博文
Nature Genetics:多个作物驯化过程中种子休眠基因的平衡选择
||
Parallel selection on a dormancy gene during domestication of crops from multiple families
First author: Min Wang; Affiliations: Institute of Genetics and Developmental Biology, Chinese Academy of Sciences (中科院遗传与发育生物学研究所): Beijing, China
Corresponding author: Zhixi Tian (田志喜)
Abstract
Domesticated species often exhibit convergent phenotypic evolution, termed the domestication syndrome (驯化综合征), of which loss of seed dormancy (休眠) is a component. To date, dormancy genes that contribute to parallel domestication across different families have not been reported. Here, we cloned the classical stay-green G gene from soybean and found that it controls seed dormancy and showed evidence of selection during soybean domestication. Moreover, orthologs in rice and tomato also showed evidence of selection during domestication. Analysis of transgenic plants confirmed that orthologs of G had conserved functions in controlling seed dormancy in soybean, rice, and Arabidopsis. Functional investigation demonstrated that G affected seed dormancy through interactions with NCED3 and PSY and in turn modulated abscisic acid synthesis. Therefore, we identified a gene responsible for seed dormancy that has been subject to parallel selection in multiple crop families. This may help facilitate the domestication of new crops.
驯化物种经常存在协同表型演化,术语叫做驯化综合征,种子休眠丢失便是其中一个特征。目前来说,作用于不同科属植物平衡选择的休眠基因还未见报道。本文,作者在大豆中克隆到了一个保持绿色的G基因,结果发现该基因还控制了种子的休眠,并且在大豆的驯化过程中受到了选择。此外,G基因在水稻和番茄中的同源基因同样在驯化过程中受到了选择。转基因试验证实了G基因在大豆、水稻和番茄中具有保守的控制种子休眠的功能。进一步的研究显示G蛋白通过与NCED3和PSY互作调控脱落酸的合成,进而影响种子的休眠。因此,作者鉴定到了一个控制种子休眠的基因,该基因在不同的作物物种中受到了平衡选择。本文的研究将有助于促进新作物物种的驯化。
Introduction
驯化是植物育种最早的形式,同时也是人类历史上最重要的一次技术创新。在驯化过程中,诸如顶端优势的增加、种子落粒性的丢失以及种子休眠的减少等一类被叫做“驯化综合征”的共同性状在不同作物间受到了趋同选择。这就带来了一个十分有趣的问题,即是否存在控制驯化综合征的基因在不同作物驯化过程中受到了平衡选择。早期利用限制性片段长度多态性RFLP标记进行的比较作图显示高粱、水稻和玉米经历了趋同选择,并且某些驯化综合征,比如种子大小、落粒及日长不敏感型开花是由少数的几个数量性状位点QTL所控制的,并且这些QTL在这三个物种之间非常一致。然而,进一步的 QTL作图及克隆分析显示大多数已知的某个特定性状的驯化基因对于每个驯化系来说是特异的。尽管已经鉴定到了一些在不同物种间受到平衡选择的基因,但都是在同一个植物科属内的物种。因此,是否在亲缘关系较远的作物之间存在驯化综合征性状的平衡驯化还不清楚。
种子休眠的丢失是一个典型的驯化综合征。在野外条件下,种子休眠的丢失是十分有害的,因为所有的种子将在第一次下雨或者气候变暖之后全部发芽。因此,休眠通过延迟或者去同步植物发芽,来保证至少有部分的种子能够在适合的环境条件下发芽,得以存活。对于驯化的植物来说,所有种子同步发芽是能够保证后期作物的同时成熟,并有利于作物的田间管理。植物种子休眠由十分复杂的机制调控,涉及到形态、生理和物理结构等多个方面,难以检测和比对休眠相关的特征。因此,到目前为止仅有少数的几个与作物驯化中休眠相关的基因被鉴定出来,而且这些基因在不同科的物种中并不存在平衡选择。本文中,作者克隆了大豆的一个保持绿色的基因,并偶然发现该基因与种子的休眠相关。此外,该基因在不同科的物种间功能十分保守,并且在驯化过程中受到了平衡选择。
Results
1. GWAS鉴定大豆绿色种皮基因
大豆是一种重要的产油和植物蛋白的作物。大豆种子的种皮颜色在不同的种质资源间十分多样化,包括黑色、黄色、绿色、棕色,以及条纹色彩(Figure 1a)。大豆的绿色种皮是由三种保持绿色位点通过不同的机制所控制的,其中G位点主要控制种子种皮的颜色,而其它的两个位点控制其他器官的颜色。为了鉴定G基因的遗传基础,作者选择了重测序品种中带有绿色或黄色种皮的栽培种大豆(Table S1),并进行了WGAS分析。基于一个标准的混合线性模型,作者在1号染色体上鉴定到了一个显著相关的单核苷酸多态性位点SNP1128991(Figure 1b)。该SNP位于大豆一个编码CAAX氨基末端蛋白酶蛋白Glyma.01G198500基因中。基因转录水平的研究显示,相对于绿色种皮大豆中的G核苷酸(G等位基因),黄色种皮大豆中的A核苷酸(g等位基因)会导致一个可变剪切位点,并产生一个提前终止密码子(Figure 1c & Figure S1)。g中的突变会导致该基因缺少最后的跨膜结构域(Figure 1d),但并不影响该基因在叶绿体中的亚细胞定位(Figure S2a)。
为了进一步确认Glyma.01G198500基因是否是控制大豆种皮绿色的基因,作者将具有G等位基因并且种皮为绿色的Kuaiqingpi的Glyma.01G198500基因核苷酸序列全部转入到具有g等位基因并且种皮为黄色的栽培种D50中。两个独立的T3代转基因,即纯合转基因株系大豆的种子都表现出绿色的种皮(Figure 1e),并且在种皮中有残留的叶绿素(Figure S2b)。与此同时,在D50中过表达Kuaiqingpi的Glyma.01G198500基因编码序列能够获得类似的结果(Figure 1e & Figure S2b)。以上的试验结果说明Glyma.01G198500基因就是大豆中种皮表现为绿色的控制基因。
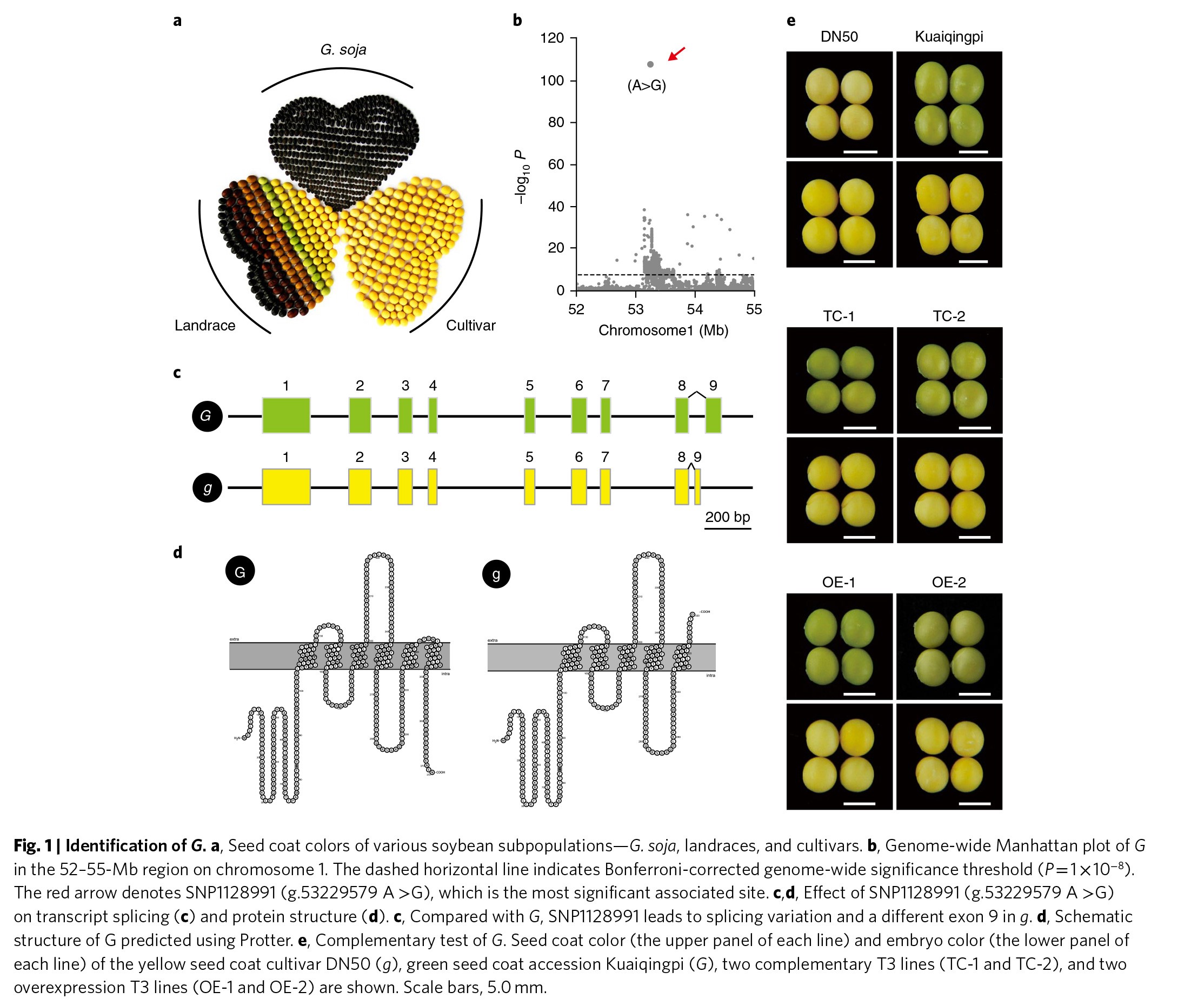
2. G基因是大豆中作用于种子休眠的驯化基因
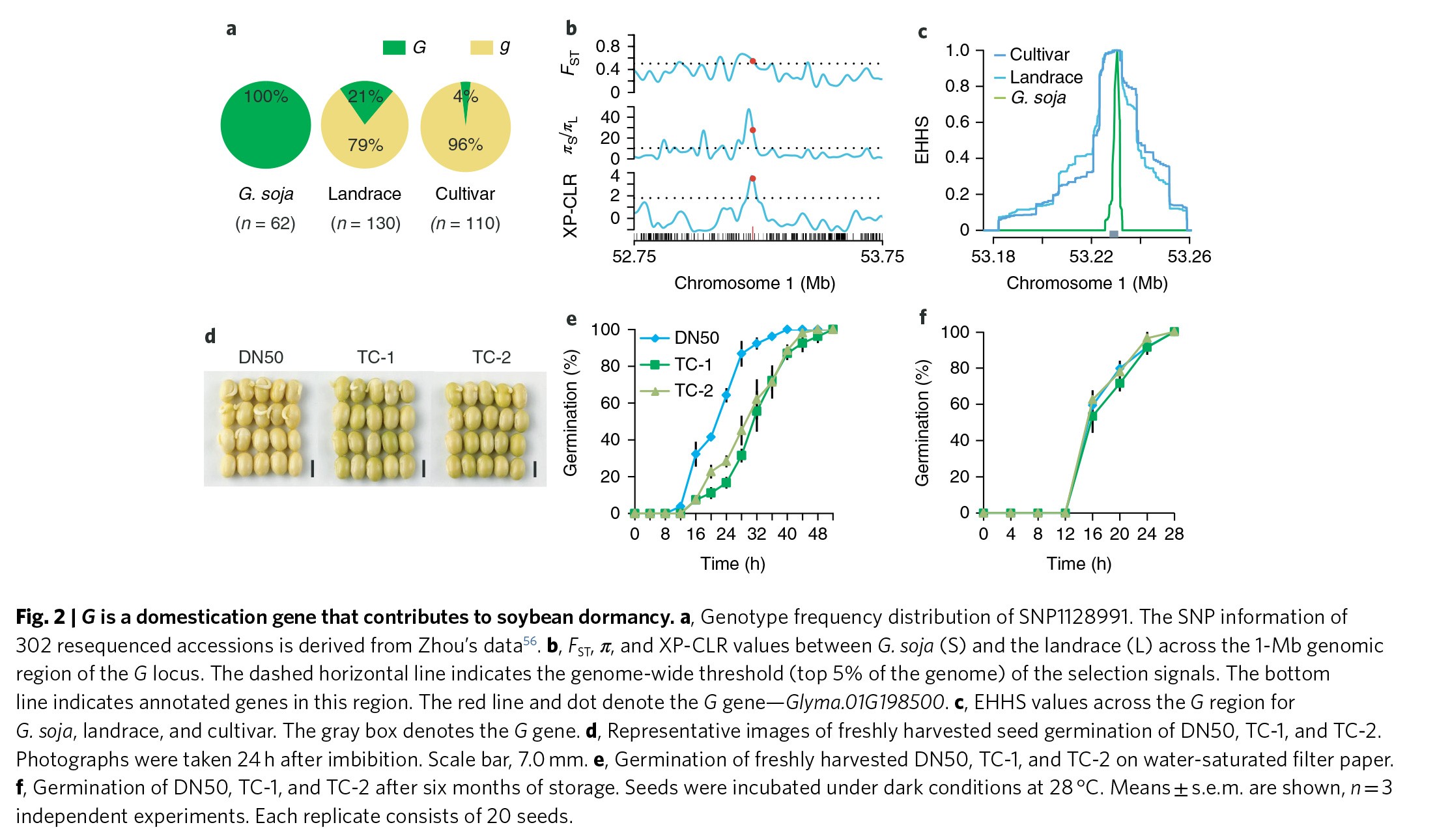
栽培种大豆是从野生大豆中驯化出来的,这一过程在中国5000多年前就已经开始。野生的大豆种子都表现为黑色,而各地的地方品种则颜色比较多样,其中表现最为优良的大豆品种的种皮则是黄色磨光种子(Figure 1a)。有趣的是,作者发现几乎所有的野生型大豆都带有G基因型(Figure 2a)。相反,在地方品种的大豆中G基因型占比降低到21%,而在栽培种大豆中则更是降低到4%(Figure 2a),说明在大豆驯化过程中G基因受到了选择。作者进一步通过群体间分化指数FST,核苷酸多样性π以及跨种群复合似然比XP-CLR来寻找G基因的序列在驯化过程中受到了选择的证据。这三个参数的结果均显示G基因所在的基因组区域处于选择的扫描区,即整个基因组序列排名前5%的受选择区域(Figure 2b & Figure S3)。此外,SNP位点的扩展单倍型纯合度分析(EHHS)显示野生型大豆相比于地方种和栽培种,存在更快的纯合衰减(Figure 2c),再一次证明了G基因可能是驯化相关的基因。
有研究显示种子黑色的种皮颜色将有助于保护种子在黑暗、模糊的环境下被捕食。因此,野生型大豆为何要保留绿色的种皮就是一个令人困惑的问题。而G基因侧翼区域在驯化过程中受到了选择,以及其功能与种皮的颜色相关,这让作者想到G基因可能还与大豆其它的一些重要性状相关联,比如说种子休眠。因此,作者比较了D50种子及转基因种子的萌发率,结果发现相比于D50的种子,转基因的种子萌发速率更慢(Figure 2d,e & Figure S2c)。当在室温中贮藏6个月之后打破休眠,转基因种子与栽培种大豆的种子表现出相似的种子萌发速率(Figure 2f & Figure S2d),说明G基因并未影响种子的萌发速率,而是影响了休眠。以上的试验结果显示G基因是大豆中作用于种子休眠的驯化基因。
3. 水稻和番茄中的G同源基因受到了平衡选择
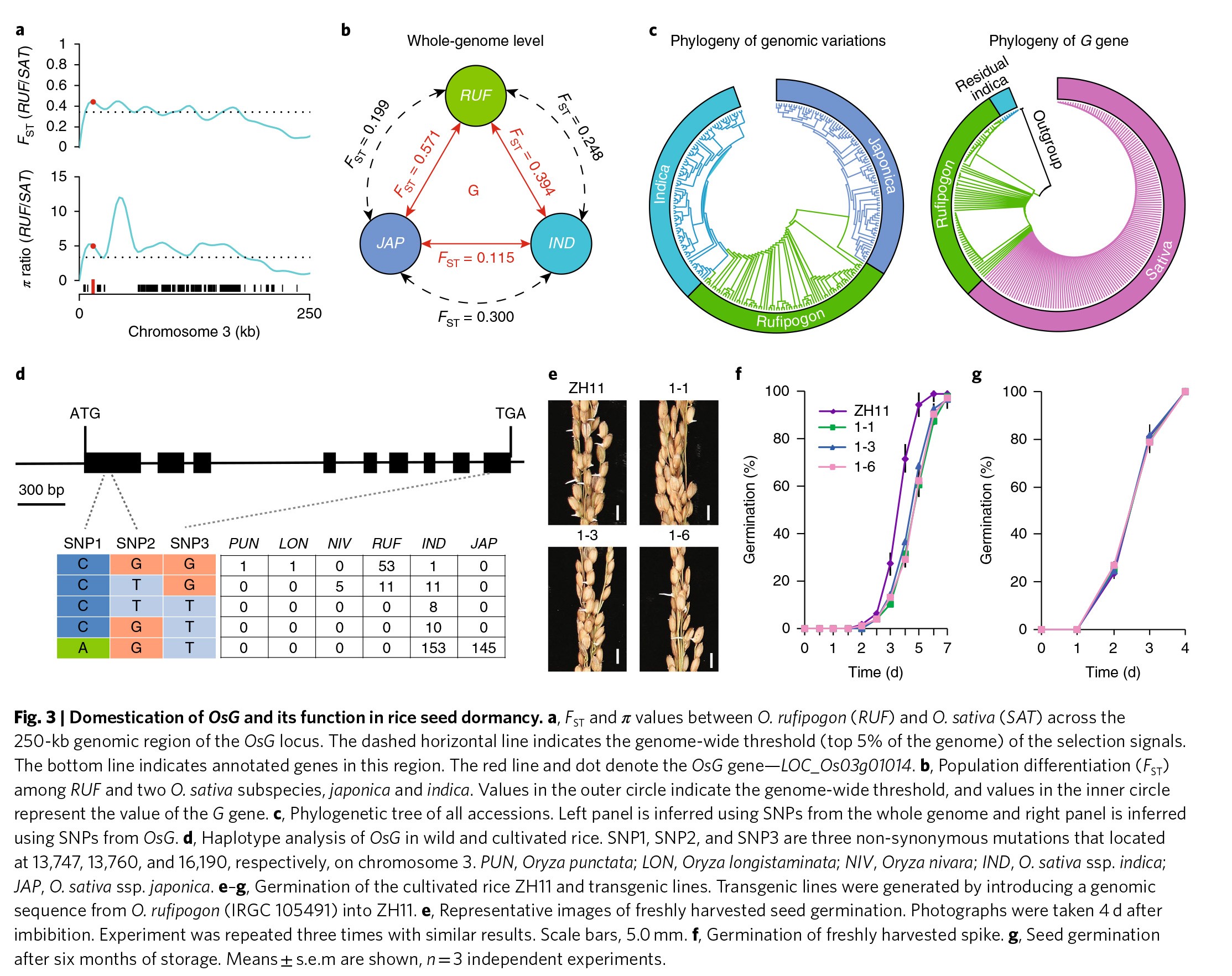
种子休眠的丢失,作为一个重要的驯化性状在不同科属作物间多次受到了独立的选择。因此,作者测试了在不同的作物中G基因的功能是否保守,以及是否收到了平衡选择。作者鉴定了不同物种中G基因的同源基因(Figure S4),并发现这些基因在C端高度保守(Figure S5)。水稻,属于禾本科的植物,是世界上最为重要的主食作物。亚洲栽培稻主要有两个栽培种:粳稻和籼稻,都是从其近缘种野生稻中驯化而来。为了检测大豆G基因在水稻中的同源基因LOC_Os03g01014是否也在驯化过程中受到的选择,作者对42个野生稻和134个栽培稻进行了平均深度为15×的重测序(Table S2)。随后,作者整合了这176个重测序数据以及之前已经报道的66个高深度、高质量的重测序结果,进行了全基因范围的选择扫描。FST,π以及EHHS的分析结果均显示水稻G基因也处于选择扫描区(Figure 3a & Figure S6, 7a)。
栽培稻的两个亚种,粳稻和籼稻形成了两个明显区分的类(Figure 3b, c),可能是由于相互独立的驯化或杂交过程所导致的。作者近一步对这些重测序数据以及之前报道的69份野生稻和328份栽培稻重测序数据(Table S3)进行水稻G基因的单倍型分析。作者共鉴定到了3个非同义SNP,可将水稻划分为5类单倍型(Figure 3d)。此外,位于水稻G基因C端上的SNP3将这些水稻划分为两大类:第一类包含了单倍型1和2;第二类包含了单倍型3、4、5(Figure 3d)。作者还发现所有的野生稻均含有第一类的基因型。除了少部分的籼稻,剩余的93.4%的籼稻和所有的粳稻都具有第二类的基因型(Figure 3d)。籼稻和粳稻中G基因的固定说明该基因在籼稻和粳稻还未分化时水稻早期驯化过程中就已经受到了选择,要么就是该等位基因能够克服粳稻和籼稻之间的部分生殖障碍,并且选择使得其在两个亚种中均保持在较高的频率。
为了研究水稻的G基因是否同样作用于种子休眠,作者将含有单倍型1的野生稻IRGC 105491中的基因LOC_Os03g01014序列转到含有单倍型5的栽培稻ZH11中。与正常的ZH11相比,转基因株系的种子萌发时间受到延迟(Figure 3e,f)。将该基因转化到另外一个含单倍型5的栽培稻HJ19中能获得类似的表型(Figure S8a)。然而,当贮藏6个月之后休眠被打破,转基因和正常植株的种子在萌发速率上没有差异(Figure 3g),再次说明了G基因影响的是休眠,而不是萌发速率。随后,作者利用CRISPR技术敲除了ZH11中的LOC_Os03g01014基因(Figure S8b)。相比于正常的ZH11,敲除株系的种子萌发速率更快(Figure S8d,e)。作者还在ZH11中过表达了两个野生稻的LOC_Os03g01014等位基因以及栽培稻ZH11本身的LOC_Os03g01014等位基因,结果显示转野生稻G基因的要比转栽培稻G基因的种子存在更强的休眠表型(Figure S8c-f)。此外,水稻G基因正好处于之前报道的与种子休眠相关的QTL区域,进一步证明了大豆G基因在水稻中的同源基因同样作用于种子休眠。
�
�番茄是茄科的一种重要的蔬菜作物。圣女果可能从红色果实的醋栗番茄驯化而来,并进一步发育成为果实较大的番茄。先前,有研究对包括番茄野生种、圣女果以及栽培种番茄在内的一共360个番茄物种进行了重测序。作者调查了重测序群体中大豆G基因在番茄中的同源基因Solyc08g005010的遗传多样性。FST,XP-CLR以及EHHS的分析结果均显示番茄G基因处于选择扫描区内(Figure S7b & 9)。进一步的研究在番茄G基因的第5个外显子上发现了一个非同义SNP(Figure S10a),该区域属于所有同源G基因十分保守的区域。大多数的醋栗番茄具有G等位基因,而大多数的圣女果和番茄具有C等位基因(Figure S10b),说明了番茄的G基因同样在驯化过程中受到了选择。
4. 拟南芥中的G同源基因具有保守的功能
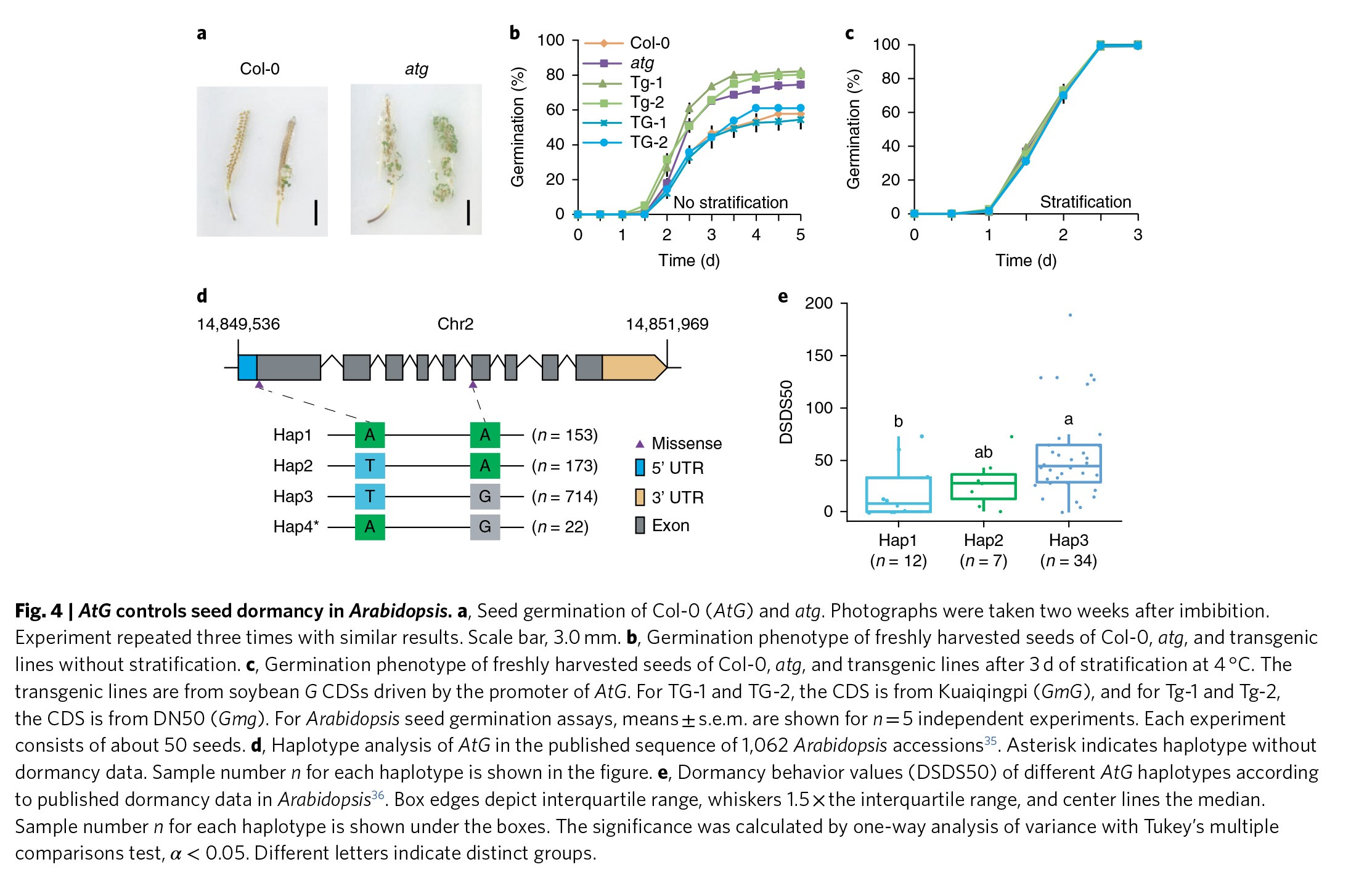
大豆G基因在拟南芥中的同源基因是At2G35260。作者调查了At2G35260基因在种子休眠中的作用。相比于野生型拟南芥,两个拟南芥G基因突变体具有显著的更快的萌发速率(Figure 4a,b & Figure S11)。作者通过层积的方法来打破拟南芥种子的休眠,结果显示野生型和G基因突变体层积之后具有类似的萌发模式(Figure 4c & Figure S11),说明拟南芥的G基因同样作用于种子的休眠。然而,拟南芥的G基因对于种子休眠的影响要小于另外一个影响种子休眠的基因DOG1(Figure 4b,c)。
作者通过拟南芥G基因的启动子分别带着大豆Kuaiqingpi的G等位基因和DN50的g等位基因,将其分别转入到拟南芥G基因突变体中。结果发现大豆Kuaiqingpi的G等位基因能够完全互补拟南芥G基因突变体的表型,而DN50的g等位基因则不能(Figure 4b,c)。这些结果说明G基因的同源基因具有十分保守的控制种子休眠的功能,同时也说明了大豆G和g等位基因确实与种子休眠的驯化有关。
作者还对已经报道的超过1000份拟南芥种质进行了G基因的多态性分析,结果显示存在4中单倍型(Figure 4d)。有趣的是,通过很多种质的驯化数据,作者发现该基因不同的单倍型具有差异显著的种子休眠水平(Figure 4e)。
5. G基因通过ABA途径影响种子休眠
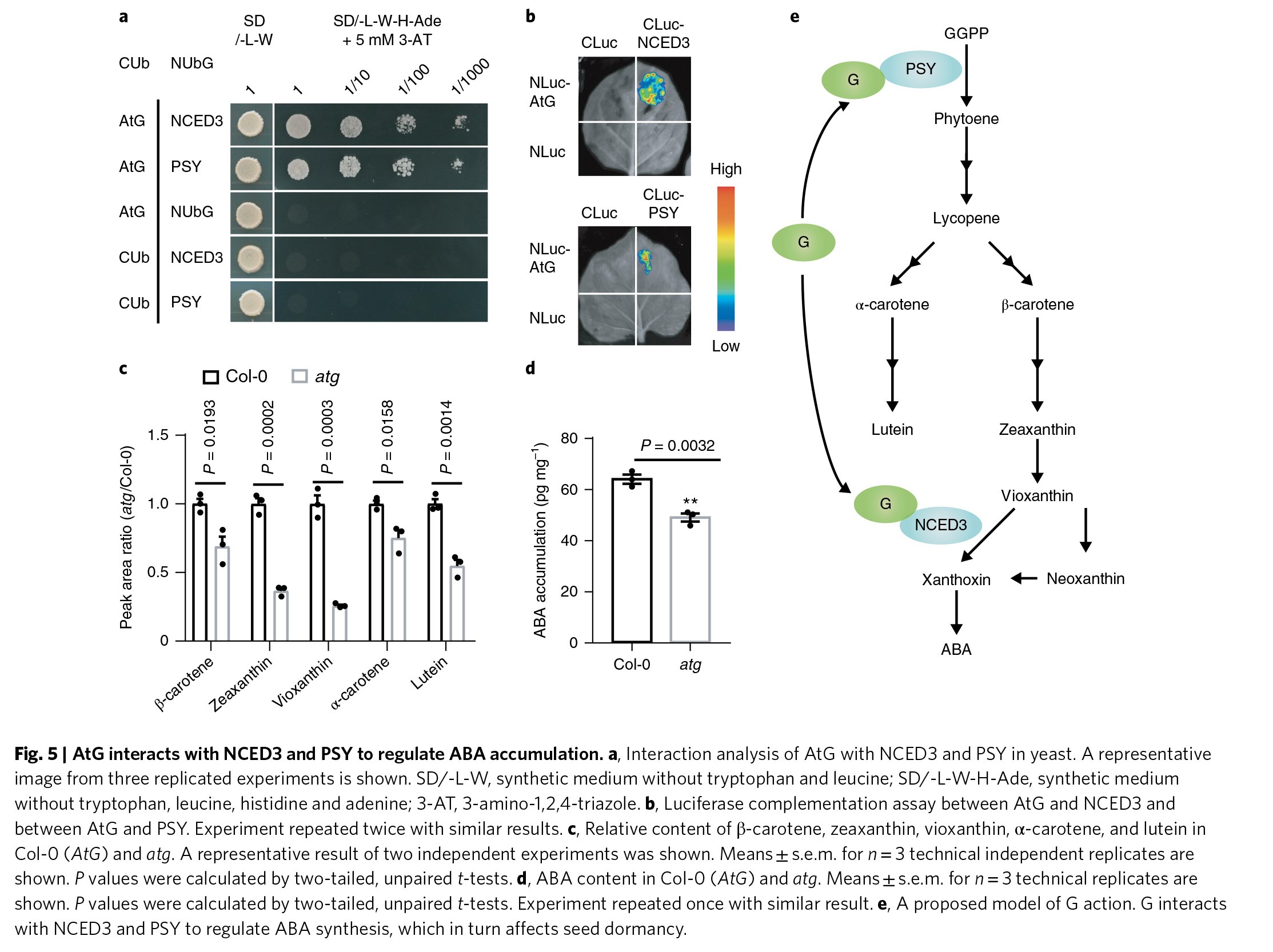
为了研究G基因影响种子休眠的分子机制,作者进行了酵母双杂的试验。作者发现拟南芥的G基因与9-顺环氧类胡萝卜素双加氧酶NCED3和八氢番茄红素合成酶PSY相互作用(Figure 5a),这两个酶是参与ABA生物合成的关键酶。作者进一步通过荧光素酶互补试验、免疫共沉淀及双分子荧光互补试验验证了它们之间的互作(Figure 5b & Figure S12a,b)。作者还检测了ABA生物合成前体的量,结果显示类胡萝卜素通路中的主要成份,包括β-胡萝卜素、玉米黄素、紫草黄素、α-胡萝卜素及叶黄素的含量在野生型中均要比拟南芥G基因突变体中要高(Figure 5c)。同时,拟南芥野生型种子中的ABA含量要比G基因突变体中的的要显著的高(Figure 5d)。
另外,大豆中的G基因序列和过表达转基因株系相比于DN50,ABA含量显著的高(Figure S12c)。这些结果说明G基因通过调节ABA的生物合成影响种子的休眠。因此,作者提出G基因通过与类胡萝卜素和ABA生物合成途径中的NCED3和PSY互作,进而影响种子的休眠(Figure 5e)。在g突变体株系中,ABA生物合成减少,因此种子休眠减弱,进而有利于农民对于作物的管理,最终导致了不同作物中对于g基因型的平衡选择。
Conclusion
驯化可被认为是最早的育种形式。作物驯化的时间预估在4000多年前就已经开始。现在,玉米、小麦、水稻、大豆和向日葵等驯化的作物已经成为了人们主要的日常食用作物。另外,全球的食物供给正在变得越来越同质化。作物品种多样性低对于食物安全是一个潜在的威胁。育出环境上可持续型、低投入、且能适应极端气候环境的作物被认为是第二次绿色革命。对现有作物的改良可能并不能满足上面提出的需求,这些已有的作物品种并不满足低投入、适应极端环境的需求。因此,驯化新的作物品种可能对于未来的食物安全来说非常重要。
鉴定参与驯化的基因,尤其是那些在不同作物物种间受到平衡选择的基因,对于我们进一步理解驯化的进程以及加速新作物物种的驯化具有非常重要的意义。作物驯化可以分为两步走,其中驯化综合征的驯化会被优先考虑。迄今为止,研究者们已经鉴定了很多的驯化基因,但极少有基因受到平衡选择;比如说,禾本科内不同物种间,作用于粘性的Waxy基因、作用于落粒性的sh1基因以及作用于开花时间的HD1基因。但具作者所了解的情况,还未见对不同科的植物之间的驯化基因受到选择的报道。因此,对于亲缘关系较远的作物之间是否存在驯化基因受到平衡选择是一个悬而未决的问题。寻找驯化综合征潜在的驯化基因吸引了许多生物学家的长期关注。
本文中,作者揭示了G基因的同源基因至少在豆科、禾本科及茄科3个作物谱系中受到了平衡选择,这可能是第一个独立于作物科属而受到平衡选择的驯化基因。鉴于拟南芥中的G基因同样具有相似的功能,所以G基因可能同样在十字花科作物中作用于种子休眠。一旦有更多的重测序数据公布,对于G基因是否在其它作物物种中存在平衡选择的问题非常值得研究。
种子休眠是非常复杂的性状,由许多的QTL所控制,水稻中至少有30个QTL与种子休眠相关,小麦中则至少有20个QTL,生菜中则有一些QTL。尽管有30多个QTL作用于水稻的种子休眠,但没有一个主要的QTL能够解释休眠和非休眠亲本间表型变异的25%。通常来说,每个QTL大约能够解释表型变异的5%~15%。两个独立的研究显示水稻G基因分别位于能够解释表型变异6.13%和11.3%的QTL位点上,说明G基因是众多微效基因中的一个。这个结果与本文作者的研究结果一致。事实上,G基因的选择并不是功能丢失等位基因,而是功能弱化等位基因。尽管功能丢失等位基因可能存在更加强烈的表型(Figure S8),它将导致叶片变黄的表型(Figure S13)。在大豆中,种子休眠更加复杂,涉及到生理和物理结构两个方面。之前的研究鉴定到了一个与种皮物理结构相关的基因Hs1-1。本文所鉴定的G基因作用机制不同于Hs1-1基因,其代表了生理层面的种子休眠机制。是否存在更多的影响种子休眠的基因在不同作物物种间受到平衡选择仍然需要未来更多的研究。更多的驯化基因的鉴定将促进人们对于驯化综合征的进一步理解,同时也能加速新作物物种的驯化进程。
通讯:田志喜(http://sourcedb.genetics.cas.cn/zw/zjrck/201105/t20110503_3124592.html)

�
�个人简介:1997年,河北农业大学,学士;2000年,河北农业大学,硕士;2007年,中国科学院遗传所,博士;2007-2010年,美国普渡大学,博士后。
研究方向:利用关联分析、图位克隆和转基因手段,并结合比较基因组、生物信息学分析方法对影响大豆产量和品质的网络调控系统进行系统分析,揭示调控大豆器官发生、种子发育、植株形态建成以及品质形成的内在机制,为大豆分子育种奠定基础。
doi: https://doi.org/10.1038/s41588-018-0229-2
Journal: Nature Genetics
Published date: 24 September, 2018
(P.S. 原文下载:链接:https://pan.baidu.com/s/1qFiSsqwlF_7JjmGMbedfhQ 密码:dmu6)

https://blog.sciencenet.cn/blog-3158122-1137924.html
上一篇:Nature:气候变暖对于冻原生物群落植物功能性状的改变
下一篇:Plant Cell:植物解毒反应作用于光氧化胁迫下的耐受性
全部作者的其他最新博文
- • Plant Physiology:CsMADS3促进柑果中的叶绿素降解和类胡萝卜素合成(华中农业大学)
- • Molecular Plant:LBD11-ROS反馈调节作用于拟南芥的维管形成层增殖和次生生长(浦项科技大学)
- • Science Advances:根结线虫通过调控植物的CLE3-CLV1模块,促进侵染进程(日本熊本大学)
- • Nature Communications:油菜素内酯参与植物营养生长期转变的分子机制解析(浙江农林大学)
- • Current Biology:光合作用产生的蔗糖驱动侧根“生物钟”(德国弗莱堡大学)
- • PNAS:花同源异型基因在叶中被抑制、花中被激活的分子机制(南卡罗来纳大学)