中不能没有氧气等。
总之,生物的电子受体的演变历程,被认为是一个从水难溶性且相对难以利用的氧化物,到水可溶性且相对较容易利用的氧化物,再到氧气这一分子量更小且更容易被利用的氧化物的过程,同时,还是一个从种类繁多到种类单一的过程。
(三)生物的演变
根据前文所述,生物的价值体现于其在自然界中能量传递能力。因而,生物的进化历程本质上也是能量传递能力的进化历程。那么,生物的能量传递能力又体现在哪些方面呢?
对于生活在同一片草原上的羚羊和狮子而言,羚羊是狮子的能量供体,氧气是狮子的能量受体。氧气在草原上的分布相对均匀,而羚羊的分布却很不均匀,有的区域羚羊的数量很多,有的区域却很少,甚至没有。此外,不同季节或不同时期,羊群的数量也会有差异。例如,在生育高峰期或环境优越时,羊群的数量就会很多,如果遇到自然灾害(如干旱等),羊群的数量就会减少。再者,狮子可以很容易的获取氧气,但要捕获羚羊,则需要消耗一定的能量。由此可见,相对于能量受体,能量供体具有种类繁多、数量多变及分布不均等特点,这便使得生物获取能量供体的难度往往要比获取能量受体更难一些,说明生物对能量供体的获取能力要比对能量受体的获取能力更能体现生物的能量传递能力。基于此,本部分首先阐释生物对能量供体获取能力方面的演变历程。
3.1能量供体获取能力的演变
在电子受体驱动的生物进化模型中已有阐述,生物起源于水难溶性的电子受体表面。水难溶性电子受体的稳定性使得生物只能被动的从周边环境中获取能量供体。随着水溶性氧化物的大量出现及越来越多的生物可以利用这类氧化物作为电子受体,生物逐渐具备了主动获取水溶性电子受体和电子供体的能力。当人类出现后,在意识能动性的驱动下,生物不仅学会了制造和使用工具,还具有主动创造能量供体的能力。基于物种之间对能量供体获取方式的差异,生物的能量供体获取能力方面的进化过程被分为三个阶段:被动获取、主动获取和主动创造(图4)。

图4 生物对能量供体获取能力的演变历程
(1)被动获取
对于属于“被动获取”的生物而言,它们需要借助一些外界力量才能获取相应的能量供体,进而行使自己的能量传递能力。按照借助力量的作用又可将属于“被动获取”的生物分为两类:
1)依靠自然因素(如风、水流等)将生物携带至有能量供体的环境。例如,引发水稻瘟疫病害的稻瘟病菌,需要借助风力将孢子传播至新的水稻叶片表面,才能侵染新的叶片,进而从叶片中获取所需能量供体。
2)依靠自然因素将能量供体传送至它们身边。例如,贻贝、蛤蜊等,会黏附在经常浸没在海水中的固体表面,一旦黏附,其一生都将在那里度过,当海水上潮时,只需要打开身体的硬壳便可获取它们的能量供体,即海水中的浮游生物。
总之,这类生物可以认为是被动的行使自身能量传递能力的生物,也可称之为“靠天吃饭”的生物。
(2)主动获取
对于属于“主动获取”的生物而言,它们能够通过自身的组织、器官(如爪子、牙齿)等主动获取能量供体。也就是说,主动行使其能量传递能力。按照主动获取的能量供体的类型进行分类,可以将生物分为以下三种:
1)主动获取光能等“能量”型能量供体的生物。例如,植物的地上茎、叶部分有向光生长的特性,这有利于植物获得更多的光能,进而更好的生长。
2)主动获取“化学物质”型能量供体的生物。例如,蜂鸟、蜜蜂、蝴蝶等会主动飞到花朵上,以获取它们的能量供体(花蜜)。
3)主动获取“生物”型能量供体的生物。这类生物又可分为两类,一类是以死亡的生物作为能量供体,例如秃鹫、螃蟹、龙虾等。另一类是以鲜活的生物作为能量供体,例如,以细菌等微生物作为能量供体的线虫;以嫩草作为能量供体的鹿、牛、羊等;以鲜活动物作为能量供体的狮子、狼等。在以鲜活生物作为能量供体的物种中,有的学会了使用工具。一般情况下,制造这类工具的材料是直接来源于自然界,未发生化学性质的变化。例如,觅食的水獭总是在自己的腋下夹着一块石头,当它发现牡蛎、蛤蜊等贝类生物时,会用石头将这些贝类生物的硬壳敲碎,以获取夹在硬壳中的贝类肉体。
对于“主动获取”型生物,其对能量供体的获取能力主要体现于搜寻能量供体的能力及捕获能量供体的能力。例如,在搜寻能量供体方面,鹰和隼的眼睛可以看到1.6公里以外的一只兔子;北极熊的鼻子能嗅到4.8公里以外的海豹;大耳狐的耳朵能听到地下洞穴内小虫移动的声音等。在捕获能量供体方面,狮子的牙齿和爪子可以帮助它们杀死并撕开猎物;鹰和隼的喙坚硬且呈钩状,利于它们捕捉并撕开猎物;毒蛇利用牙齿将毒素注射入猎物体内以麻醉或杀死猎物等。由此可见,生物在能量供体获取能力方面的进化历程,也是生物组织或器官不断演变的过程。
(3)主动创造
对于属于“主动创造”类型的生物,主要是指我们人类。“创造”是我们人类主观意识能动性的体现,而主动创造的东西主要体现在工具和能量供体两个方面。
(a)对工具的创造。
1)利用自然界中的物质拼接而成的工具。这类工具是生物有意识的对原始材料进行一定程度的加工,但未改变材料的化学性质。例如,原始人类将木棍和锋利的石块捆绑在一起作为捕猎工具。这类工具主要用于获取和分割能量供体。
2)对自然界中的原始材料进行提炼、加工、改造后而制成的工具。这类工具不仅夹杂着大量主观因素,还体现了较强的意识能动性。例如,望远镜、雷达等探寻能量供体的工具,刀叉枪炮等捕获能量供体的工具,以及呼吸机、心电监护仪、磁共振等改善人体获取能量供体能力的工具等。
(b)对能量供体的创造。
1)品种的培育和改良。例如,超级杂交稻、海水稻、转基因大豆等的培育。
2)原始能量供体的深加工。例如,酒的酿造、食材的烹饪等。
总之,工具的制造和使用是人类区别于其它生物的主要体现。从望远镜的发明到卫星遥感的使用,人类在探寻能量供体方面的能力得到极大的提升;从刀、叉的锻造到枪、炮的使用,人类在捕获能量供体方面的效率得到极大的提高;从对能量供体的简单烹饪到各种产品的加工制造,人类在释放和传递能量供体能量的能力也得到极大的提升……
在这一过程中,除了与人类直接相关的能量供体中的能量得以释放外,环境中的很多能量相对较高的物质的能量也得到释放。例如,金银铜铁等各种材料的提炼,车、船、飞机、导弹、航母等工具的制造和使用,能源的勘测、开采和运输,能量的转换和输送等,这些过程都是耗能的过程,都伴随着能量的转化和释放。对于这些过程:
1)从大处讲,是自然界中的能量向均衡状态转变的过程。例如,对于一个铁矿而言,未开采之前,铁只分布在铁矿区域。人类将铁从铁矿里面提炼出来后,可以将其加工成各种产品,如菜刀、钉子、螺丝等。对于铁而言,铁元素在提炼产物中的分布比在铁矿中的分布更均匀,因而铁的提炼过程是能量由非均衡状态向均衡状态转变的过程(图5a)。再者,铁质产品的流通,致使铁矿中的铁元素得以在世界更广的范围分布,这也是能量由非均衡状态向均衡状态转变的过程(图5b)。此外,这一过程还伴随着大量化学能向热能的转化,利于自然界向能量均衡状态转变。
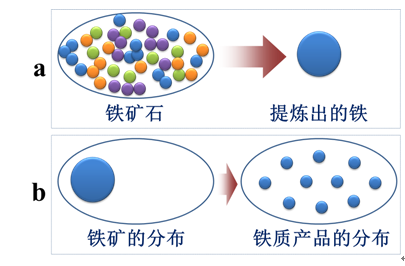
图5 人类活动对自然界铁元素分布的影响
2)从小处讲,是为了人类自身能量传递能力的提升。例如,车船的制造和使用可以让人类更容易的追捕猎物。在自然界中,很多能量相对较高的物质的储存量是有限的,例如煤炭、石油、天然气等。自然界对能量平衡能力的需求,使得能量平衡能力越强的能量传递单元中的生物个体或群体,就越适合自然界的需要。为了不被淘汰,生物个体或群体就需要提升自身的能量传递能力。自然界对人类的需求也是如此,人类能量传递能力的提升就需要消耗更多的能量,能源的有限性不可避免的引发人与其它生物之间、人与人之间、家与家之间、地区与地区之间乃至国与国之间对有限资源的竞争。
综上所述,工具的制造和使用,不仅大大提高了人类自身探寻能量供体的能力、获取能量供体的能力以及释放和传递能量供体中能量的能力,还促进了自然界中其它物质的能量释放,因而极大的推动了自然界向能量均衡状态转变的速率。此外,工具的制造和使用还加剧了人类之间及人类与其它生物之间对能量的竞争。在竞争过程中,人类的能量传递能力也在不断的改进和提升,尤其体现在对工具的制造和创造方面。
3.2遗传能力的演变
生物的能量传递能力不止体现在能量供体的获取方面,还体现在生物自身的遗传方面。如果一个物种的增殖能力提升了,对于特定环境中的能量供体而言,能量释放和传递效率也会随之提升。例如,对于微生物而言,如果所在环境中的能量供体充足,它们的增殖速度就会提高;如果环境中的能量供体缺乏,那么微生物的增殖速度就会降低,有的甚至会进入滞育状态。考虑到特定环境中的能量供体和能量受体的含量是相对有限的,单个个体能量传递能力低的物种,一般增殖能力会相对比较强,而单个个体能量传递能力相对较强的物种,一般增殖能力会相对比较弱。这是因为,单个个体能量传递能力相对越强的物种,所对应的能量供体的量就会越容易受其影响,在能量供体数量一定的情况下,物种之间对能量供体的竞争就越激烈,相互之间造成的伤害也就越大。例如,单只羚羊的能量传递能力要弱于单头狮子的能量传递能力,因而,正常情况下,羚羊的增殖能力要强于狮子。
3.3生物能量传递能力遵循的法则
在生物起源过程中,防御系统及攻击系统的形成决定了生物的利己主义本性。随着意识的出现及意识能动性在竞争中扮演的角色越来越显著,生物的利己主义在越来越多的方面得以呈现,而且影响也越来越显著。
对于微生物而言,在能量供体充足时,会疯狂的增殖,有的物种甚至还会分泌一些抑制剂或毒素来抑制其竞争对手的生长。对于这类生物而言,有能量供体就生长,没能量供体就休眠或死亡,不会去考虑如何更经济有效的利用能量供体。简单来说,如果微生物所在环境中有充足的能量供体和能量受体,能量就会自动开始从能量供体经微生物流向能量受体。如果由于一些环境因素的存在使得该物种的能量传递能力降低,那么,更多的能量供体就有可能被其竞争者获取,或者说,能量供体中的能量会更多的经其它物种传递。这个过程就好像是在电池的正极和负极之间连接一根导线,导线一连接,电子便会自动从正极流向负极;如果电池的正极和负极之间连接两根导电能力不同的导线(假设两根导线只有材质上的差异),一根铁线,一根铜线,由于铜的导电能力要优于铁线,可知单位时间内,从铜线流过的电子数量要多于从铁线流过的电子数量。由此可见,微生物就是一根复杂化的“导线”,微生物的这种行为也可以看作是一种极端的被动利己主义行为。
对于一些动植物而言,它们的利己主义行为中包含的主观因素往往会更多。例如,很多植物会通过花朵产生花蜜,用以吸引蜜蜂、蝴蝶等昆虫,昆虫采食花蜜的过程也是帮这些植物授粉的过程。在这个过程中,植物虽然付出了花蜜,但是却产生了更多的种子,进而会有更多的同种个体产生。个体数量增加的过程,是生物能量传递能力增强的过程,也是生物适应自然界的过程,因而这一过程是一种生物的主观利己主义行为。有些鸟类,如布谷鸟和燕八哥,会将自己的卵产在其它种类的鸟巢里,借助它们为自己孵卵及养育幼仔,这一过程也是一种生物的主观利己主义行为。
对于我们人类而言,意识能动性的参与使得利己主义行为更加的显著,影响也更大。例如,为了取火或制造家居,大片的森林被砍伐,最终导致土地的沙漠化;为了使农作物长的更好,进而获取更多的能量供体,除草剂、化学肥料及化学农药的大量施加,使得很多生物死亡,有得物种甚至灭绝;对教育资源、公共卫生资源、水资源、矿产资源等的竞争,使得人与人之间、家与家之间、地区与地区之间乃至国与国之间的矛盾不断地升级……
对于生物而言,利己主义行为是把双刃剑。以我们人类为例,一方面它让我们人类的能量传递能力越来越强,在与其它物种之间在能量供体或能量受体的竞争中,优势越来越明显,例如,枪支弹药的使用,使得老虎、狮子等猛兽在人类面前是那样的不堪一击;另一方面,如果该能量传递能力用在解决人类之间的内部矛盾中,那么,对人类造成的伤害也将会非常的惨痛,例如,一颗炮弹的爆炸可轻易的夺走成百上千人的性命。
此外,自然界驱动生物的产生,其目的是为了自身能量的分布更好的实现均衡。这说明,如果生物的能量传递能力适合自然界的需要,自然界会驱动生物的继续发展,如果生物的能量传递能力不适合自然界的需要,甚至有悖于自然界的要求,那么自然界就会抑制生物的发展。这就要求生物在行使一些功能时,不能只顾及自身的利益,还要考虑是否符合自然界的要求,尤其在能量流动方面的需求。换句话说,自然界中的能量流动是要遵守一定法则的。例如,森林有森林的能量流动法则,草原有草原的能量流动法则。草原的上的狮子本可以一天捕获很多头羚羊,然而,实际上,狮子只会根据自己的实际需要来捕获羚羊。如果生物不遵从自然界的能量流动法则,影响自然界正常的能量流动,那么生物的能量传递能力也将会受到影响,生物的存在性也会受到限制。例如,人类对森林的滥伐,致使更多的土地沙漠化,同时沙尘暴的爆发也更加频繁,这些变化使得我们人类的正常生活受到威胁。
总之,生物的利己主义行为,是自然界驱动的能量传递介体能量传递能力提升的结果,是生物被动生存中的主动表现。此外,自然界中的能量流动是一定法则下的能量流动,对于生物而言,遵循自然界能量流动法则的利己主义行为有利于驱动生物能量传递能力的升级,不遵循自然界能量流动法则的利己主义行为将会被自然界所抑制。
综上所述,生物的进化反映的是生物能量传递能力的进化,而生物的能量传递能力主要体现在能量供体的获取能力及自身遗传能力方面。因此,当比较不同生物的能量传递能力时,能量供体的获取能力及遗传能力可以作为主要参考指标。
https://blog.sciencenet.cn/blog-3479210-1309682.html
上一篇:能流进化论——生命起源
下一篇:能流进化论——“生物进化”还是“生物演化”
全部作者的精选博文
- • 再谈科研思路
- • 科研的思路应该是什么?
- • 面点统一的高等教育培养模式
- • 我眼中的王老师
全部作者的其他最新博文
- • 冲破枷锁不容易
- • 再谈科研思路
- • 科研的思路应该是什么?
- • 再读吉布森的生态光学
- • 赋予冰冷的数字以生物学意义
- • 大道至简