博文
人类会亘古长存吗?
 精选
精选
||
相信很多人都思索过我们人类自身的命运,它会亘古长存吗?然而,地球生命的演化历史却告诉我们,“创造、进化与毁灭”才是地球系统之永恒的生命旋律!如果你问:是否有诞生以后永不灭绝的物种? 答案是否定的。据估计在地球的生命历史中,超过90以上(可能接近98%)的物种都已经灭绝了(Gastonand Spicer 2004)。
一、生命的宏观演化—地球上物种的吐故纳新
1. 物种的自然寿命—一般100万~2000万年
既然物种总会绝灭,那到底一个物种可以持续多久?这似乎也不是一个容易回答的问题。这首先需要解决如何确定一个物种在自然界中的寿命,很显然,迄今为止最好的一个办法就是根据化石记录。一般来说,可将某个物种的化石在地质历史时期的跨度作为该物种的自然寿命。譬如,某种螺类最早的化石出现在距今1310万年前,消失于距今1230万年前,因此估算其寿命约为80万年(图1)。当然,有学者认为这种依赖于化石的估计可能偏向于寿命长的物种,以此来推算所有物种的平均寿命可能导致平均寿命的高估(Belk and Borden 2004)。当然,根据化石推算的物种寿命可能被该物种更早或更晚的化石的出现而改写,此外,也不一定所有的物种都会保留清晰完整的化石。
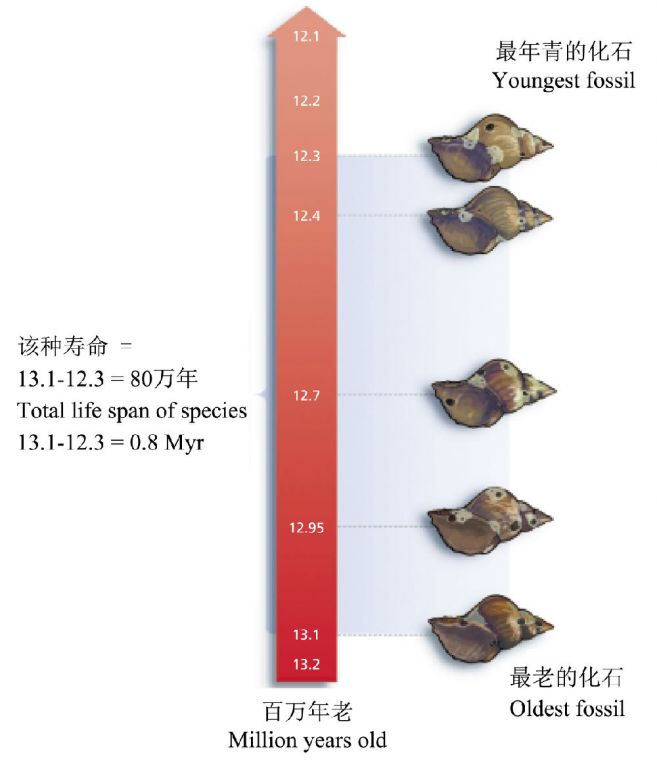
图1 物种寿命的估算。这些化石贝壳的年龄是通过包埋它们的岩石的放射性测年来确定的(引自Belk and Borden 2004)。
基于来自各种类群(海洋和陆地)的化石证据,估计一个物种的平均寿命大约为500万-1000万年(May et al. 1995)。也有认为这个数字约为100万年(Belk and Borden 2004)。而从表1可以看出,不同类群的物种的寿命差异很大,海洋生物的寿命(670万-2500万年)比非海洋生物的寿命(100-400万年)要长得多,这可能反应了海洋环境相对稳定有利于物种的生存,而陆生环境变化相对剧烈,物种容易绝灭。
表1 估算的化石物种的平均寿命
类群 | 寿命(百万年) |
海洋 Marine |
|
珊瑚Reef corals | 25 |
双壳类Bivalves | 23 |
底栖有孔虫类Benthic foraminiferans | 21 |
苔藓虫类Bryozoa | 12 |
腹足动物Gastropods | 10 |
浮游有孔虫类Planktonic foraminiferans | 10 |
海胆类Echinoids | 7 |
海百合类Crinoids | 6.7 |
|
|
非海洋Non-marine |
|
单子叶植物Monocotyledonous plants | 4 |
马Horses | 4 |
双子叶植物Dicotyledonous plants | 3 |
淡水鱼类Freshwater fish | 3 |
鸟类Birds | 2.5 |
哺乳动物Mammals | 1.7 |
昆虫Insects | 1.5 |
灵长类Primates | 1 |
(引自McKinney 1997)
Raup(1994)统计了17500个海洋化石动物属的寿命,发现大多数属的寿命在2000万年以内,而少数属的寿命很长(图2),但与地球生命的历史相比,没有属能幸存很长的时期,最长的虽然可持续1.6亿年,但也只有生命历史的约5%(Gaston andSpicer 2004)。
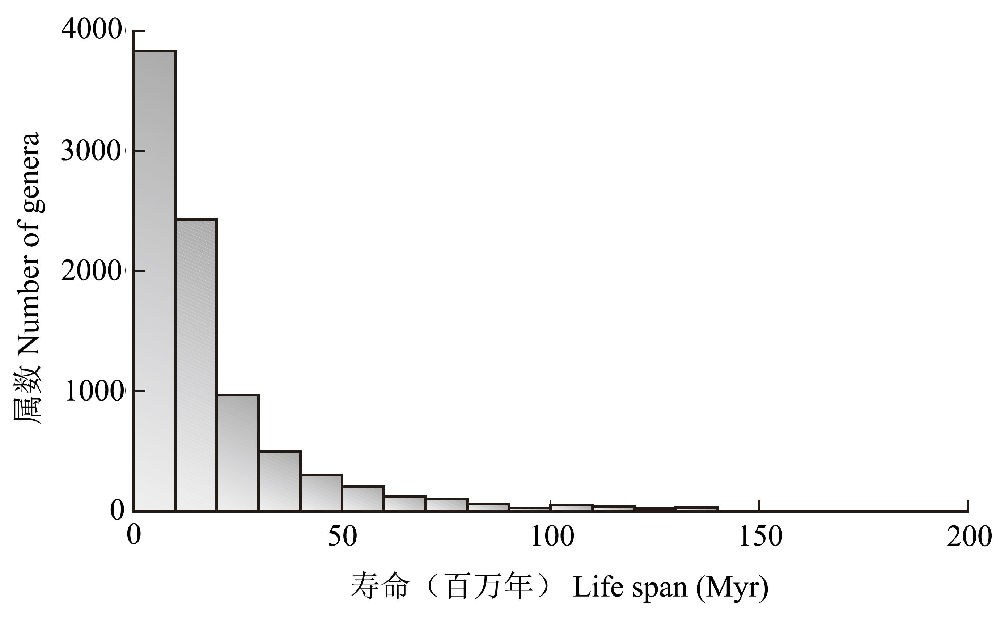
图2已经灭绝了的17,500个海洋动物(脊椎动物、无脊椎动物和微体化石)属的寿命(仿Raup 1994)
2. 成种的时间—一般10万~500万年
迈尔(2008)认为,控制成种事件速度的主要是生态因素,而不是所谓的“突变压力”,当物种被地理或生态屏障隔离时,而且该物种很少有基因流动时,成种事件就会非常迅速和频繁,而在通畅的大陆成种事件则很少。一般估计,从一个祖先开始,大概要用10万~500万年的时间才能演化出一个彼此生殖隔离的后裔物种(科因2009)。
但是,迈尔(2008)认为,在化石基础上对成种事件平均速度的计算可能会导致错误的估计,因为化石记录的主要是分布广泛的群体,这些群体通常有很长的生命周期,因此成种事件的速度很慢。
马拉维湖中的丽鱼是一个快速的物种分化(同域分化)的案例,仅在过去70万年间,就从单一的丽鱼祖先衍生出400多个新物种(Danley & Kocher 2001)。但是,也有相反的例子:北美东部存在的一些植物(包括臭菘),也存在于东亚的一些地区,两个大陆的群体不仅在形态上没有什么区别,虽然分离了600~800万年,但还可以相互交配(迈尔2008)。
虽然化石证据存在不足,但如果都用化石进行比较,还是会有一定的参考价值,即物种的化石寿命要显著高于化石成种时间。这也符合地球上的物种多样性正在日益增加的宏观演变趋势。
3. 维管束植物的演化与更替—从蕨类到裸子植物再到被子植物
高等植物依据结构、繁殖特性等的不同,分为苔藓植物、蕨类植物(又称羊齿植物)、裸子植物和被子植物。蕨类植物、裸子植物和被子植物又称为维管束植物。绿色植物在水陆之间的界限分明:低等植物统治水体,高等植物统治陆地。
植物的演进还留下了不可磨灭的生态历史足迹,漫长的结构和功能演化叠加在明晰的生态适应目标之中。高等植物的演化与更替反应了其从适应水生逐渐向适应陆生的转变过程。苔藓类的维管系统发育很不完善,因此它们局限于在潮湿的环境中生存。维管束植物体内有输送水分和养分并具有支撑作用的维管系统的存在。蕨类植物是早期登陆的原始的维管束植物,由于其输导系统的效率不高,一般也难以在干旱环境中生存。裸子植物形成花粉管和种子,花粉管的出现使受精作用摆脱了对水的依赖,这对适应陆生环境具有重要意义,而被子植物是现代植物界中最高级和最繁荣的一个类群,因其具有真正的花,故又称为显花植物。被子植物对陆地生活方式的适应最为完善,它们甚至能在极干旱的荒漠地区生存。
此外,为了适应陆地生活,陆生植物的外表还演化出角质层,以减少水分的蒸发,仅通过较小的气孔吸收光合作用的必需底物—CO2,并通过根系和导管弥补蒸腾引起的水分损失。
在古生代(6.0—2.3亿年前),植物由水生登陆后演化出蕨类,裸子植物步入缓慢的发展;中生代(约2.3—0.65亿年前)蕨类植物显著衰退,裸子植物迅速扩展,取代了蕨类植物的优势地位;在新生代(6500万年至今),被子植物迅速扩展,裸子植物逐渐衰退,蕨类植物的分布范围也进一步缩小,并多限于温暖地区,世界上第一次出现了百花争艳的景象,被子植物极度繁盛,植物界迎来了它的新霸主(图3)。

图3 泥盆纪以来维管束植物的多样性,虽然不同的时期不同类型的植物占优势,总的多样性增加了(Jeffries 2006重绘自Groombridge1992)
4. 动物群的演化与更替—大爆发与大灭绝
动物也有高等和低等之分,前者指脊椎动物,后者指无脊椎动物,这就类似于高低等植物以维管束的有无为重要特征一样,脊椎和维管都是重要的支撑系统,是动植物得以大型化的结构基础。与植物明显不同的是,低等动物和高等动物无论在水体还是在陆地,都是生态系统重要的结构成分。
与植物类似,动物在结构上的演化也经历了从小型到大型、从简单到复杂的历程,只是动物的演化历史似乎更为曲折与精彩。在古生代早期(寒武纪和奥陶纪),无脊椎动物在海洋中一统天下,随着“掠食者”的出现,生物界开始进入弱肉强食的残酷时代。在古生代中期(志留纪和泥盆纪),植物和无脊椎动物(昆虫)登上陆地,鱼类统治了海洋,在泥盆纪晚期,从鱼类进化出两栖类,脊椎动物开始登上陆地。在古生代末期(石炭纪和两叠纪),栖居水边沼泽中的两栖类称霸地球,并继续向爬行类演进。在中生代,爬行类的恐龙横空出世,动物界迎来了新霸主。进入新生代,恐龙日益衰落,并几乎灭绝,同时,披上毛发的恒温动物—鸟类和哺乳类开始繁荣,成为了动物世界的新霸主。
与植物学家略有不同,动物学家将历史上统治地球的动物类群大致按地质年代的先后可分为三大类—寒武纪动物群(Cambrian fauna)、古生代动物群(Paleozoic fauna)和现代动物群(Modern fauna)。按照这种分类系统所看到的动物类群演化与更替的一个最大的特点就是大爆发与大灭绝。
在距今约5.3亿年前(寒武纪),绝大多数无脊椎动物门(节肢动物、软体动物、腕足动物和环节动物等)在几百万年的短时间内出现了,而没有在寒武纪之前更为古老的地层中出现,被称作“寒武纪生命大爆发”。大约0.6亿年以后,出现了古生代动物群。但是,3亿年后,出现了二叠纪大灭绝(GreatPermian extinctions),接着又出现了三叠纪生物大爆发(Triassic explosion),由此演化出现代动物群(Purves et al. 2007,图4)。
为何在寒武纪早期发生持续时间很短的结构创新大爆发呢?迈尔(2010)认为,之所以发现了很多新的化石,是因为这些化石动物都有骨骼,而它们的那些软体祖先可能无法石化,而且可能个头太小,无法在地层中留下痕迹,因此,这么多门类表面上的突然同时出现或许只是个假象而已。除此之外,后生动物早期进化速度太快也可能是一个原因:当一个身体构型内的整合还足够疏松时,往往允许大的创新发生。当然,这都只是猜测而已,难以证实。
有证据表明,地球曾经历过5次大灭绝事件,其中50-90%的物种在数千年到数十万年期间灭绝了(Belk and Borden 2004,图4),其中,二叠纪大灭绝(距今约2.5亿年)为有史以来最严重的大灭绝事件,堪称地球历史从古生代向中生代转折的里程碑。据估计,在这次大灭绝中,地球上有96%的物种灭绝,其中90%的海洋生物和70%的陆地脊椎动物灭绝。三叶虫、海蝎以及重要珊瑚类群全部消失,陆栖的单弓类群动物和许多爬行类群也灭绝了,生态系统获得了彻底更新。
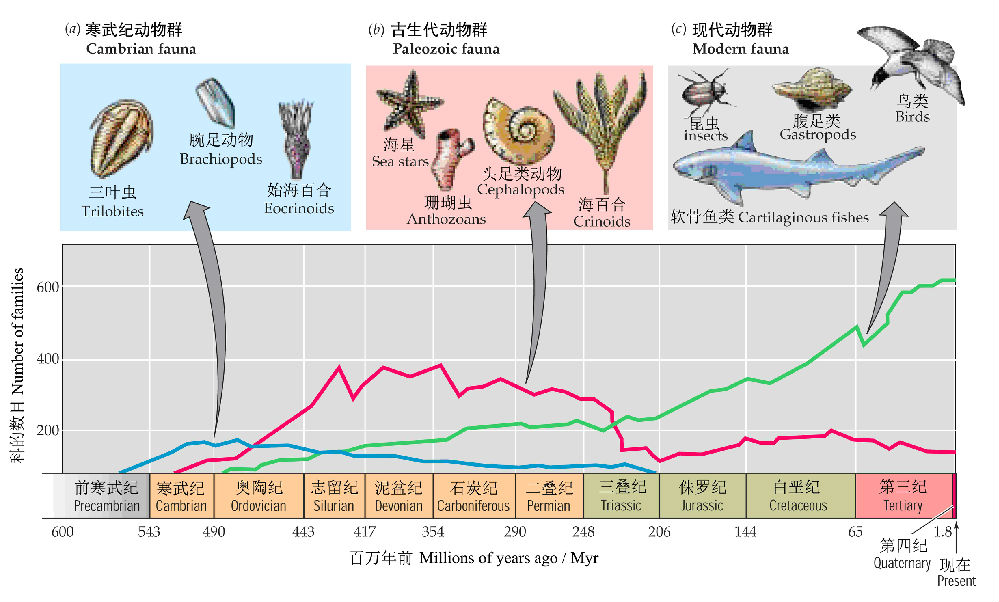
图4 三个大的进化动物群的代表以及地质年代中每个动物群的科的数目(引自Purves et al. 2007)
据称,每一次大灭绝后,物种多样性经过500万年—1000万年都不能恢复到灭绝前的水平,而且后来的物种也与灭绝前的物种非常的不同(Belk and Borden 2004)。历史上的生物大灭绝可能源于巨大的地球环境灾变—譬如由于气候波动导致的海平面变化或者起由于地球构造板块运动引起的海洋和地形的变化,等等。许多科学家担忧我们可能正在见证第六次生物大灭绝,而引起这个灭绝的无处不在的全球变化却是人类活动(Belk and Borden 2004)。
二、创造与毁灭—轨迹不同的轮回
1. 地球环境—永恒变化但不可预测
我们不能忘记,如今我们生活的这个绚丽多彩的生命世界其实是地球环境在漫长的地质岁月中艰辛演化的产物。因此,生命的演进模式必定是适应地球的结构与环境演化轨迹的结果。那地球环境是如何演化的呢?
十八世纪,苏格兰地质学家赫顿(Hutton J, 1726-1797)提出地球历史没有方向,只是不断重复自我创造与毁灭的轮回,无始无终。十九世纪,一个名叫赖尔(Lyell C, 1797~1875)的英国律师在其著作“Principles of Geology”中重新提出了赫顿50年前地球不断规律变化的学说,他不同意赫顿提出地球整体经历创造及毁灭循环的说法,认为地球不断在进行局部改变,这里侵蚀,那里爆发,并无一定方向,且历时久远,不可想象。
毋容置疑,地球的结构及其环境处于永不停息的变化之中,但却充满着偶然与随机,几乎不可预测,当然,这并不否认人们对一些很短期的自然过程的可预测性。我试图询问一下这样的问题:谁能说清楚寒武纪以来的陆地板块为什么以这种方式崩析、组合和漂移?谁又能说清楚未来的5亿年陆地板块将会以怎样的模式运动和变化?世界除了永不停息的变化外,还可能有其它事物能够永恒存在吗?
2. 物种更替—休克式的毁灭
地球上的生命以何种形式进行毁灭与更替?是均衡渐变式还是集中突发式?大量的地质历史证据表明,物种的更替采取了一种剧烈的节律性的脉动模式:自古生代的奥陶纪以来,地球曾经历过5次大灭绝事件:1)距今4.4亿年前的奥陶纪末期,约85%的物种灭绝;2)距今3.65亿年前的泥盆纪后期,海洋生物遭遇灭顶之灾;3)距今2.5亿年前的二叠纪末期,超过95%的地球生物灭绝;4)距今2亿年前的三叠纪晚期,爬行类动物遭受重创;5)距今6500万年前后白垩纪晚期,侏罗纪以来称霸地球的恐龙几乎绝迹。
生物大灭绝的一个特点就是突发性—在地球经历过的5次大灭绝事件中,50-90%的物种在数千年到数十万年的“短”时期内灭绝了(Belk and Borden 2004)。这简直就是一种休克式的灭绝!这可能折射出这样一个事实,即地球环境的变化并不温和—长期缓慢的变化夹杂着若干次重大灾(巨)变!
大多数海洋动物属的寿命在2000万年以内(Raup 1994),而陆生动植物的寿命仅有100-400万年(McKinney 1997)。显然动物的平均寿命特别是陆生动植物的寿命远短于大灭绝的间隔期,这似乎意味着,绝大多数动植物的适应与进化难以应对周期性的环境灾(巨)变,这反过来也许是地球上的生命反复出现休克式毁灭的原因之一。
比较3和图4不难看出,动物界对灾变更为敏感,反应更为剧烈,物种的更替也似乎更为彻底。而高等植物则表现出了更强的整体稳定性与可塑性,似乎沿着一条明确的进化路线(蕨类→裸子植物→被子植物)“毫不动摇”地演进。从这点来看,与动物界相比,植物界似乎对地球环境的周期性波动具有更好的适应性或耐受性,整体上呈现出一种宏观性K-选择者的特质,也显示出它们的生活史策略在进化上的成功;而动物界则是大起大落,整体上像个不太成功的机会主义的r-选择者!其实这不无道理,如果将较大型的高等动植物进行比较,植物体型更大,寿命更长,种子的休眠(而哺乳动物的胎生几乎使其休眠能力丧失殆尽)与抗逆性更强……这些都使得植物在进化策略上更像K-选择者,而地球环境的巨变好像也在它们整体能够适应的范围之内。其实,被子植物的类型也十分多样,从草本到灌木到高大的木本,具备各式各样的生态对策。
3. 适应悖論—复杂的毁灭
1)生命表型“进”“退”自如—可能因为基因喜欢插入和缺失
从本质来看,生命为了应对不断变化的环境而进行的对自我(结构与功能等)进行不断修正和适应是通过基因的变化来实现。有意思的是,在原核生物经历了极其漫长的地质岁月艰苦地完成了支撑所有生命基础的基因原始创新之后,真核生物则选择了一种快速而便捷的方式来实现基因的改变,即主要是通过积木式的拆拼来实现功能的删除或添加或叠加,而这主要是通过有性生殖(或许关键在于减数分裂同源染色体的联会与频繁的遗传重组)的平台来实现的。
为何生命能“进”“退”自如,即既能复杂化也能简化?这可能源自有性生殖中细胞减数分裂的特性—喜欢玩耍基因的插入和缺失,由于表型来自基因,因此基因的插入和缺失就意味着表型的添加和丧失。这种改造方式不仅便捷快速,而且相对安全。
胚胎的相似、重演和痕迹结构在某种程度上来看,是一种个体发育重演了系统发生的过程,也是这种生命拼接式叠加与传承模式的缩影或写照。一种解释是,因为受遗传控制的发育程序无法省去这些含有祖先生理结构的发育阶段,而且在后面的发育阶段中不得不改变这些结构,以便使它们适应新的生命形式,即胚胎中祖先器官的出现成了确保重构器官发育的内在程序(麦尔2010)。按照弗朗索瓦∙雅各布的话说,大自然是出色的修补匠,而不是高超的发明者(古尔德2008)。
2)温和的完美—复杂的毁灭,简单的永生
在较为温和(相对于灾变来说)的生存环境中,有性生殖堪称“完美”,创造出了各式各样的无与伦比的精美的生命形式,好像无所不能,但为什么(特别在动物界)还会一次又一次地遭遇地质历史时期生物物种的休克式大灭绝?为什么生命经历如此惨烈的灭绝之后还留下了可以燎原的火种?
依我之愚见,在允许复杂化的生命舞台上(譬如掠食者与猎物系统),表型的适应就像一台欲望无尽的游戏,或许出于一种生命自私的本能,无论是在无情且无奈的达尔文式自然选择压力下,或是拉马克式“完美”适应欲望下,一些强者(如猎食者)欲变为更强者,对万物的控制欲望催生更加或过于复杂的遗传调控机制,其结果可能是这些“王者”获得了对温和的环境变化具有强适应力的同时,却失去了对灾变环境(往往是非生物)的抵御能力,因此它们往往难逃灭绝的噩运。
从遗传本质上来看,有性生殖通过基因不断堆积来创造变异与适应(最终新物种)的方式,不可避免地会导致基因组、表型、结构与调控的过度复杂化以及体型的过度大型化,这就埋下了致命的隐患。基因及其调控还有生命整体绝不可能趋于无限复杂,终究会将发展途径自我阻断,使某个方向的进化嘎然而止。
结构复杂的动物个体(可视为中尺度上的K—对策者)对中度以下的环境波动具有完美的缓冲性,但是,这已牺牲了它们的繁殖速率,也牺牲了它们更广泛的变异能力与适应范围。因此,一旦遭遇到环境灾(巨)变,复杂的生命往往难以进行大幅度的改变与适应,不可避免地遭受灭顶之灾!庞然大物恐龙的灭绝就是一个活生生的悲惨案例。我推测,生命特有的这种不断复杂化的遗传进化方式可能是导致这种灭绝的内在机制之一。这种现象常常被直生论者称为“内部完美原则”或内在的“种系动力”,但我不完全同意这种神话般的本性或种系动力的说法,我认为,最初是为了适应,但是由于这种适应的创造方式使机体越来越复杂,逐步降低了机体的可塑性(这与生态系统的复杂性与可塑性的关系完全不同),最终在环境巨变来临时惨遭灭绝!
3)古老的简单—永恒的生存
早期动植物体积微小,结构极为简单,分化的细胞类型也极为有限,遗传调控机制也应该相对简单。一些简单而低等的生命(如细菌、蓝藻等),虽然甘为他人做嫁衣,角色也不华丽,但是凭借着不起眼的变异能力加上超强的繁殖力(r-对策),却能长期占据属于它们的那一份舞台,而近乎永恒地留在了地球生命的历史舞台上!这似乎是一种“以不变应万变”,或以小变(或快变)应大变的生存机制!
或许正是这些较低等生物顽强的生命力使它们在地球生命经历大的灾难后的恢复与重建过程中起到了至关重要的作用,因为它们默默无闻的先锋与开拓作用以及卓越的创造复杂的能力,才可能为更高级生命(特别是一些却后余生者)的重新登场与进化奠定了基础。灾难之后,这些卑微者将重新制造或迎接一批批新的统治者的喧嚣登场,目睹它们之间残酷的争斗,直到下一次大灾难来临,世界又彻底恢复寂静,它们又开始收拾残局,艰难地铺垫新的进化之旅,送迎一批又一批热热闹闹登台表演的“明星”……如此循环往复。
谁会是那些却后余生者呢?美国古生物学家柯普曾提出了一个很有意思的“非特化法则”:1)地质时代中高度发展、特化的类群,并未孕育出下一代的类型,相反,后代主要从前代的非特化祖先那里获得进化的原动力,这些非特化的种类往往对栖息地和气候具有更广泛的适应性;2)而各个时期特化的种类通常无法适应新时代的变化,这些变化对于大体型通常造成严重的后果,因为它们需要大量的食物,食物缺乏时,小型种类可以生存,大型的就遭到消灭;3)杂食性的动物可以继续生存,但需要特殊食物的种类就会死亡;4)非特化的种类,体型都不大,哺乳纲源于小体型的祖先,其它脊椎动物也一样(古尔德2009)。
4. 生命的演进—不喜欢简单重复
物理世界的内在驱动力就是以惯性(牛顿)质量为基础的吸引力与排斥力的对立统一,外部表象即相对的运动与静止,总体演化方向是从有序趋于无序。而生机勃勃、充满着奇异的有机世界除了蕴含无机化学的吸引与排斥、运动与静止外,其内在的核心驱动力就是遗传与变异的对立与统一,并呈现出多重相互对立的外部表象—追捕与逃亡(行为)、进化与退化(器官)、出生与死亡(个体)、诞生与灭绝(物种)……等等。但是,生命系统演化的总体方向是从无序趋于有序、从简单趋于复杂或多样。
自然界是由生命—化学—物理系统相融与镶嵌而形成的统一体,换言之,生命系统是与地球环境长期协同演化(进化)而来的产物,这是无可争议的事实之一。另一个不可争议的事实是进化在整体上并未以一种稳定可预测的方式运行与发展。试问,一个(整体)不可预测的地球环境怎能创造出一个(整体)可预测的生命系统?
不同于非生命的封闭系统(那里变化趋于复杂性减少、失去秩序、衰变退化∙∙∙∙∙∙),开放的生命系统,在太阳辐射和水的联合驱动下,不断地重复着从简单到复杂、从无序到有序、从非平衡到相对平衡(从生态系统功能的角度)并且有机的物质与能量不断积聚的演进过程,这种演进以条件约束式的自发、自组织为特征(可谓“杂而不乱”)。当然,不同地质历史时期的无序、简单与非平衡的起点可能会很不相同。总体来看,条件约束型的演化过程或格局在精细的较低级生命层次(如物种)上趋向于更为随机性和不可逆性,而在宏观的较高级的生命层次(如地带性植被)上则趋向于更为决定性与可逆性。
从遗传的角度来看,生命对环境的适应与进化是物种基因库被生态力量—气候、地理或生物间关系(在相当程度上随机地)选择的结果。生命擅长用简单的创造原理制造表型复杂的生命,在经受大自然一次又一次的惊涛骇浪的洗礼之后,又顽强地(虽然或多或少随机地)进行着生命系统的重塑和进化。但是,物种基因库的复杂性(譬如人类的基因数多达2~2.5万个)从概率上来说使生命演化的精确重复几乎不可能。换句话说,如果地球再一次推倒重来,重新回到细菌的时代,再经过几十亿年,人类重新出现的几率就几乎不会存在,但也无法否认或许会演化出比人更聪明的物种。
著名的比利时古生物学家路易斯∙多洛曾提出“进化的不可逆”观点,认为基本的或然性就可以保证趋同绝不可以产生任何完全相同的东西。生物不可能抹去过去的痕迹。两个谱系的生物可能会发展出表面上明显的相似性,并适应相同的生命模式。但是生物含有非常多的复杂、特有的部分,根本不存在进化两次而导致一种完全相同结果的机会。进化不可逆,祖先的标记永远保留着,而且无论多么趋同给人留下多么深刻的印象,总是表面的(古尔德2008)。
生命演化的历史明晰地告诉我们,生命进化的总体趋势是必然的,但运行的具体轨迹却充满着随机、偶然和不可预测性。虽然地球生命系统在地质历史的长河中经受了若干次重大的洗礼,但又顽强地且不可逆地向新的生命系统方向挺进。
结语
自然界不会设计永生的生命,也不可能设计永生的物种。人类如同其它一切物种一样,其终极命运只有两个:要么彻底灭亡,要么消逝于变化之中(由有性生殖的本质所决定)。从这种意义上来说,解释生命进化的所有的“适应”或“完美”都是相对的或短暂的,周而复始的创造—进化—毁灭才是永恒的生命旋律。借用考克斯和穆尔(2007)的说法:“绝灭不是规范的一个例外,它是经常变化着的生命格局的一定特征”。
主要来源:
谢平. 2013. 从生态学透视生命系统的设计、运作与演化—生态、遗传和进化通过生殖的融合. 北京:科学出版社
(英文:Xie P. 2013. ScalingEcology to Understand Natural Design of Life Systems and Their Operations andEvolutions – Integration of Ecology, Genetics and Evolution through Reproduction. Beijing: Science Press)
引述该博文的相关内容时,请引用该专著。
https://blog.sciencenet.cn/blog-1475614-813621.html
上一篇:鸟瞰地球历史—生命系统在革新与跃升中演进
下一篇:全球气候变化之真相——淡定 or 惊慌
103 武夷山 梁高道 王文静 倪乐意 郝乐 王和云 袁军法 陈亮 国晓春 李尚春 张骥 柯志新 金刚 陈静 袁桂香 符辉 陈楷翰 唐汇娟 祝国荣 周琼 秦逸人 焦飞 王修慧 沈宏 张大文 李威 吴耀 赵丹丹 徐晓 喻海良 蔡雁 蒋滨 刘光银 朱朝东 黄永义 肖里 李宇斌 刘兵钦 张启峰 董洁林 张萌 肖振亚 李土荣 李红莉 赵正成 董侠 杜振亭 张云扬 梁洪泽 吴欣 陈泽宇 鲍得海 应行仁 许培扬 罗会仟 朱欢 吴世凯 陈琳琳 科学出版社 陈钢 刘学凯 赵斌 乔中东 吕乃基 唐常杰 张霄林 肖陆江 邓道贵 李亚平 覃剑晖 王江超 杨正瓴 蒋永华 余得昭 王伟 Lazi711 leihh fishstop0910 tangxin farniu kingz456852 liulanxiang lovenzaw truth21ct liguangyu2001 wangqinling sgcst hkcpvli rfm2007 tuoxun81 shenlu chenansb yyzhao1104 yanghua7788 biofans yunmu Vetaren11 peosim capenter rubygeng freebird2323 scr3322 sjtzcqn