博文
用数学统御生命世界的梦想
 精选
精选
||
地球上如此之多的生物物种是如何相依相伴地共存在一起的?我们面对的是一个寂静与喧嚣交融、幽雅与杀戮混杂的令人晕眩且无限纷繁的生命世界—时而蓓蕾绽放、生机盎然,时而刀光剑影、鲜血淋漓,时而旦夕祸福、朝生暮死……。那么,我们如何才能剥离生命世界中纷繁杂乱、变化万千的表象寻找出定量的普适性原理或法则?或许生命过程的模型化正是对这种普适性量化规律的一种抽象。自然界的生命系统的特性是随机的还是决定性的?能否具有可预测性?
从结构上来看,地球上的生命绝不是简单而杂乱无章地堆积或拼凑在一起的杂物(可谓杂而不乱),因为现存的生命系统其实是一个包含空间跨度极大的各种异常复杂的生命系统的集合,通过结构层次化形成了一种组织化与一体化的生命世界:即细胞→组织→器官→个体→种群→群落→生态系统→生物圈。而且,这些生命层次是一种通过一系列复杂的内在关系(如营养联系等)紧密而相当程式化地联结起来的包含式的构成关系,即低层次生命体结构化地构成了高层次生命体,这是生命系统的本质特征之—。用系统科学的说法,生物圈实质上是一个包含着不同等级亚系统的总系统,一层套一层并形成有序上升的等级。不同层次的生命系统是如何运作的?
动态是所有生命层次固有的另一个本质特征,因为现代的科学技术已经证实,一切形式或层次的生命系统都是开放的,它们的存在均实现于与外部环境永不停息的物质、能量和信息交换的动态过程之中,并不断的进行自我更新,而任何层次生命系统动态的停滞即意味着该生命系统的终结。而如何定量描述各种生命的动态过程一直是生命科学的重要目标之一,特别是理论生物学家和理论生态学家孜孜不倦的追求目标。
本文拟从不同生命层次的动态模型来透视生命系统的运作过程。虽然生命世界的细节充满着随机性、偶然性、复杂性与不可逆性,难以用决定性的数学模型(或许多数情况下只能估算概率)来描述,但这并不能说整个生命世界就只有一片杂乱无章的混沌,有限的确定性依然值得去探寻与挖掘。
一、酶促反应速率模型
微小的细胞通常只有在显微镜下才能观察得到,但它却是所有生物有机体的最基本结构材料,无论是怎样的庞然大物(如大象、巨鲸)。不同性质的细胞用不同的组合方式构建了数百万种之多的物种。从一个微小的细胞—受精卵发育成一个复杂的庞然大物就是细胞生长、分裂和分化的结果,而这些过程本质上是由一系列生化反应来推动的。
1. 酶惊人的催化能力使生化反应速率无可比拟
细胞内的生化反应控制着各种细胞的生长、分裂或分化,而生化反应的核心就是代谢。代谢维持着生物体的物质和能量交换过程,代谢的停止就意味着生命的终结。而生物体内的代谢反应是由一类特殊的生物大分子—酶(大多为蛋白质)来催化的,即通过一系列酶的催化作用将一种化学物质转化为另一种化学物质。酶(enzyme)的概念是由德国生理学家WilhelmKühne于1877年首先提出的。与其他非生物催化剂不同的是,酶具有高度的专一性,只催化特定的反应。
生命所展现出的惊人的适应、进化、扩增与繁荣无不与这种神奇的酶息息相关,正是酶才保证了生物体内为了获得(释放)能量和物质的高效生化反应的有条不紊地进行,许多酶可以将其催化的反应速率提高数百万倍。
酶可以在一秒钟内催化数百万个反应,例如,乳清酸核苷5'-磷酸脱羧酶(orotidine 5'-phosphate decarboxylase)所催化的反应在无酶情况下,需要七千八百万年才能将一半的底物转化为产物,而在这种脱羧酶的催化下,同样的反应过程只需要25毫秒(Radzicka& Wolfenden 1995)。酶的高效性是支撑生物个体的快速生长(快速细胞分裂)的基础。
2. 酶促反应速率的基本模型—米—曼方程
那么,酶促反应速率如何描述?早在1902年,法国物理化学家Victor Henri提出了酶动力学的定量理论(Henri1902);在此基础上,1910年,美国生物化学家LeonorMichaelis和加拿大医生Maud Menten提出了著名的描述酶动力学的Michaelis-Menten方程(Michaelis & Menten 1913)。
在大多数酶动力学反应中,酶促反应速度与底物之间的关系遵循所谓米—曼方程(Michaelis-Menten equation),其中V0=初速度,[S]=底物浓度,Vmax=最大速度,Km=是反应速度达最大反应速度一半时的底物浓度(米氏常数)(图1)。米—曼方程也是一种双曲线方程。
当[S]远小于Km时,[S]可以忽略不计,这时为一直线,即初始反应速度与底物浓度成正比,而当[S]远大于Km时,初速度V0等于最大速度Vmax。
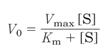
根据图1分析初始反应速度V0的增长速率的变化。不难看出,底物浓度[S]越小,单位底物浓度增加引起的V0增加的速率越快,之后,随着底物浓度[S]的不断增加,单位底物浓度增加引起的V0增加的速率越来越慢,当[S]远大于Km时,单位底物浓度增加引起的V0增加的速率趋于0,此时,初速度V0等于最大速度Vmax。

图1 初始反应速度对底物浓度的依赖性(引自Nelson and Cox 2004)
因此,米—曼模型是一种描述生化反应的初始速度V0随着底物浓度的增加从初始的线性增加向饱和(最大速率)转变的动力学过程。这里,Vmax与描述种群数量增长的逻辑斯蒂模型中的K(环境容量)有本质的不同,达到K时,种群增长速率为零。
在底物浓度很低的情况下,酶促反应速率与底物浓度呈现出一种正反馈关系,但是,随着底物浓度的不断增加,酶促反应并不会无限增加,而是趋向一个极限速率,这应该可以看成是对一个有限的细胞空间的一种适应。换言之,一些酶促反应速率虽然极快,但依然没能显示出像种群指数增长那样的无限性。
3. 生化反应的高效和精确性造就了差异极大的米氏常数
在米氏方程中,V0到达极值的历程会有所不同,而米氏常数Km就是与此相关的一种特征性参数。与Vmax不同, Km只与酶的种类有关,而与酶的浓度和底物浓度无关。各种酶和底物的Km差异很大(表1),细胞在设计生化反应系统时,也分配了差异巨大的Km(包括一种酶在不同的底物之间)。Km在生化反应中的主要意义如下:
1)Km反映了酶和底物之间的亲和能力,Km值越大,亲和能力越弱,反之亦然;
2)通过Km可以确定某一代谢途径中的限速步骤:一些代谢途径前一步反应的产物正好是后一步反应的底物,例如,EMP途径,限速步骤就是一条代谢途径中反应最慢的那一步,也就是Km值最大的那一步反应,该酶就是这一途径的关键酶。
3)Km可以用来判断酶的最适底物,某些酶可以催化几种不同的生化反应,叫多功能酶,其中Km值最小的那个反应的底物就是酶的最适底物。
表1一些酶和底物的Km
酶Enzyme | 底物Substrate | Km (mM) |
己糖激酶(脑) Hexokinase (brain) | ATP | 0.4 |
| D-葡萄糖D-Glucose | 0.05 |
| D-果糖D-Fructose | 1.5 |
碳酸酐酶 Carbonic anhydrase | HCO3ˉ | 26 |
胰凝乳蛋白酶 Chymotrypsin | 甘氨酰酪氨酸氨基乙酸 Glycyltyrosinylglycine | 108 |
| N-苯甲酰酪氨酰胺 | 2.5 |
ß-半乳糖苷酶 ß-Galactosidase | D-乳糖 D-Lactose | 4 |
苏氨酸脱水酶 Threonine dehydratase | L-苏氨酸 L-Threonine | 5 |
(引自Nelson & Cox 2004)
已知的可以被酶催化的反应多达数千种(Bairoch 2000),为了保证高效性,在通常情况下,酶对于其所催化的反应类型和底物种类趋向于高度的专一,这或许是一种不得已的进化选择过程,它增加了体内生化反应控制的精准性和高效性,却使这个系统变得异常复杂。当然,这从另一种角度来看,又使生命系统多样化,增加了变异和物种分化的潜能。一方面物种难以被复制,另一方面又变化无穷,这种特性可能是与这种系统的复杂性难以分割开来的。
遗憾的是,迄今为止还根本无法定量描述一个数以千计的各种酶(不同酶的Km值还变化极大)催化的极为复杂的反应体系的整体行为,因此更谈不上以此来推测个体或种群的行为。因此,如何将酶促反应模型与个体生长模型进行对接和整合将是生物学家未来面临的巨大挑战。
二、有机体整体代谢速率模型
为了探讨有机体整体的生化过程与生态现象的关系,只得撇开过于复杂的酶促反应的细节。由于能量转换是所有生命形式共有的必须过程,可以作为有机体整体代谢的重要指标,因此,整体代谢模型可能成为衔接生理生化与生态过程的重要桥梁。一些理论生态学家建立了质量—温度—代谢速率之间关系的异速模型(Brown et al. 2004),试图更为逻辑化地阐述了这种定量关系,而非仅仅像在第2章那里给出经验性的回归模型。
1. 有机体整体代谢速率与质量的关系
20世纪30年代, Huxley(1932)注意到一些关键的生命过程(Y)与生物体自身的质量(M)之间存在指数函数的关系:
Y = Y0Mb (3-1)
这里Y可以是代谢速率、发育时间等,Y0是一个与质量无关的归一化的常数,b是一个被称之为异速指数(allometricexponent)的常数,在大多数情况下为1/4(而不是1/3)的倍数,方程式(1)也称为异速方程式(allometric equation)。
Kleiber(1932)提出有机体整体的代谢速率(I)和身体质量(M)之间存在如下关系:
I = I0 M3/4 (3-2)
这里,I0是与Y0类似的常数。作为一个直观的例子,一头大象的整体代谢速率要比一只老鼠的高得多。
2. 有机体整体代谢速率与温度的关系
早在十九世纪后期,人们就认识到代谢速率以及几乎所有其它的生命活动速率都随温度的增加而呈指数增加。这种动态规律遵循所谓波尔茨曼因子(Boltzmann factor)或范特霍夫—阿累尼乌斯(Van’tHoff-Arrhenius)关系(Boltzmann 1872,Arrhenius 1889):
e-E/kT (3-3)
这里,E为活化能(activation energy),k为波尔茨曼常数(Boltzmann’s constant),k =1.3806488 × 10-23 J/K, T为绝对温度(K),E的单位为电子伏特(electronvolt,1 eV =23.06 kcal/mol = 96.49 kJ/mol)。这一关系仅适用于正常活动的温度范围,对大多数生物物种来说,位于0o - 40oC之间。
绝对温度T越高,式(3)的值就越大。作为一个直观的例子,在温暖的热带环境中的微生物活动与凋落物分解速率比寒冷的亚北极地区要快得多。
3. 身体质量和温度对有机体整体代谢速率的联合效应
在自然的情况下,生物质量和温度对生命过程(如代谢速率)的效应难于完全剥离开来,往往是联合作用。将式2和3相乘(Gillooly et al. 2001),即得到下列等式:
I = i0 M3/4e-E/kT (3-4)
这里,i0是一个与体积和温度无关的归一化常数。这就是一个考虑了生物质量和温度联合效应的方程式。
4. 关于代谢速率的若干概念
对等式(4)做适当的变换,得到下述若干有重要价值的概念。
(1)“质量矫正”的代谢速率
将等式(4)移项并对两边取对数,得到下式:
ln(IM-3/4) =-E(1/kT) + ln(i0) (3-5)
这里IM-3/4即所谓进行了“质量矫正”的代谢速率(‘‘mass-corrected’’metabolic rate),从式(5)可以看出,生物个体“质量校正”的代谢速率的自然对数与1/kT呈负的线性相关(也即与绝对温度正相关),代谢活化能E为斜率,归一化常数的自然对数ln(i0)为截距(图3-2A)。
(2)“温度矫正”的代谢速率
将等式(4)移项并对两边取对数,得到下式:
ln(IeE/kT) =(¾)ln(M) + ln(i0 ) (3-6)
这里IeE/kT即所谓进行了“温度矫正”的代谢速率(‘‘temperature-corrected’’metabolic rate)。
令人吃惊的是,从图3-2A可以看出,所有的生物类群的“质量矫正”的代谢速率都具有共同的斜率E ≈ 0.69 eV (1 eV= 96.49 kJ/mol),而截距C则出现差异:植物<单细胞生物<无脊椎动物<爬行动物<两栖动物<鱼<恒温动物。另一方面,“温度矫正”的代谢速率都具有共同的斜率E ≈ 0.71 eV,截距C为单细胞生物<植物<无脊椎动物<爬行动物<两栖动物<鱼<恒温动物。
从图2B可以看出,直线的斜率(0.71)接近3/4的理论预测值,而不同类群回归直线的截距即归一化常数的自然对数ln(i0)有所不同。纵观所有分类类群,针对基础代谢归一化的常数i0约有20倍的差异。
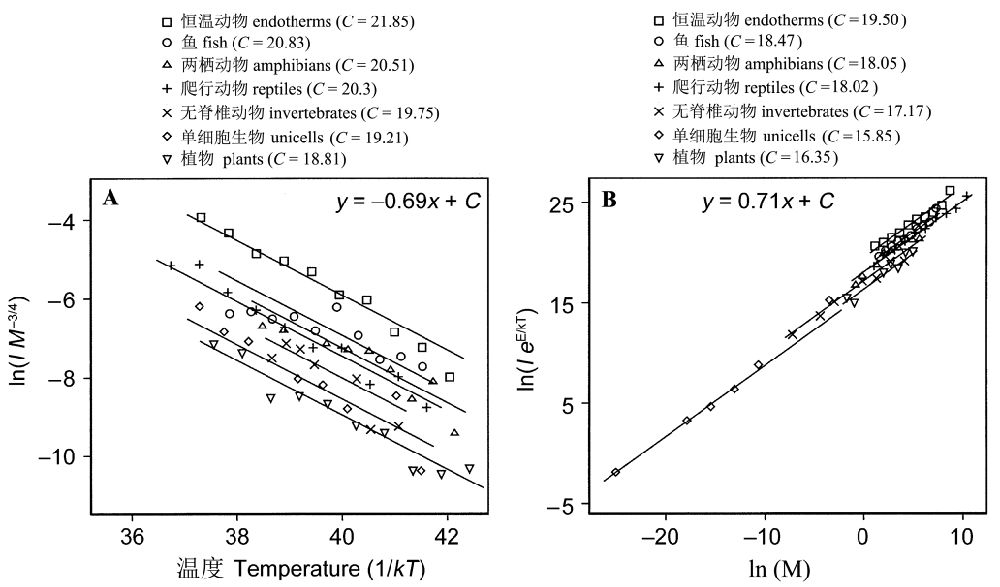
图2 从单细胞真核生物到植物到脊椎动物的若干生物类群,依赖于温度和质量的代谢速率。(A)质量矫正的代谢速率,ln(IM-3/4),单位watts/g3/4,与温度,1/kT,单位K之间的关系。(B)温度矫正的代谢速率,ln(IeE/kT),,单位 watts,和质量,ln(M),单位 g,之间的关系。变量为M(体重)、I(个体代谢速率),k(波尔茨曼因子),T(绝对温度,单位T)。E为活化能(引自Gilloolyet al. 2001)
(3)单位质量的代谢率
因为单位质量的代谢率B=I/M,式4可写成(其中∝为正比例符号):
B∝M-1/4e-E/kT (3-7)
与“质量矫正”的代谢速率相比,B还保留了质量的影响,也就是说单位质量的代谢率受到质量与温度的双重影响,与质量M负相关,而与绝对温度T正相关。
总体来看,基于异速方程所获得的关于质量—温度—代谢速率之间的关系与基于回归方程建立的经验模型的推测基本吻合。无论进行质量还是温度矫正,恒温动物的代谢成本均为最高,而植物和单细胞藻类最低。
三、细胞的体积及增长
与细胞内的酶促反应速率相比,人们对细胞体积生长的关注程度要小得多。与生物个体体积巨大的差异相比,不同物种间细胞体积的差异要小得多。
真核细胞一般大于原核细胞,大多数真核细胞的大小约为10-100 μm,而大多数原核细胞约为1-10 μm,当然也有极少数的单细胞真核生物大大超过这一范围,譬如一种阿米巴(Amoeba proteus)的原生动物长度可达1000 μm,还有一种单细胞的伞藻Acetabularia(图3)其柄和“帽”加起来可达到10 cm的高度(Verma & Agarwal 2005)。

图3 采自意大利Otranto的一种伞藻(Acetabularia acetabulum)(图片由Gianni Felicini提供)
多细胞生物的细胞大小一般在20-30μm之间,虽然也有少数例外,譬如鸵鸟的卵细胞的直径达到450px,人的一些神经细胞有近1 m长的“尾巴”或轴突,马尼拉麻的纤维细胞长达100 cm(Verma & Agarwal 2005)。
显然,无论是植物还是动物,都没有采取靠增加细胞体积来实现个体生长的策略,而是通过不断的细胞分裂、堆积和分化等来实现个体的生长。细胞体积只是呈现一种简单的周期性变化:即细胞分裂形成的新细胞,最初体积较小,只有母细胞的一半,但它们能迅速合成新原生质,细胞随之增大,到母细胞一般大小时,便可继续分裂,如此循环往复(图3-4)。因此,对细胞的增长无需要像对个体或种群那样用复杂的模型进行描述。
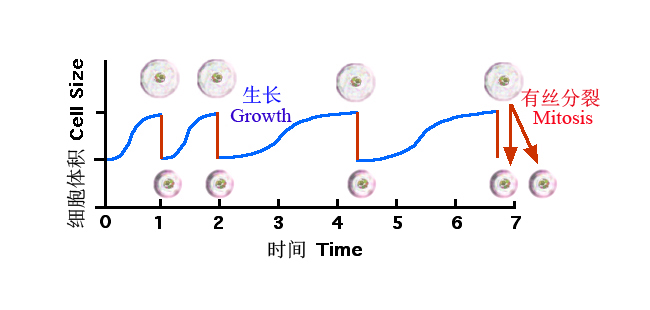
图4 细胞体积随细胞分裂的变化(引自Wikimedia)
四、个体生长模型
生命个体由单细胞或多细胞组成。对单细胞生物(如原生动物),细胞体积的增长等于个体体积的增长,如前所述,一般细胞体积的变化范围十分有限。而对多细胞生物来说,个体的生长主要建立在细胞数量的快速增殖与堆积之上,如动物的一个小小的受精卵可以长成一个庞然大物,植物的一粒种子可以长出一颗参天大树。
地球上有如此之多的生物物种,不同物种的个体生长模式也不可能完全一样,甚至同一物种的不同个体以及在不同的环境条件下都有可能不同。但至少有一点可以肯定:与可能在相当的空间范围内无限增长的种群不同,个体生长(体长或体重的增长)是有限的。那么,能否用数学模型来描述个体的一般生长过程?
1. 常见的个体生长模型
由于生产实践的需求,人们历来十分重视动物个体的生长模型研究,也发现了普遍的规律:动物在生长良好的情况下,呈现出一种典型的S型生长曲线(将体重对年龄或时间作图)。为此,动物学家们找来了各种各样的数学公式来进行描述,譬如López(2008)列举的描述动物生长的方程式多达40多种。
渔业管理实践推动了渔业生物学家对各种鱼类的个体生长规律的广泛研究,在渔业文献中可见到各种各样的数学方程式用于描述鱼类的生长,其中一些常见的模型如表2所示。
在表2列举的各种生长模型中,S表示个体大小(体长或体重),t表示年龄或时间,t0为积分常数,S∞为极限大小(如果存在的话),而a、b、c和n为待确定参数;描述无限生长的方程式(如指数函数、幂函数)不趋向于极限成体大小,而那些描述有限生长的方程式(Gompertz、逻辑斯蒂型、vonBertalanffy、Richards)则趋向极限成体大小。
表2 常见的生长方程

(引自McCallum 2000)
2. 主要生长模型的参数特征比较
在个体生长模型中,最常见的几种模型为von Bertalanffy模型、Gompertz和逻辑斯蒂模型。vonBertalanffy模型(von Bertalanffy, 1938)如式(3-1)所示,其中lt为个体在时间t的长度,l∞为极限体长(或称渐进体长、最大体长),K表示趋近极限体长的相对生长速率,t0为允许年龄0时非零的外插体长的转换系数
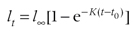 (3-8)
(3-8)
常常加一个异速生长参数b,特别是用重量而不是体长表示时。von Bertalanffy生长曲线的一般形式如图3-5A所示。值得注意的是,等式(3-8)不能生成S型曲线,而等式(3-9)则可以。
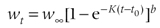

图5 生长曲线的例子,所有曲线的S∞ =100,大小和年龄的单位为任意的(引自McCallum 2000)
在生长模型中,拐点的存在与否及位置是这些曲线的重要特性之一。在数学上,拐点是凸曲线与凹曲线的连接点。在生长模型中,如果拐点存在,则拐点前为生长加速区(近似于指数生长),拐点后为生长减速区。
其实,由于个体生长的有限性,任一物种的生长都会趋于一个极限体长(此时生长速率为零),只是到达极限体长的生长轨迹可能略有不同,特别是拐点出现的位置可能不同,这可能反应了物种不同的生存策略,如适应于资源的可利用性或其它环境条件。生物不需要也不可能有统一的生长模式,变化的生长模式显然是对自然界千变万化的一种适应。
从图3-5可直观的看出,vonBertalanffy曲线(A)无拐点,表示初始生长速率最快,以后逐渐降低;vonBertalanffy曲线(B)的拐点约在S=1/3 S∞处;而逻辑斯蒂曲线(D)的拐点约在S=1/2S∞处;Gompertz曲线(C)的拐点位置也要低于Logistic曲线(d)。拐点位置越低,越早进入生长减速期。因此,拐点的位置是区别这三种生长模型的特征性参数之一。
3. 描述鱼类生长的vonBertalanffy模型的生长速率K和极限体长l∞的比较
von Bertalanffy模型在鱼类生长的研究中应用最为广泛,该模型的二个重要参数—极限体长l∞和相对生长速率K也是被探讨的焦点问题之一。Henderson(2006)依据世界鱼类数据库的资料,比较了热带和非热带鱼类的K值(图3-6)。当然,这是近似值,因为许多种无论是生长速率还是极限大小在其地理分布区甚至年际间都有宽幅的变异。很显然,生长速率超过0.8的比例,热带鱼类比非热带鱼类要高得多,这表明热带鱼类趋向极限体长要比非热带鱼类快得多。
热带鱼类的l∞的分布情况见图3-7,不难看出,极限体长在10-70 cm之间的鱼类占绝大多数,而超过1m的鱼类十分罕见。很显然,热带鱼类的生存策略—以小个体来实现快速生长。

图6 源自世界鱼类数据库的热带(A)和非热带(B)鱼类的von Bertalanffy生长常数K的频度分布(引自Henderson 2006)
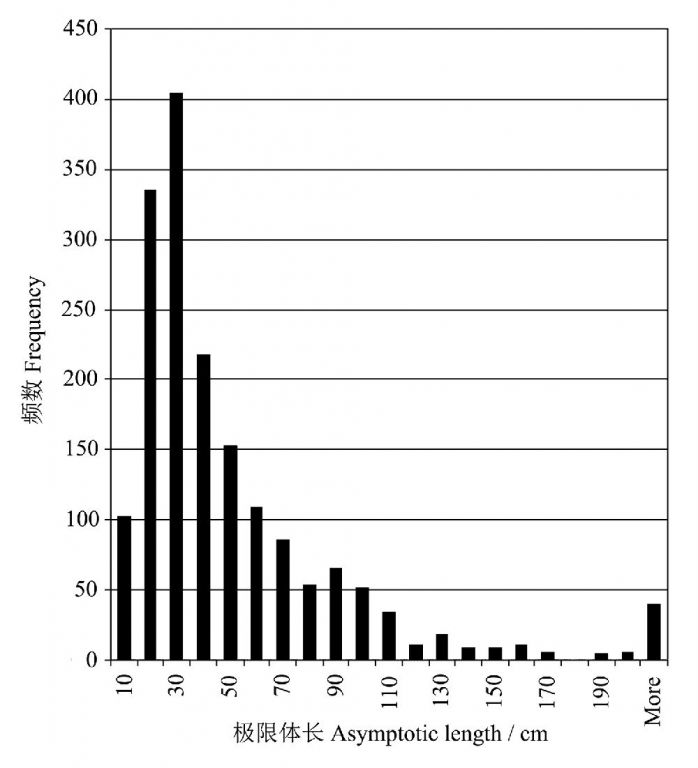
图7 鱼类数据库中的热带鱼类的极限体长l∞的分布(引自Henderson 2006)
4. 主要生长模型之间的数学转换
从纯数学的角度,一些模型之间随着参数的变化可以互相转化,通过这种变化,可以加深对一些模型特性及相互间关系的认识。Thornley(2008)以动物生长模型为例,探讨了不同模型之间的数学联系。首先可用下述方程式描述动物生长:
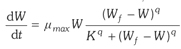 (3-10)
(3-10)
这里,W为重量(kg),μmax为最大生长速率(day-1),K类似米氏常数,Wf为最终(极限)重量。
1)当q=0,无限生长;当q=1时,米-曼生长;
2)当Wf→∞时,得到特定生长率为μmax的指数生长,
3)当K→∞,μmax→∞,μmax/Kq =常数c时,得到一个修改的逻辑斯蒂方程式:
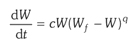 (3-11)
(3-11)
如果q=1,则等式(2)为逻辑斯蒂方程

图8 类似于米-曼S型底物限制的生长(式1)。所有曲线均具有同样的初始重量(W0 = 1),渐进的Wf = 100,以及初始斜率(调整μmax以满足这一点)。(A)保持q=1,而使K变化;当Wf = 1 × 1010为无限的指数生长,μmax = 0.18165;当K = 1 × 109和μmax= 0.18349 × 107 = 0.18165K/99时,得到逻辑斯蒂生长。(B)保持K=10,使q变化。实心圆圈表示拐点(引自Thornley2008)
注意从逻辑斯蒂模型到指数模型转变过程中拐点的变化(图8a),即逻辑斯蒂曲线的拐点位于渐进线高度1/2的位置,向指数模型推移的过程中,拐点的位置逐步上移,最后在指数模型中消失。而对小的q值,指数生长占据绝对优势直到接近渐进线,而对大的q值,紧跟着指数生长的是向渐进线的较缓慢的接近(图8b)。
总的来看,个体生长一般呈现具有拐点的S型曲线,拐点前可看成无限生长区,在此期间幼体生长处于加速阶段,拐点之后为减速区,并最终停止生长。
依我看来,可将个体的生长看作有限环境下种群增长模式(逻辑斯蒂模式)的一个缩影,若以细胞数量的增长来考虑(这里个体的极限体积类似于细胞的环境容量),就不难理解了。即与种群一样,个体也是从初始的无限的指数增长开始,接着穿越拐点后进入减速,最后停止增长。极限长度和拐点便成为了个体增长模式的两个重要参数。
五、单一种群的数量变动—始于无限,终于有限
种群如何变动?在巨大的地球系统中,数百万种生物物种在各种环境千变万化、生物物种间错综复杂的相互作用以及在种群的边界几乎可以无限缩放等的背景下呈现出的变动模式是无限的。这就给如何用数学模型来描述种群的变动规律带来了极大的困难。
幸运的是,人可以作为一个特殊的种群,还有在一些小而简单的实验生态系统以及一些边界清晰的岛屿中的种群边界也易于确定,等等,这些给种群数量变动的模型研究提供了绝佳的契机。在描述种群增长的数学模型中,最著名的无限与有限模型均是最先用于人口学—1798年马尔萨斯在描述了人口呈几何级数增长的模型,1834年Verhulst首次用逻辑斯蒂函数描述人口的有限增长。
1. 种群增长模型—无限寓于有限之中
任何一个物种都有使其种群无限增长的内在潜力(即所谓指数增长),这是生命设计的基本原理,失去这种特性的物种其命运就是灭绝。而环境的有限性将阻止这种趋势的无限发展,使其趋于一个平衡值K(即所谓的环境容量),种群大小N越接近K,环境阻力越大(图9)。
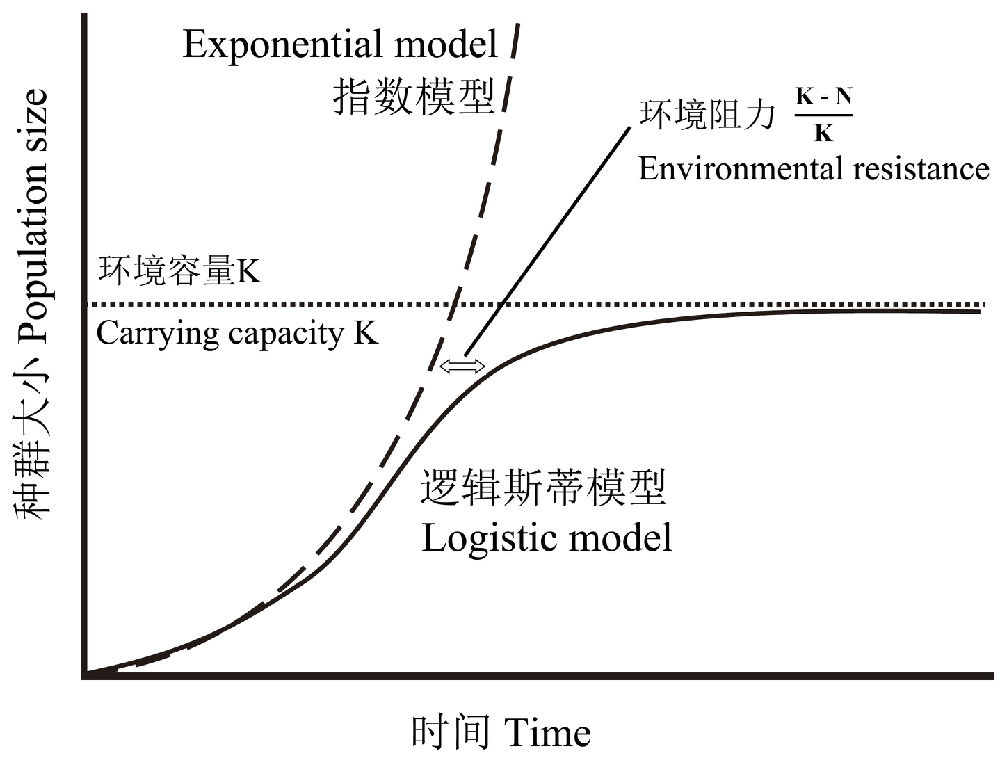
图9 描述种群增长的指数和逻辑斯蒂模型。指数模型描述一个无限增长的种群,而逻辑斯蒂模型描述一个种群趋向一个环境容量(K)的渐进线
再来看看指数模型和逻辑斯蒂模型种群的增长速率的变化。在指数增长模型中,随着时间t的增大,种群数量P的增长速率(即单位时间增加引起的种群增加量)也越快,t趋于无穷大时,P的增长速率也趋于无穷大。而在逻辑斯蒂模型中,开始随着t的增加,P的增长速率增加,当P为K/2时,P的增长速率达到最大,之后,t的进一步增加,P的增长速率不断减慢,最后趋于0。
值得注意的是,在P达到K/2之前,P的增长近似于指数增长,即在近乎理想的条件下,逻辑斯蒂模型的初始阶段为指数增长,而K/2是种群增长速率的拐点。因此,逻辑斯蒂模型的本质是一种描述种群数量从初始的指数性(无限)增长向环境容量逼近的动力学过程。
2. 逻辑斯蒂增长—实验种群的常见模式
逻辑斯蒂方程式被用来描述自然界广泛存在的有限种群增长,从单细胞的酵母、小型的浮游动物到大型的脊椎动物,既可以是实验种群,也可以是自然种群。一些经典实验表明,在恒定和有限的环境中,很多实验种群(酵母、无脊椎动物等)的数量增长可以用逻辑斯蒂方程式来描述。
1)单细胞生物—酵母、草履虫
酵母是一种单细胞的真核生物,也是一种广泛使用的实验动物。在培养条件下,一个经典的酵母种群逻辑斯蒂增长案例如图10所示,拟合下述逻辑斯蒂方程式:

获得参数K = 664.3,a = 4.2017和rm = 0.5384(Neal 2004)。

图10 酵母种群的逻辑斯蒂增长,生物量数据(单位未提供)源自Carlson 1913(仿Neal 2004)
草履虫是一种单细胞的原生动物,也是有名的实验动物。在培养条件下,草履虫种群的增长(图11)没有像上述酵母那样呈现一条十分完美的逻辑斯蒂曲线,而是在K值附近上下波动,拟合逻辑斯蒂模型后得到参数K = 202,a = 5.1和rm= 0.74。

图11 实验室培养条件下草履虫种群的逻辑斯蒂增长,数据源自Gause 1934(仿Neal 2004)
2)多细胞生物—昆虫和甲壳动物
果蝇是著名的实验动物,在遗传学研究中立下了赫赫战功。图12a是黑腹果蝇种群的增长曲线,图12b是浮游甲壳动物—多刺裸腹溞的种群增长曲线。不同遗传结构的果蝇种群的K值明显不同,而多刺裸腹溞在不同温度条件下的K值也不同。显然,K既与种群内在的遗传与生理特性有关,又依赖于基本的生存环境,反应了二者之间的一种平衡。

图12 实验种群的增长曲线:(a)黑腹果蝇种群:(i)野生型,(ii)包括残翅在内的5个隐形突变的杂合或纯合型个体,(iii)有一半(i)的野生型;(b)多刺裸腹溞种群:三种不同的温度条件。转载于Hutchinson(1978)。
3. 指数增长—惊人的爆发vs 惊人的崩溃
1)引入St Paul岛的驯鹿—从指数增长到崩溃
1911年,4头雄性驯鹿和21头雌性驯鹿被引入到位于白令海的St Paul岛(面积106 km2),到1938年,驯鹿种群增加到约2000头,成为种群指数增长的经典案例(Krebs 1985),接下来种群逐渐崩溃,至1950年,驯鹿只剩下6头(图13)!运用1911—1940年期间的数据,估算的种群增长速率r = 0.167(McCallum2000)。

图13 在1911-1950年期间,St Paul岛上驯鹿种群的增长与崩溃(仿McCallum 2000)
2)世界的人口—还在延续指数式疯长
虽然人类的进化试图将自己从普通动物界区分开来,虽然人类也已经主宰了整个世界,但人类依然脱离不了自然的动物属性。人类是一个大的动物种群,从其起源中心(一般认为在非洲)开始扩散,现在已经遍布了全世界。
在人类历史的大部分时期,人口都很少,快速的人口增长发生在近代,1850年,世界人口达到10亿大关,然后开始快速增长(图14)。近千年的人口增长呈现出经典的指数增长模式,但这种趋势决不可能无限持续下去,除非人类自我控制,否则St Paul岛上驯鹿种群崩溃的命运终究有一天会降临到人类的头上。

图14 世界人口的指数增长曲线。注意在最近200年的快速增长(引自Chiras1991)
为什么近千年人口会如此持续地进行指数增长?这主要源自出生率和死亡率平衡的打破,特别是由于医学的进步使人类的死亡率大大降低。我很同意道金斯(1981)的观点:乞灵于农业科学的进展—“绿色革命”之类,是无济于事的,增加粮食的生产可以暂时使问题缓和一下,但肯定不可能成为长远之计;如果放任人口自由增长,限制人口的“自然方法”就是饥饿!
六、先天的出生,后天的死亡
种群的增长绝不仅仅取决于出生,它体现了出生与死亡之间的一种平衡(当然在一个开放的系统中还包括迁入与迁出),这与个体的体长或体重的变化有本质的差异。死亡率对种群的影响有时比出生率显得还要重要,譬如人类的暴发型增长(图14)就起因于死亡率的下降。一般来说,一个物种的出生率是一种先天的或固有的属性(当然从进化上看也是生态对策与自然选择的产物),而死亡则是一种在后天更易于改变的特性(当然,这也是相对的)。两种不同的生态对策(r和K)其实反应了它们对出生和死亡的相对投入,以及在进化上选择的不同方向。
常常用存活率来计算死亡率,但是,一般来说,对野外种群存活率的确定往往比确定出生率要困难得多。每个物种的生死虽然受制于诸多的因素,也呈现出相当大的变异性,但是还是能归纳出一般的模式。生态学家常常用存活率曲线(survivorship curve)来描述某个物种或群体(如雌/雄)在每个年龄存活个体的数量和比例。
1. 存活率曲线—不同的死亡策略
早在20世纪初,Pearl(1928)就把成活率曲线归纳为三类:I型—死亡率集中在极限寿命的结束时期,II型—在不同年龄的死亡概率保持恒定,III型—早期死亡率大,而剩下的个体接下来有高的存活率,这一类型的物种产很多的后代,但是最初仅有少数个体存活下来,而一旦个体达到某一体长,它们的死亡率就降低且较为恒定,这一存活率曲线在自然界的动植物最为常见(图15)。
I型—可能是富裕国家人类的缩影,也见于动物园里的宠物或养殖场里的生命;II型—可能适合于许多植物种群的被埋藏的种子库;III型—可见于许多海洋鱼类,能产数百万卵,但没几个可活到成体。当然,这只是一种理想的划分,实际还会存在许多中间类型。
存活率可用指数方程或Weibull方程描述:指数的存活率方程为F(t) = e-ρt,Weibull的存活率方程为F(t) = exp[-(ρt)κ]。

图15 成活率曲线的划分:(a)为线性尺度,(b)为对数尺度。图形根据Weibull存活率函数绘制,I型:κ = 5,ρ = 0.00736;II型:κ = 1,ρ = 0.03454;III型:κ = 0.2,ρ = 78.642。选择ρ值以使年龄200的存活率的每个κ值标准化到0.001(引自McCallum2000)
2. 人类的存活率曲线—趋向极限死亡的转变
在动物界,由于存活率曲线的变化导致种群数量趋势发生根本改变的最好例子之一就是人类。为何在人类历史的大部分时期,人的种群数量一直保持较低的密度,而近千年来才开始加速生长(图14)?存活率曲线类型变化所反映的人类寿命的延长是一个重要的因素。从图16可以看出,1977年日本人的存活率遵循典型的I型曲线,而石器和青铜时代的人类则是呈现典型的II型存活率曲线,在这二条曲线之间则是一些过渡类型。
按图14下去,不久的将来地球上很快就会人满为患,要么面临马尔萨斯式的解决方式,要么人类严格控制自身的生育。

图16 人类的存活率曲线的历史变化(引自Environment Agency, Government of Japan1995)
图17为美国1900-2000年期间存活率曲线的变化,1900年的曲线介于I、II型之间,而2000年则为一个典型的I型曲线。从人类的例子不难看出,某一物种存活率曲线的改变,将会对种群的动态产生极大的影响。

图17 美国在1900-2000期间特定年龄的存活率(引自Rogerset al. 2005)
七、二个种群间的相互作用模型
任何物种都存在于一定的生态系统之中,有的生态系统中包含的物种相对简单(如农业生态系统),有的则十分复杂,如热带雨林中的一棵大树可为数以万计的物种提供栖息场所(霍兰2000)。生物之间的相互关系也是多种多样,有直接的,也有间接的。有竞争(对光、营养、食物、空间等),有残杀,有寄生,还有互惠……,这些关系一级又一级、一层又一层错综复杂地交织在一起,形成了一种极为复杂的网络体系。生物学家并不擅长从这种过于复杂的相互关系中从动态变动的视角来厘清头绪,更谈不上模型化。
而理论生态学家热衷于也擅长这种研究,它们的特点是能够恰如其分的对关注的对象进行简化(而知道过多细节的生物学家往往不知道该从何处下手),譬如最初的杰出工作就是开始于两个种群的相互作用—捕食者-猎物(或寄生虫-宿主),这也是种群间的基本关系之一,也是自然生态系统中食物链得以存在的基础。他们先假设一个最简单的生态系统—只有捕食者-猎物,然后用模型来描述它们之间的相互作用以及所引发的动态过程。
1. 经典的Lotka-Volterra方程—永无止境的周期性波动
最早提出描述两个相互作用物种的种群动态的数学模型的科学家一个是美国学者Alfred James Lotka,另一个是意大利学者Vito Volterra。Lotka是一名数学家、物理化学家和统计学家;Volterra是一名数学家和物理学家。他们俩彼此独立地得到了两物种相互作用的数学方程。
Lotka于1925年发表了经典著作“物理生物学原理”(Elementsof Physical Biology, Lotka 1925,该书1956年再版时更名为Elements of MathematicalBiology)。在该书中,他发现了Ronald Ross的疟疾方程与逻辑斯蒂方程的相似性,评论了Thompson W. R. 从数学上分析寄生虫对宿主影响的研究,发现Thompson的公式不适用后,他自己发展了一对微分方程,用来描述寄生虫(或捕食者)对宿主(或猎物)的影响,这些方程产生了二个物种种群的周期性波动(McIntosh 1985)。
与通过扩展逻辑斯蒂方程到二个种群来发展捕食者—猎物模型不同,Volterra(1928)则借用了质量作用的化学原理,即他假定种群的响应与其生物量或密度的产物成比例(Berryman 1992)。
Lotka-Volterra模型又称为捕食者-猎物方程式是一对一元、非线性的微分方程式:
 (3-12)
(3-12)
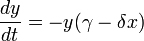 (3-13)
(3-13)
这里
· y为某一捕食者的数量(如狼);
· x为其猎物的数量(如兔子);
· dy/dt和dx/dt表示二个种群在单位时间内的增长率;
· t表示时间;
· α、β、γ和δ为代表二个物种相互作用的变量。
从方程式3-12和3-13可以看出:
αx为在没有捕食者时猎物的增长速率,因此在没有捕食者存在的情况下,猎物的增长为dx/dt = αx,积分后得到xt= x0eαt,即呈无限的指数式增长;
βxy为猎物被捕食者攻击所引起的死亡率;
δxy为捕食者后代的生产速率,与被捕食的猎物的数量直接相关;
γy为捕食者在没有猎物存在的情况下的死亡率,因此,在没有猎物存在的情况下,捕食者的死亡遵循dy/dt = -γy,积分后得到yt = y0e-γt,即呈指数式衰减。
Lotka-Volterra方程所揭示的就是捕食者和猎物之间相互作用导致二个种群的一种普遍的变动模式—周期性振荡(May 1976),即高的猎物密度往往产生高密度的捕食者,而捕食者的增多又会导致猎物数量的减少,后者又使捕食者密度降低,而捕食者的密度降低又导致较大的猎物密度,如此循环往复。寄生虫与宿主之间的相互作用也类似。
2. 两个相互作用种群的周期性振荡案例
无论在实验条件下还是在自然条件下,都可观察到Lotka-Volterra方程所描述的两个相互作用种群周期性震荡的案例。
1)实验系统中的寄生虫—宿主
如果一个实验系统,只含有捕食者和猎物,就容易观察到这种相互作用。日本学者提供了这样一个经典的例子:Utida(1957)在实验培养条件下,观察了绿豆象(Callosobruchuschinensis)和一种寄生蜂(Heterospiliusprosopidis)的相互作用,在25个世代时间中,出现了4-5个周期性波动,宿主和寄生虫的密度峰值总是交替出现,且宿主的密度峰值总在前面出现(图18)。
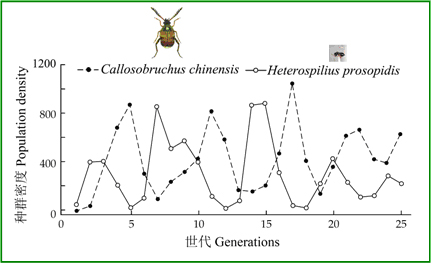
图18 Utida(1957)的绿豆象(实线)和一种寄生蜂相互作用(虚线)呈现出周期性波动(仿May and McLean 2007)
2)自然系统中的捕食者—猎物
在自然生态系统中,一个经典的捕食者—猎物的周期性震荡案例就是北美的北方林中的猞猁和兔的故事,猞猁和兔的数量主要根据哈德逊湾公司(Hudson Bay Company)的皮毛贸易的历史记录,从猞猁和兔长达90年的数量变动可以看出平均大约10年出现一个波动周期,种群的高峰总是兔在前,猞猁在后(图19)。

图19 猞猁和兔相互作用(Elton, 1924)每隔9-11年就出现波动一个周期(Stenseth et al. 1997)(仿May andMcLean 2007)
这种二个物种相互作用导致种群周期性震荡的现象在很多自然生态系统中应该也是真实存在的,但在大多数现实的生态系统(如森林、草地、湖泊等)中,往往是许多(少则数十,多则成千上万)物种错综复杂地共存在一起,即各种食物链交织在一起形成复杂的网络,不说几十种,如何用数学模型描述相互作用的3~4个物种的种群动态都是一件极其困难的事情。
在自然的生态系统中,两个物种间的相互关系往往还会波及或影响到更多物种的命运,引发一连串生态链式反应。布查纳(2001)讲述了这样一个动人的故事:“20世纪70年代……成群的野兔吞噬着上万英亩肥沃的良田。幸运的是,英国政府已准备了一套安全方便的解决办法……通过引进兔瘟,他们可以控制野兔数量……瘟疫确实使野兔数量在几年内急聚下降……随着饲养动物和吃草的野兔的减少,英国南部地里的草长得比以往更高了……但是有一种叫做MS的蚂蚁很快大批死亡了,因为它们在矮草中繁殖迅速,但在较高的草中,生命力却不很强。这种蚂蚁与一种叫做MA的蓝色大蝴蝶有一种特殊的关系。当这种蝴蝶产下卵后,蚂蚁把它们运进洞穴,孵化出幼虫,并一直将其培育成成虫。不幸的是,在20世纪70年代这种蝴蝶的种群已经岌岌可危了,当蚂蚁数量下降时,这种蝴蝶的数量便也骤然下降。兔瘟的引入使草增高、蚂蚁减少,并使这种美丽的蓝色蝴蝶在英国完全绝迹了”。不可否认,这种生态链式反应给物种间相互作用的模型化带来了极大的困难。但是,这并不意味Lotka-Volterra方程毫无价值,生态系统中再复杂的相互作用很多也离不开这一基本关系,即也是运用这一简单关系交织而成的。
八、不同生命层次的运动—难觅统一的动态模式
1. 酶促反应速率—既高效又专一,依赖于底物浓度
在细胞水平,酶促反应速率只依赖于底物的浓度,与时间和产物均无关系。因为酶本质上只是一种催化剂(虽然它高效而专一),它本身在反应过程中不被消耗,也不影响反应的化学平衡。酶以近乎无限的高效性(可比普通的化学反应速率提高数百万倍)加上专一性控制着细胞内极为复杂的代谢过程快速而有条不紊地进行,支撑着细胞的快速分裂,虽然它们未能呈现数学上的无限性(可能限于一个有限的细胞空间)。由于细胞内酶种类繁多,如何评价酶的整体行为及其对细胞增殖速率的影响都极为困难。
令人惊讶的是,细胞中的这种酶促反应动力学模型似乎只是一种底物调节型,并未受制于产物的调节,这或许是因为一般情况下酶促反应的产物会被(其它反应)快速利用或清除,或许昭示着生命系统正是在一种平衡中实现高速运转。但是,一旦有害产物得不到及时清除而出现堆积的话,很快就会导致生命系统的崩溃,这也从另一种角度昭示了个体生命系统潜在的脆弱性。
2. 个体和种群的增长—从无限到有限,依赖于时间与自身质(数)量
而个体的生长就与酶促反应完全不同了,它是时间和自身质量的函数。个体的生长十分类似一个有限环境中细胞数量从无限的指数增长开始逐渐过渡到零增长(趋于极限体长)的逻辑斯蒂过程,虽然它常用von Bertalanffy方程来描述(该方程的拐点比逻辑斯蒂更早出现,即意味着更短的指数增长期,相对于极限个体大小来说)。
与个体生长类似,在有限环境容量中,种群的逻辑斯蒂增长为时间和种群数量的函数,它由无限的指数增长开始,接近环境容量的一半时开始减速,最后在环境容量附近增速趋于零(出生与死亡相等)。它与个体生长的模型应该最为接近,但迄今为止还没有人关注两者之间的可能关系以及如何将两种动态模型进行对接与融合。
结语
地球生命系统形成了一个包含式的结构体系:细胞→组织→器官→个体→种群→群落→生态系统→生物圈。如何定量描述各种生命系统的动态过程?广泛关注的动态模型主要针对酶促反应、有机体整体代谢、个体生长和种群增长的动态过程。
酶促反应惊人的速率可能是生命世界得以在地球上如此繁荣的本质机制之一。单位质量的代谢率与质量负相关,与绝对温度正相关,恒温动物的代谢成本均为最高,而植物和单细胞藻类最低。个体和种群的增长模式基本类似,虽然前者常用von Bertalanffy方程,而后者常用逻辑斯蒂方程,都是始于无限,止于平衡。存活率曲线的改变可以显著影响种群的动态(如人类),因此,提高存活率也是物种进化的方向之一。
显然,不同层次的生命系统(如细胞、个体、种群)具有不同的结构特征、调节机制和动态模式,其稳定维持(当然任何活结构都不可能永恒地稳定下去)的机制也不尽相同。负反馈(如捕食者与猎物系统)是一种平衡与稳定机制,而无限的正反馈(如种群的指数增长)将会导致系统失稳乃至崩溃。细胞与个体通过复杂的自我更新、适应与调节来维持稳定运行,而种群则在与外部环境(生物的或非生物的)永不停息的相互作用中生存、发展与演化。
总体来看,生命在对动态过程进行设计时,赋予了不同生命层次相对独特的动态模式,虽然生命层次在结构上是一种包含式的构成关系,但至少到目前为止,科学家对这些不同层次的动态过程还难以(或确切地说还未能)成功进行模型对接式的简约叠加。
主要来源:
谢平. 2013. 从生态学透视生命系统的设计、运作与演化—生态、遗传和进化通过生殖的融合. 北京:科学出版社
(英文:Xie P. 2013. ScalingEcology to Understand Natural Design of Life Systems and Their Operations andEvolutions – Integration of Ecology, Genetics and Evolution through Reproduction. Beijing: Science Press)
引述该博文的相关内容时,请引用该专著。
电子版下载:http://wetland.ihb.cas.cn/lwycbw/qt/
https://blog.sciencenet.cn/blog-1475614-812047.html
上一篇:审读•覃思——生命是什么?
下一篇:鸟瞰地球历史—生命系统在革新与跃升中演进
116 陈亮 吴世凯 柯志新 郝乐 梁高道 祝国荣 袁桂香 符辉 王荣林 倪乐意 焦飞 王德华 王文静 刘洋 国晓春 石彭灵 熊倩 张学振 沈宏 李亚平 蔡雁 张能立 梁作论 应行仁 黄永义 朱朝东 刘胜强 王伟华 陈钢 金刚 肖里 李威 袁军法 王继华 陈苏华 赵丹丹 李帅军 邓道贵 史彭慧 王秉钧 鲍得海 徐传胜 贠可力 张大文 刘兵钦 张萌 谷安辉 白悦辰 靖新 廖晓琳 李土荣 陈波 梅志平 余国志 陈文峰 吴耀 谢维 覃剑晖 唐汇娟 赵斌 谭向峰 程起群 杨森 张凯军 郑小康 杨文斌 陈静 王明明 彭勇 李万春 李莉 强涛 朱欢 葛峰 张霄林 蔡裕华 董洁林 夏鹭 蔡志全 余得昭 李红莉 董云伟 杨新国 leihh Xiawulai fishstop0910 wangqinling huahen134 yyzhao1104 shaoyunpeng shenlu sgcst hkcpvli rfm2007 lipingren fangfeng1979 ttqq tangxin farniu capenter biofans chenansb scr3322 Vetaren11 rubygeng wanghua4 yunmu lingling101 pangxinghong88 laijianshan Lazi711 zkyshj rosejump sjtzcqn lovenzaw yanan2015