博文
【文献阅读】凝集素受体样激酶通过酪氨酸磷酸化LORE调节植物免疫
||
文章题目
【Tyrosine phosphorylation of the lectin receptor-like kinase LORE regulates plant immunity】
所属期刊
The EMBO Journal ,11
关于作者
刘俊课题组所属中科院微生物所,一直致力于研究植物抗病免疫的分子机制,在植物的凝集素类受体激酶介导的先天免疫以及水稻抵抗稻瘟菌侵染的基础免疫方面取得了一系列重要成果
本文重点
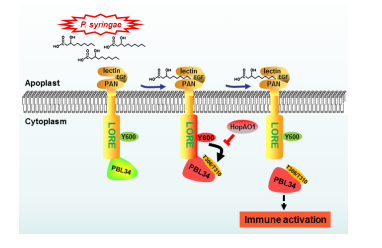
![]()
mc-3-OH-FAs可以在LORE的Y600处诱导其磷酸化
三个受体激酶PBL34、PBL35和PBL36参与了LORE激活后的下游信号传递,且均能被受体LORE磷酸化
LORE上Y600的磷酸化对PBL34、PBL35和PBL36的磷酸化至关重要
丁香假单胞菌在侵染过程中可以分泌一个磷酸酶HOPAO1,该酶可以直接结合并靶向LORE、去除LORE受体上Y600的磷酸化,进而抑制LORE介导的植物免疫
解决的科学问题
LORE在拟南芥中介导信号响应3-OH-C10:0的分子机制
相关背景知识介绍
中链3-羟基脂肪酸(mc-3-OH-FAs):广泛存在于革兰氏阴性菌中,最近被证明是拟南芥中新的PAMPs;
LORE:拟南芥中的PRR,是一种G型凝集素受体样激酶,可识别mc-3-OH-FAs,随后产生免疫应答;
结果一:LORE的酪氨酸磷酸化是3-OH-C10:0激活免疫应答所必需的

[体外激酶活性测定]→LORE是真正的激酶,K516和D613是激酶活性的两个重要位点.
图AB[激酶抑制剂A23(酪氨酸抑制剂)K252a(S/T/Y非特异抑制剂)处理拟南芥,并检查是否有3-OHC10:0引发的ROS爆发】→K252a和A23都强烈抑制了3-OH-C10:0引发的ROS爆发,A23更是废除了这一反应→作者接着检查了PTI标记基因FRK1和NHL10的转录水平发现也是相同情况→LORE介导的PTI需要激酶活性和酪氨酸磷酸化。
体外激酶检测显示即使在酪氨酸磷酸化被A23抑制的情况下,LORE的丝氨酸/苏氨酸磷酸化仍然存在。但A23完全抑制3-OH-C10:0引发的ROS爆发的观察则表明酪氨酸磷酸化是用于LORE介导的3-OH-C10:0识别的非常重要的位点。图CDE进一步确认。
结果二:3-OH-C10:0在Tyr600诱导LORE磷酸化

【LC-MS/MS】→Y600作为LORE的磷酸化位点,在水稻和拟南芥中保守→作者接着利用实验进行确定,并且作者通过实验证明3-OH-C10:0可以进一步促进了Y600磷酸化。
结果三:LORE的Y600磷酸化是其介导3-OH-C10:0的识别所必需

(图E)与先前研究一致,在3-OH-C10:0反应中,lore突变体明显的破坏了ROS爆发,回补实验则恢复了异常表型;但是在lore突变体背景下的激酶突变体的ROS水平则明显降低,作者利用实验证明这个现象并非是由于激酶mRNA水平的不稳定造成的,因此得出结论Y600磷酸化对于LORE介导的3-OH-C10:0的识别和PTI反应至关重要。(图FG)作者又利用RT-qPCR检测拟南芥叶片中PTI反应基因FRK1和NHL10在3-OH-C10:0处理后的转录水平重复验证→LORE的Y600磷酸化参与了3-OH-C10:0感知的下游信号传递
结果四:LORE 与PBL34 /35/36有关联
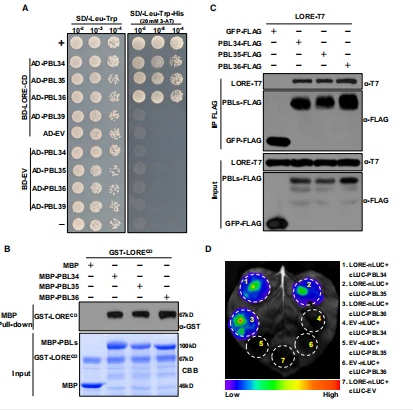
图A【co-IP、 LC-MS/MS】→寻找LORE的互作蛋白→因为之前有报道过VII RLKs和植物免疫信号相关,因此作者重点筛的互作蛋白是定于质膜或者属于RLCKs家族蛋白
【Y2H】验证确定了PBL34可以和LORE互作→由于PBL34有两个同源蛋白PBL35和PBL36,同时也属于RLCK VII亚家族所以作者也通过酵母双杂证明PBL35/36也能和LORE互作。
图BCD【pull-down ,co-IP,luc】针对上述结果再次重复确认
【农杆菌转化观荧光】确定PBL34/35/36定位于质膜上。
总结:这些结果表明,PBL34/35/36是LORE复合物的组成部分,参与LORE介导的拟南芥免疫反应
结果五:pbl34/35/36三突表现出对3-OH-C10:0引起的反应不敏感
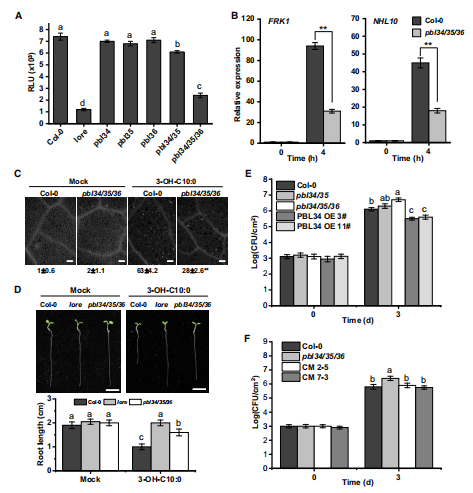
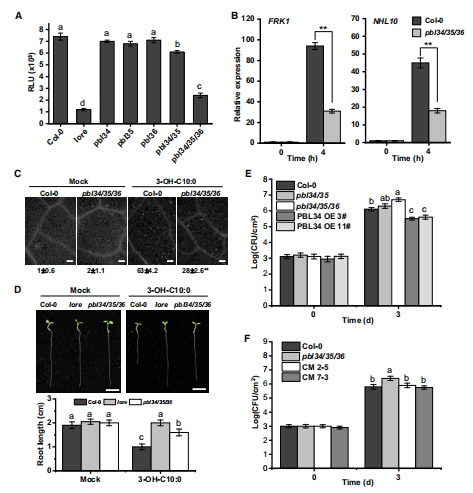
由于这PBL34/35/36是同源蛋白,因此作者想看一下他们在3-OH-C10:0诱导免疫反应的角色上是否功能冗余→作者通过检测在pbl34,35,36单突变体,pbl34/35双突变体和pbl34/35/36三重突变体对3-OH-C10:0的PTI反应中的ROS爆发、PTI标记基因FRK1和NHL10的转录水平、胼胝体沉积以及生长状况→PBL34/35/36的确存在功能冗余
作者还利用了其他的PAMPs分子重复这个实验→PBL34/35/36对3-OH-C10:0具有特异性。
结果六:LORE可以磷酸化PBL34
因为之前已经证明了LORE是激酶,并且LORE可以和PBL34相互作用,因此作者猜想LORE可以直接磷酸化PBL34
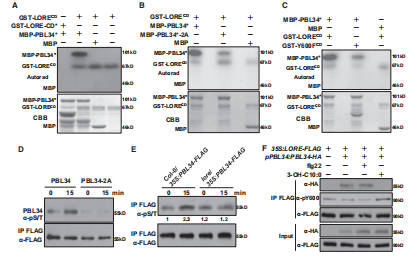
图A【vitro kinase assays】证明LORE可以磷酸化PBL34,PBL35,PBL36
【LC-MS/MS】对PBL34中磷酸化位点的检测发现T306/T310被LORE-CD磷酸化→LORE介导的T306/T310磷酸化激活了PBL34的激酶活性。
图B【模拟去磷酸化】为了证实T306和T310是PBL34的主要磷酸化位点,我们将这两个位点突变为丙氨酸→T306和T310是PBL34中被磷酸化的两个主要位点
图C LORE在Y600上的突变降低了其反式磷酸化活性→PBL34的磷酸化依赖于LORE的Y600磷酸化
图D.作者在拟南芥原生质体中表达PBL34和PBL34-2A,然后用3-OH-C10:0处理原生质体。3-OH-C10:0处理增加了PBL34的磷酸化,而PBL34-2A则没有→表明T306/310在响应3-OH-C10:0时被磷酸化
图E作者在Col-0和Lore背景下将35S:PBL34-FLAG引入→3-OH-C10:0诱导的PBL34磷酸化依赖于LORE
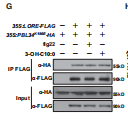
图F →在没有3-OH-C10:0的情况下,PBL34被LORE免疫沉淀,而3-OH-C10:0(但不是flg22)处理促进了PBL和LORE解离
图G →3-OHC10:0触发的PBL34与LORE的解离依赖于PBL34激酶活性,因为PBL34K180E的激酶突变体没有与LORE解离
总结:LORE与PBL34,PBL35,PBL36的形成一个复合体,在3-OH-C10:0的存在下复合体会解离
结果七:PBL34在T306/T310处的磷酸化是LORE介导的3-OH-C10:0的识别所必需的

结果八LORE被细菌酪氨酸磷酸酶HopAO1靶向
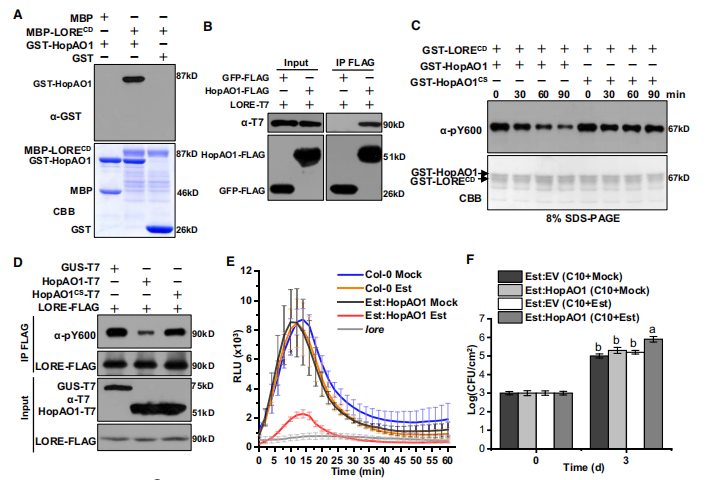
图AB【pull down,co-IP】HopAO1 可以和 LORE-CD相互作用
图C【 vitro phosphatase assays】HopAO1可以在体外对LORE的Y600去磷酸化
图D在体内又进一步验证
图E 【co-IP】HOPAO1抑制3-OH-C10:0触发的ROS爆发.
图F 抑制3-OH-C10:0诱导的抗病性。
总结:HopAO1的靶点是LORE,可以通过去磷酸化LOREY600从而抑制PTI。
https://blog.sciencenet.cn/blog-3420446-1219155.html
下一篇:效应因子触发植物免疫的广谱图