博文
气候干暖化引起的蓝藻增加危及非洲象等大型动物
 精选
精选
|
蓝藻——“非寻常嫌犯”抑或是“连环杀手”?2020年5月至6月,非洲博茨瓦纳至少330头非洲象(Loxodonta africana)突然死亡。这一事件迅速引起了全世界的关注和担忧(详见New York Times, Washington Post, Chicago Tribune 和 BBC News等主流媒体的报道见附件text S1)。最初,病毒和细菌被认为是造成这一悲剧事件的最可能的原因,并排除了恶意投毒、偷猎、饥饿和炭疽病等的可能性[1]。航拍图像和实验室检测结果显示,生物毒素或疾病可能是罪魁祸首(对各种可能原因进行的分析见附件text S1),而大象饮下了富含有毒蓝藻的水则是其中最可能的原因之一(图1)(有关蓝藻、藻毒素和富营养化等概念的更多信息见附件text S2)。

图1. 在一个长满蓝藻的水坑边死去的非洲象
(图片源自https://www.reuters.com/article/us-botswana-elephants-idUSKCN24N2JK)
目前,人们对蓝藻毒素与大象大规模死亡之间的因果关系仍然存在疑问。在理想情况下,要确定蓝藻毒素中毒是否真的是引发这次神秘的大规模大象死亡的直接原因,应当对所涉及水域范围的水质、蓝藻种类和藻毒素浓度的样本进行详细检测,同时结合组织病理学分析。然而,受限于当前全球新型冠状病毒 (2019-nCoV) 大流行的影响,再加上大规模死亡地点的偏远,使得现场测量变得非常困难。此外,当这一事件开始引起人们注意时,大多数水域已经干涸,许多死亡大象身体已经严重腐烂。本研究收集了非洲大陆各个地区各种水域有关蓝藻毒素浓度的文献数据,并查阅了非洲大陆蓝藻毒素致死事件的历史记录,结合已有各种哺乳动物急、慢性毒性实验资料进行了回顾性的毒理学分析,以期探讨(包括大型哺乳动物在内的)蓝藻毒素暴露风险。另外,利用长期的气象记录,探讨了气候变化触发这一悲剧的可能性。通过进一步分析,识别了大型动物多样性、藻毒素高浓度、干热气候以及其他有利于蓝藻生长的因素等要素在空间的热点区,对当前和未来大型动物的藻毒素暴露风险展开了情景分析。
高浓度蓝藻毒素——大型动物的杀手
人们可能会质疑,在自然生态系统中,蓝藻的毒性是否足以在短时间内杀死数百头世界上最大的陆生动物。的确,过去因蓝藻毒素所致的大规模死亡事件主要发生在鱼类、鸟类和海龟等小型动物身上[2]。然而,通过文献的综述分析发现,在非洲大陆,蓝藻毒素常常被疑为中型和大型陆生哺乳动物包括家畜(牛和羊)以及非涉水野生哺乳动物(白犀牛、蓝角马、长颈鹿、斑马和黑斑羚)大量死亡的可能原因 (图2A)(历史事件汇编见附表S1)。一个有趣的现象是,出于对捕食者的警惕,这些种类常在水域的下风处饮水,而那里也往往是蓝藻聚集地,使得它们往往较早暴露于高浓度的藻毒素(图2C)。相比之下,大象是可涉水物种,倾向于在水域中央饮水,因此,当蓝藻在局部水域发生时,接触高浓度藻毒素(蓝藻细胞内的毒素含量远高于细胞外浓度)的机会相对较小。简言之,动物行为特征可能会影响到它们对有毒蓝藻的暴露风险。
到目前为止还没有大象死亡地点藻毒素浓度的现场监测数据,但通过收集和分析众多水域的文献资料后发现,非洲大陆水体的微囊藻毒素浓度(简称MCs,是最常见和毒性最强的蓝藻毒素种类)在0.36至124,460 μg L-1之间,平均8836 μg L-1,大多数值远高于世界卫生组织WHO建议的哺乳动物和人类暂定的每日摄入耐容量指导值1.0μg L-1(详见附表S2)[3]。其中,非洲南部的情况尤其严重(发现大象大规模死亡的博茨瓦纳和津巴布韦均位于南部),MCs平均浓度达到了WHO指导值的13400倍以上,而非洲东南部有两个水体中的MCs分别为103mg L-1和124 mg L-1,几乎达到了125 mg L-1的哺乳动物急性致死剂量(见附表S2) [3]。根据大象饮水量计算了每天摄入的毒素量,也得到了类似的结果。小池塘和水坑作为野生哺乳动物重要的饮用水源地,在夏季高温作用下蒸发很快,这种浓缩效应会导致藻毒素浓度进一步升高[4]。该地区如此高的蓝藻/藻毒素浓度其实并不意外,水体中极其丰富的营养物来源(例如富含氮的污水和野生动物的粪便导致Hartbeespoort水库的氨氮水平高达273 mg L-1,是未经处理的生活污水的10倍)、静止的水体和炎热的气候条件,从多方面为蓝藻的快速繁衍提供了近乎“完美”的生长环境。
从上述分析来看,蓝藻毒素确实存在高浓度的环境风险。如果藻毒素被证实是导致这一事件的主要因素(虽然求证极为困难),这将是首例发现的由蓝藻毒素所致的大规模大象死亡事件。 通过更大范围的回顾性分析发现,在过去几十年来哺乳动物受胁的风险可能在持续上升(图2A)。根据历史报道资料可以看出,受胁对象先是从围篱家养牲畜扩展到围篱野生动物,后又扩展到自由活动但非涉水的野生动物,最后扩展到自由活动且可涉水的野生动物,即非洲象。虽然这些数据可能受到“幸存者偏差”的影响,但该现象背后的趋势不容忽视:蓝藻毒素可能威胁到越来越广泛的非洲大型动物(包括其中最大的物种——非洲象)。本次事件中大象死亡数量已经接近博茨瓦纳整整一年大象被偷猎(大象死亡的主要原因)的数量(385±54) [5],因此,蓝藻毒素对于濒危物种非洲象可能产生灾难性后果。

图 2. 不同时期非洲大陆蓝藻毒素致死的大型动物类群与数量(A);与1986年至2005年基线气候学相比,博茨瓦纳奥卡万戈三角洲塞隆加村(大象大规模死亡发生地)2000年至2020年有效积温(℃ 天)和净降水量(降水量-蒸发量,mm)的异常性分析(B);各种温度和降雨综合条件下植被、水域范围和蓝藻浓度及其对大象中毒风险的变化(C);非洲南部被确定为当前和未来藻毒素暴露风险的热点区(D);蓝藻生长对气候变暖的响应(E)。
干热天气——突发事件的导火索
在富营养化和全球变暖的推动下,全球蓝藻水华在发生频率、强度和持续时间上都有迅速增加的趋势[6, 7]。炎热的天气是蓝藻快速繁殖的主要触发因素,而野生动物和牲畜的粪便和生活污水的大量输入又进一步促进了南部非洲一些水体蓝藻的激增。对长期气候记录分析发现,在博茨瓦纳大象大规模死亡发生地区,连续多年出现了炎热和干燥情况(图2B)。
这一发现似乎不太可能是巧合。相似的气候条件也与津巴布韦的大规模大象死亡和南非的非涉水哺乳动物的大规模死亡事件有很好的吻合(详见附图S1)。炎热干燥的天气本身就促进藻毒素的产生。在干燥的天气下,哺乳动物对饮用水的需求会增加,水域又因蒸发而大幅缩小进而导致藻毒素因浓缩而浓度上升,因此进一步加剧动物中毒的风险。通过对事件发生地相关水域进行了遥感分析,发现在2020年3-7月期间,浮游藻类指数(FAI,一个蓝藻丰度的近似指标)随着水体的萎缩而持续增加(详见附图S2),意味着藻毒素暴露风险的增加。
气候变化——暴露风险的放大器
从博茨瓦纳大象大规模死亡事件中引发出了一个关键问题,即正在进行的气候变化将如何影响未来的藻毒素风险。非洲东南部存在几个“藻毒素高风险热点”,那里同时是大型动物高多样性(包括主要大象种群)热点区、藻毒素(微囊藻毒素)高浓度区以及历史上藻毒素所致动物死亡高发地(图2D)。该地区(津巴布韦)也曾首次报告了自然微囊藻毒素所致的5000例人体健康受害事件[8]。通过气候模型预测(SSP585情景),在未来五十年内,该地区变暖速度也将是整个非洲大陆最高,预计到2070年平均年气温上升将超过4℃;降水量也将低于当前条件下的降雨量(图2D,附图S3,附图S4)。令人担忧的是,随着未来气候变暖和变干,这种中毒风险将很可能增加。在更热更干燥的气候下,营养物质的浓缩将为蓝藻的生长创造更有利的条件(图2E)。在其它夏季炎热的干旱或半干旱地区,在干热的气候条件下,类似的水源地有毒蓝藻爆发与大型动物中毒事件也可能会发生。
另外,随着未来气候的变化,湖泊表层水温预计将以与气温相似甚至更高的速度上升[9]。气候变暖除了刺激毒素释放外,还会使少数毒株变种成为优势。炎热干燥的天气将促使更多动物频繁地聚集在所剩的地表水周围,这可能会通过粪便加速富营养化,进一步放大暴露风险(图2C)。最严重的情况下,气候变暖和富营养化促进蓝藻生长和毒素释放的协同效应可能会给已经因频繁的极端气候事件而遭受饥渴的脆弱野生动物种群带来致命风险[10]。
本研究的当前分析表明,随着全球变暖,未来大规模死亡事件的风险将会增加。然而,需要有更多的研究来定量预测未来蓝藻毒素对非洲象以及其他对气候变化较敏感的大型动物的致死风险。有意思的是,化石记录的更新世和始新世大象和其他大型动物史前大规模死亡,也被认为很可能是由气候变化引起的反复出现的有毒蓝藻水华所致。因此,进一步研发藻毒素与动物死亡之间的定量关联模型,可以为这些史前大规模死亡事件提供有力的解释,也可以预测野生动物种群未来面临的风险。
此外,像蓝藻毒素中毒这样的悲剧事件可能会对人类和社会产生意想不到的连锁效应。受污染水体蓝藻毒素暴露风险的增加将不可避免地损害到人类的健康和生活,尤其当人们的生活依赖于那些牲畜和野生动物,或者把受污染的水作为饮用水或者灌溉用途。蓝藻毒素在博茨瓦纳大象群体性死亡事件中的作用仍需进一步研究,提醒我们亟需对野生动物和人类藻毒素暴露风险进行更全面和系统的定量评估,以便能够有效地实施面对气候变化下的预防行动,削弱蓝藻毒素对本已脆弱的非洲社会-生态系统的胁迫。
结语
近期发生在非洲博茨瓦纳的非洲大象大规模群体死亡事件引起了国内外的广泛关注。蓝藻毒素被认为是可能的原因,但一直缺乏有效的科学证据。本文通过综合多源数据进行分析,发现在过去的几十年里,南部非洲大范围的超富营养化水域是该区域的高风险区,该地区的野生动物频繁地暴露于高浓度藻毒素中,而该区域也是非洲哺乳动物物种多样性的热点分布区。近年极端炎热和干燥的气候很可能是触发这次大规模死亡事件的重要因素,古生物学证据也表面,类似的事件或许也曾发生在更新世和始新世时期。根据全球气候模型预测,这种极端气候条件在未来半个世纪内将在南部非洲变得更加频繁,类似的事件可能会再次发生,从而对非洲大型动物产生威胁,特别是可能增加濒危灭绝的面临风险。此外,气候变化对蓝藻毒素致毒性的“增强效应”还可能会对人类社会产生意想不到的级联效应。非洲大象是世界上现存体型最大的陆生哺乳动物,近期大规模死亡的悲剧事件可能是环境变化的重要早期预警信号,提醒我们亟需关注非洲南部未来的环境风险。建议今后应当对蓝藻毒素的风险开展系统定量评估,并果断采取行动以减轻风险,以确保该区域内大型动物的健康和人类社会的可持续发展。
参考文献
1. Azeem, S., Bengis, R., Aarde, R.V., and Bastos, A.D.S. (2020). Mass Die-Off of African Elephants in Botswana: Pathogen, Poison or a Perfect Storm? sawr.1 50, 149–156.
2. Svirčev, Z., Lalić, D., Bojadžija Savić, G., Tokodi, N., Drobac Backović, D., Chen, L., Meriluoto, J., and Codd, G.A. (2019). Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch Toxicol 93, 2429–2481.
3. World Health Organization ed. (2011). Guidelines for drinking-water quality 4th ed. (World Health Organization).
4. Jeppesen, E., Beklioğlu, M., Özkan, K., and Akyürek, Z. (2020). Salinization Increase due to Climate Change Will Have Substantial Negative Effects on Inland Waters: A Call for Multifaceted Research at the Local and Global Scale. The Innovation 1, 100030.
5. Schlossberg, S., Chase, M.J., and Sutcliffe, R. (2019). Evidence of a Growing Elephant Poaching Problem in Botswana. Current Biology 29, 2222-2228.e4.
6. Paerl, H.W., and Paul, V.J. (2012). Climate change: Links to global expansion of harmful cyanobacteria. Water Research 46, 1349–1363.
7. Huisman, J., Codd, G.A., Paerl, H.W., Ibelings, B.W., Verspagen, J.M.H., and Visser, P.M. (2018). Cyanobacterial blooms. Nature Reviews Microbiology 16, 471–483.
8. Zilberg, B. (1966). Gastroenteritis in Salisbury. European children--a five-year study. Cent Afr J Med 12, 164–168.
9. Woolway, R.I., Kraemer, B.M., Lenters, J.D., Merchant, C.J., O’Reilly, C.M., and Sharma, S. (2020). Global lake responses to climate change. Nature Reviews Earth & Environment 1, 388–403.
10. Nangombe, S., Zhou, T., Zhang, W., Wu, B., Hu, S., Zou, L., and Li, D. (2018). Record-breaking climate extremes in Africa under stabilized 1.5 °C and 2 °C global warming scenarios. Nature Climate Change 8, 375–380.
本研究发表在Cell旗下的综合性期刊——The Innovation(创新)上:Wang, H., Xu, C., Liu, Y., Jeppesen, E., Svenning, J.-C., Wu, J., Zhang, W., Zhou, T., Wang, P., Nangombe, S., Ma, J., Duan, H., Fang, J., Xie, P., From unusual suspect to serial killer: Cyanotoxins boosted by climate change may jeopardize African megafauna, The Innovation (2021), doi: https://doi.org/10.1016/j.xinn.2021.100092.

原文期刊下载地址:https://www.cell.com/the-innovation/fulltext/S2666-6758(21)00017-5
本论文的第一作者为王海军(云南大学)、徐驰(南京大学)和刘莹(云南大学),通讯作者为谢平(中国科学院水生生物研究所、云南大学),参加研究的还有丹麦奥胡斯大学的Erik Jeppesen教授和Jens-Christian Svenning教授、美国亚利桑那州立大学的邬建国教授、中科院大气物理研究所的周天军研究员和张文霞博士、以及云南大学的王普泽、津巴布韦哈拉雷气象服务部的Shingirai Nangombe博士、中科院南京地理与湖泊研究所的段洪涛研究员和马金戈等。方精云院士提供了指导意见。本研究得到中国科学院战略性先导科技专项(B类)“大尺度区域生物多样性格局与生命策略”(XDB31000000)等的资助。
附件
气候数据处理与分析方法Methods for climatic data processing and analyses
To investigate the climatic conditions associated with cyanobacterial blooms, temperature and precipitation records are examined. To account for potential observational uncertainty, multiple observational datasets are used. Global monthly temperature datasets used include: (1) GISS (NASA Goddard Institute for Space Studies; 2°X2°) and (2) BEST (Berkeley Earth Surface Temperature); 1°X1°.1,2 Global monthly precipitation observation is from the GPCP (Global Precipitation Climatology Project); 2.5°X2.5°.3 Monthly precipitation and evaporation data from the ERA5 reanalysis (0.25°X0.25°) are also used.4
Accumulated temperature for cyanobacteria is defined as follows5,6:
K=N(T-C)
where K is the accumulated temperature, C is the temperature threshold above which cyanobacteria recruit (C=9°C), N is the length of the cyanobacterial growing period (i.e. when T>C), and T is the mean temperature over the growing period. In this study, annual accumulated temperature is computed from preceding June to May (in °C days).
Likewise, the annual accumulated net precipitation is calculated from preceding June to May (in mm), which represents the availability of surface water. Here net precipitation (i.e. precipitation minus evaporation) is used so that the enhanced evaporation accompanying hot conditions is taken into account.
The temperature and precpitation records from different datasets consistently reveal the anomalous hot and dry conditions during 2019-2020 in Botswana and Zimbabwe (figure not shown). Thus, only the results from the GISS and ERA5 datasets are provided in the manuscript for brevity.
遥感数据处理与分析方法Methods for processing and analyses of remote sensing data
NDWI (Normalized Difference Water Index) was calculated according to REF7:
NDWI = (GREEN-NIR)/(GREEN+NIR)
where GREEN and NIR represent green band and near infrared band of MSI and OLI. And pixels with NDWI greater than 0 are defined as constant water body.
FAI (Floating Algae Index) was calculated according to REF8:
FAI = RED - NIR'
NIR' = RED + [(SWIR-RED)]*[(λ(NIR)-λ(RED)/(λ(SWIR)-λ(RED))]
where RED, SWIR and NIR represent red band, near infrared band and shortwave infrared band of MSI and OLI. λ(RED), λ(NIR) and λ(SWIR) represent the center wavelength of red band, near infrared band and shortwave infrared band of MSI and OLI.
图2D中的方法Methods for Figure 2 D
Response curve of growth rate of cyanobacteria to temperature was modified from REF9; Data on elephant density were collected from REF,10 and data on mammal species richness were compiled based on their distribution range maps from the African Mammal Databank.
Text S1 非洲象大规模死亡的主要可能原因
可排除的原因
1偷猎象牙:身体完好无损。
2农民的破坏(9种常用杀虫剂):实验室测试。
3饥饿或脱水:发现地有丰富的植被,70%靠近水坑和池塘。
4炭疽中毒:不同于其对老年人和年轻人的选择性作用,本次事件各个年龄段死亡个体都有。
可能的原因
1藻毒素
-实验室检测(博茨瓦纳野生动物和国家公园部门发布)(1)。
-在有大量蓝藻的水坑或池塘里/附近发现大象尸体。
-大出血
-在突然死亡(有时是脸部先着地)之前,会出现转圈和头晕,这是典型的神经毒素中毒状(2,3)。
2出血性败血症(一种啮齿动物传播的病毒感染)
-神经症状
-大出血
相关媒体报道:
1. Weston, P. 2020 Botswana says it has solved the mystery of mass elephant die-off. https://www.theguardian.com/environment/2020/sep/21/Botswana-says-it-has-solved-mystery-of-mass-elephant-die-off-age-of-extinction-aoe
2. Boult, V. 2020. Hundreds of elephants are mysteriously dying in Botswana – a conservationist explains what we know. July 3, 2020. https://theconversation.com/hundreds-of-elephants-are-mysteriously-dying-in-Botswana-a-conservationist-explains-what-we-know-142004
3. Maron, D. F. 2020. What’s killing Botswana’s elephants? Here are the top theories. https://www.nationalgeographic.com/animals/2020/07/botswana-elephant-death-mystery/
奥卡万戈三角洲死亡大象图片网页链接:
https://www.reuters.com/article/us-botswana-elephants-idUSKCN24N2JK
Text S2 蓝藻,蓝藻毒素,富营养化概念介绍
Cyanobacteria
Cyanobacteria, also known as blue-green algae, are oxygen-producing bacteria and the Earth’s oldest (~ 3.5 bya) organisms, with a major shaping effect on the origin of our modern-day biosphere through oxidization of the Earth’s atmosphere.11 Cyanobacteria occur throughout the world, especially in calm water rich in nutrients. Cyanobacteria can form dense blooms under nutrient-rich conditions, resulting in increased turbidity, smothering of submersed macrophytes, and oxygen depletion followed by massive deaths of fish and sensitive macroinvertebrates. Cyanobacteria can interfere with the recreational function of waters and their use for drinking water purposes by their generation of obnoxious taste and odor compounds. Cyanobacteria can also produce a variety of toxins, called cyanotoxins, which are detrimental and even lethal to birds, mammals and humans.12 Cyanobacteria are particularly abundant in nutrient-rich and hot environments such as southeastern Africa.13,14
Cyanotoxins
Cyanotoxins can be classified into various types according to their modes of action and target cells and organs: hepatotoxins (the most frequently encountered), neurotoxins, dermatotoxins, and cytotoxins.12,15 The symptoms of cyanotoxin poisoning include skin irritation, stomach aches, vomiting, nausea, diarrhea, fever, sore throat, and headache. The largest group of cyanotoxins is the cyclic heptapeptide hepatotoxins called microcystins (MCs), in which MC-LR is the best known and most toxic structural variant.16 Microcystins predominantly cause liver injury.17,18 An acute, lethal dose exposure can result in almost total breakdown of the tissue architecture and blood accumulation in the liver, where blood loss from the circulation causes death to the animal. Besides hepatotoxicity and tumor promotion, MCs may also induce neurotoxicity, reproductive toxicity, genotoxicity, and potential carcinogenicity.19–22 Despite their aquatic origin, most cyanotoxins tend to be more hazardous to terrestrial mammals than to aquatic biota.23 Furthermore, the proportion of toxic species or strain and release of toxins tend to increase with global warming.24,25
Eutrophication
Eutrophication is an enrichment process in aquatic ecosystems created by excessive loading of nutrients (e.g. nitrogen and phosphorus), which promotes massive growth of phytoplankton, including cyanobacteria. A shift from a macrophyte-dominated state to a phytoplankton-dominated state will happen when nutrient loading surpasses a critical level.26,27 Human activities are a common cause of eutrophication, with both point-source and non-point-source discharges accelerating the rate at which nutrients enter ecosystems. In parts of Africa, wild mammals such as hippos contribute significantly to the nutrient loading of freshwater ecosystems by moving nutrients from terrestrial to aquatic ecosystems, and by foraging on land, whereas much non-feeding time is spent in water.28 Therefore, savanna waters can become highly eutrophic also without the influence of human activities.
Text S3 非洲象每日微囊藻毒素可耐受摄入量和半致死剂量的推导
世界卫生组织 (WHO)关于藻毒素安全暴露的暂行指南
为了确定藻毒素的安全水平,人们对其毒性进行了广泛的实验研究。微囊藻毒素Microcystin-LR (MC-LR)是最常见、毒性最强的一种形式,也是研究最多的一种形式。由于直接评估人类接触藻毒素风险不可行,已有研究主要是通过动物暴露实验来解决这个问题。其中,对纯MC-LR(灌胃)的小鼠进行为期13周的研究,被认为最适合用于推导MC-LR安全暴露的指导值29。根据肝脏组织病理学和血清酶水平的变化,实验确定NOAEL(无观察到的不良反应水平)为40μg kg-1·bw day-1。进一步通过总不确定系数1000,建立了0.04μg kg-1bw day-1的每日耐受摄入量(Tolerable Daily Intake, TDI)作为安全水平的暂定指标23。采用平均体重60 kg和平均每天2升水摄入量的成人指标,确定1.0μg L-1为暂定安全暴露指标。对于急性暴露,采用的是基于对小鼠的灌胃试验,确定了5mg kg-1 bw的半数致死剂量(LD50) 29
非洲象的TDI和LD50
根据非洲象(Loxodonta africana)的平均体重(5000千克,每天200升水摄入量)30,我们推导出其对应的TDI和LD50如下:
TDIL.africana
= TDIWHO2011*bwL.africana/Water ingestion
= 0.04 μg kg-1 bw day-1*5000 kg/200 L
=1.0 μg L-1 d-1
LD50L.africana
= LD50WHO2011*bwL.africana/Water ingestion
=5 mg kg-1 *5000 kg/200 L
=125 mg L
附表S1 蓝藻毒素所致的动物死亡历史事件
Year | Country | Location | Cyanobacteria species | Toxin | Impacts reported | Reference number | |
Death | Illness | ||||||
1913- 1943 | South Africa | Free State and Southeast Transvaal | Microcystis aeruginosa | microcystin | thousands of livestock (horses, sheep, cattle and rabbits) | 31 | |
1942 | South Africa | Vaal Dam | Microcystis sp. | microcystin | thousands of cattle and sheep | 32,33 | |
1973- 1974 | South Africa | Hartbeespoort Dam | M. aeruginosa | microcystin | cattle | 34 | |
1979 | South Africa | Klipvoor Dam | M. aeruginosa | microcystin | 3 white rhinoceroses | 35 | |
1980 | South Africa | Vaal Dam | M. aeruginosa | microcystin | cattle | 31 | |
1984 | South Africa | William Pretorius Game Reserve | M. aeruginosa | microcystin | several black wildebeests | 35 | |
1987 | South Africa | Eastern Transvaal | M. aeruginosa | microcystin | 47 cattle | 36 | |
1989 | South Africa | Reservoir Bloemhof Dam | M. aeruginosa | microcystin | 7 giraffes | 37 | |
1989 | South Africa | Reservoir Klipdrif Dam | M. aeruginosa | microcystin | livestock | livestock | 38 |
1993 | South Africa | Malmesbury | Nodularia spumigena | nodularin | 3 cattle | 10 cattle | 39 |
1994 | South Africa | Lake Zeekoevlei | N. spumigena, M. aeruginosa | nodularin, microcystin | dog (bull terrier bitch) | 39,40 | |
1994 | South Africa | Malmesbury | N. spumigena | nodularin | 34 cattle | 39 | |
1994 | South Africa | Paarl | M. aeruginosa | microcystin | 11 sheep | 30 sheep | 39 |
1996 | South Africa | Kareedouw | Oscillatoria sp. | microcystin | 290 dairy cows | 70 stock | 41 |
1998 | South Africa | Erfenis Dam | Anabaena spp. | cyanotoxin | livestock | 31,42 | |
2000 | South Africa | Orange River system downstream of the confluence with the Harts River | Cylindrospermopsis raciborski Anabaena sp. Oscillatoria sp. | not available | fish | 41,43 | |
2001 | Kenya | Lake Bogoria | Phormidium terebriformis O. willei, Spirulina subsalsa, Synechococcus bigranulatus | microcystin, anatoxin | mass lesser flamingos | 44 | |
2005 | South Africa | Nhlanganzwane Dam | M. aeruginosa | microcystin | 7 white rhinoceroses, 2 lions, 2 cheetahs, 9 zebras, 23 wildebeest, 1 hippopotamus, 1 giraffe, 5 buffalos, 1 warthog, 1 Kudu | 45 | |
2007 | South Africa | Nhlanganzwane Dama | M. aeruginosa | microcystin | 15 white rhinoceroses, 10 zebras, 10 blue wildebeests | wild animals | 43,45,46 |
2007 | South Africa | Sunset Dam | M. aeruginosa | microcystin | 1 white rhinoceros, 6 impalas | 46 | |
2008 | South Africa | Lake Loskop Dam | M. aeruginosa M. flos-aquae | microcystin | fish | 47 | |
2008 | South Africa | Shilolweni Dam | Microcystis sp. | microcystin | 70 wild animals (zebras, wildebeest, impalas, white rhinoceroses) | 45 | |
2008 | Botswana | Tuli block | Oscillatoria sp. | microcystin | 70 bushbucks and impalas | 43 | |
2010 | South Africa | Shilolweni Dam | Microcystis sp. | microcystin | 17 zebras, 7 wildebeests, 3 white rhinoceroses | 45 | |
附表S2. 已发表的非洲水体微囊藻毒素浓度,以此推导的非洲象可耐受藻毒素日摄入量
Code | Zone | Country | Waters | MCs, μg/L | Daily intake by African elephant per body weight (MCs*200L/5000 kg), μg kg-1bw | Reference number |
1 | Northern | Algeria | Ain Zada Dam | 69.3 | 2.8 | 48 |
2 | Northern | Algeria | Cheffia Dam | 28.9 | 1.2 | 49 |
3 | Northern | Algeria | Lake Oubcim | 46.2 | 1.8 | 14 |
4 | Northern | Egypt | Nile River | 7.1 | 0.3 | 50 |
5 | Northern | Morocco | Lake Lalla Takerkoust | 19.9 | 0.8 | 14 |
6 | Western | Ghana | Kpong Dam | 0.03 | 0.001 | 51 |
7 | Western | Ghana | Weija Dam | 3.21 | 0.1 | 51 |
8 | Western | Nigeria | Zaria aquaculture ponds | 6.34 | 0.3 | 14 |
9 | Eastern | Ethiopia | Lake Chamo | 28.9 | 1.2 | 14 |
10 | Eastern | Ethiopia | Lake Chamo | 6.1 | 0.2 | 52 |
11 | Eastern | Ethiopia | Lake Koka | 3.9 | 0.2 | 14 |
12 | Eastern | Ethiopia | Lake Koka | 51 | 2.0 | 52 |
13 | Eastern | Ethiopia | Lake Langano | 1.3 | 0.1 | 52 |
14 | Eastern | Ethiopia | Lake Ziway | 1.3 | 0.1 | 52 |
15 | Eastern | Kenya | Lake Simbi | 29163 | 1167 | 53 |
16 | Eastern | Kenya | Lake Sonachi | 64.8 | 2.6 | 53 |
17 | Eastern | Kenya | Lake Victoria, Nyanza Gulf | 0.4 | 0.02 | 54 |
18 | Eastern | Kenya | Nakuru oxidation ponds | 1.72 | 0.1 | 14 |
19 | Eastern | Kenya | Nyanza Gulf | 82.5 | 3.3 | 14 |
20 | Eastern | Kenya, Tanzania, Uganda | Lake Victoria | 1 | 0.04 | 55 |
21 | Eastern | Uganda | Lake Saka | 31783 | 1271 | 14 |
22 | Eastern | Uganda | Murchison Bay | 3.1 | 0.1 | 14 |
23 | Southern | Mozambique | Maputo and Gaza provinces | 7.31 | 0.3 | 13 |
24 | Southern | South Africa | Hartbeespoort Dam | 580 | 23.2 | 56 |
25 | Southern | South Africa | Hartbeespoort Dam | 1965 | 78.6 | 41 |
26 | Southern | South Africa | Hartbeespoort Dam | 1861 | 74.4 | 41 |
27 | Southern | South Africa | Hartbeespoort Dam | 1698 | 67.9 | 41 |
28 | Southern | South Africa | Hartbeespoort Dam | 414 | 16.6 | 41 |
29 | Southern | South Africa | Hartbeespoort Dam | 1538 | 61.5 | 41 |
30 | Southern | South Africa | Hartbeespoort Dam | 1314 | 52.6 | 41 |
31 | Southern | South Africa | Hartbeespoort Dam | 474 | 19.0 | 41 |
32 | Southern | South Africa | Hartbeespoort Dam | 4.65 | 0.2 | 57 |
33 | Southern | South Africa | Hartbeespoort Dam | 12300 | 492 | 58 |
34 | Southern | South Africa | Hartbeespoort Dam | 44878 | 1795 | 59 |
35 | Southern | South Africa | Klipvoor Dam | 22330 | 893 | 41 |
36 | Southern | South Africa | Klipvoor Dam | 21100 | 844 | 41 |
37 | Southern | South Africa | Loskop Dam | 0.36 | 0.01 | 14 |
38 | Southern | South Africa | Loskop Dam | 0.09 | 0.004 | 14 |
39 | Southern | South Africa | Loskop Dam | 3.173 | 0.1 | 60 |
40 | Southern | South Africa | Makhohlola Dam | 0.2 | 0.01 | 46 |
41 | Southern | South Africa | Makhohlolo Dam | 3.38 | 0.1 | 14 |
42 | Southern | South Africa | Mpanamana Dam | 1 | 0.04 | 46 |
43 | Southern | South Africa | Nhlanganzwane Dam | 24951 | 998 | 14 |
44 | Southern | South Africa | Nhlanganzwane Dam | 103160 | 4126 | 46 |
45 | Southern | South Africa | Nhlanganzwane Dam | 23720 | 949 | 47 |
46 | Southern | South Africa | Rietvlei Dam | 198 | 7.9 | 41 |
47 | Southern | South Africa | Rietvlei Dam | 184 | 7.4 | 41 |
48 | Southern | South Africa | Roodeplaat Dam | 981 | 39.2 | 41 |
49 | Southern | South Africa | Sunset Dam | 124460 | 4978 | 46 |
50 | Southern | South Africa | Sunset Dam | 1110 | 44.4 | 46 |
51 | Southern | Zimbabwe | Lake Chivero | 19.86 | 0.8 | 61 |
Average | Northern Africa | 34.28 | ||||
Western Africa | 3.19 | |||||
Eastern Africa | 4371 | |||||
Southern Africa | 13423 | |||||
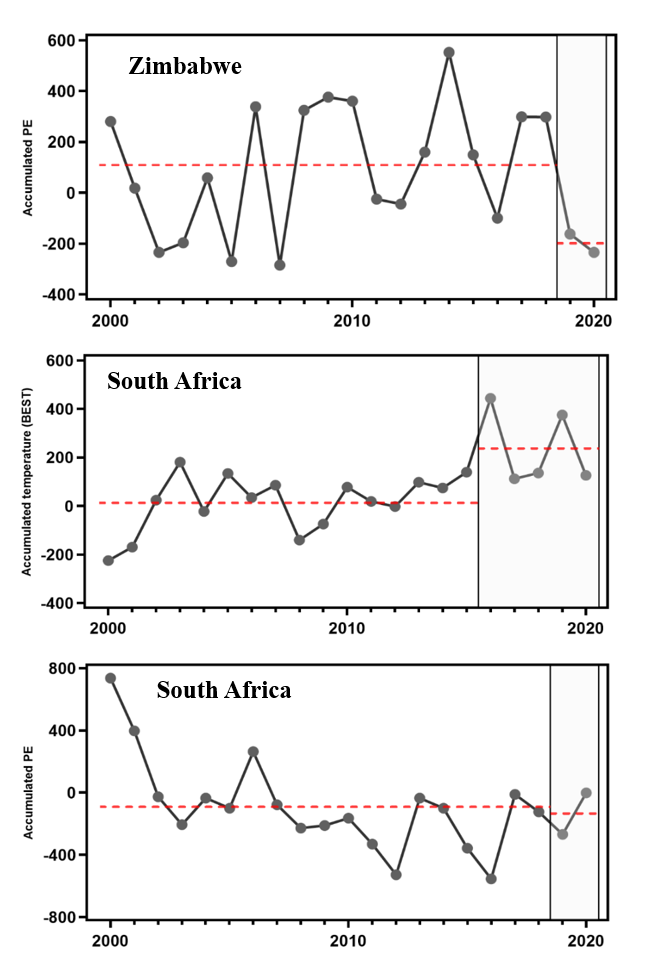
附图S1. 津巴布韦和南非2000年至2020年期间(相对于1986年至2005年基线气候)积温(从 6月至次年5月)(℃ days)和净降水量(降水量减去蒸发量,mm)的异常性分析

附图 S2. 大象大规模死亡发生地区(博茨瓦纳奥卡万戈三角洲塞隆加西北部)遥感分析区域(A),大象死亡期间(2020年3月至7月)图中方框内水域面积和浮游藻类指数(floating algae index,FAI)的变化。使用NDWI结合Sentinel-2/MSI图像提取大象死亡期的恒定水域面积,同时获得Sentinel-2/MSI和Landsat-8/OLI的水域面积和浮游藻类指数的变化。



附图S3. 2020年与非洲象死亡相关的极端干热气候条件的空间分布。A. 2015年-2020年的年有效积温(从6月到次年的5月;℃ 天)与1986年-2005年的基线气候学相比较。数据来源于 BEST data。图中的两颗星分布代表博茨瓦纳 (18.84S, 22.90E) 和津巴布韦 (18.51S, 25.95E)大象大规模死亡地。B.基于 GISS data的年有效积温,其余与A一致。C. 据ERA5 data得出2019-2020年间年累计净降水量(降水量减去蒸发量,mm) ,其余与A一样。


附图S4. 2070年前后年平均气温和降水量的变化,南部非洲被确定为气候变暖和干燥的热点地区。从9个CMIP6全球气候模式(BCC-CSM2-MR, CNRM-CM6-1, CNRM-ESM2-1, CanESM5, GFDL-ESM4, IPSL-CM6A-LR, MIROC-ES2L, MIROC6, MRI-ESM2-0) 获得2061-2080年期间预测的年平均气温和降水数据,用于4条共同的社会经济路径(SSP126, 245, 370和585。WorldClim v2.1数据用作当前基线。空间分辨率为~10公里的气候数据从WorldClim网站下载。
附件参考文献
1. Hansen, J., Ruedy, R., Sato, M., and Lo, K. (2010). Current GISS global surface temperature analysis. NASA Goddard Inst. Space Stud. N. Y., 1–34.
2. Rohde, R., Muller, R.A., Jacobsen, R., Muller, E., Perlmutter, S., Rosenfeld, A., Wurtele, J., Groom, D., and Wickham, C. (2013). A New Estimate of the Average Earth Surface Land Temperature Spanning 1753 to 2011, Geoinfor Geostat: An Overview 1: 1. of 7, 2.
3. Adler, R.F., Huffman, G.J., Chang, A., Ferraro, R., Xie, P.-P., Janowiak, J., Rudolf, B., Schneider, U., Curtis, S., Bolvin, D., et al. (2003). The Version-2 Global Precipitation Climatology Project (GPCP) Monthly Precipitation Analysis (1979–Present). J. Hydrometeorol. 4, 1147–1167.
4. Hersbach, H., Bell, B., Berrisford, P., Hirahara, S., Horányi, A., Muñoz‐Sabater, J., Nicolas, J., Peubey, C., Radu, R., Schepers, D., et al. (2020). The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 146, 1999–2049.
5. Cao, H.-S., Kong, F.-X., Tan, J.-K., Zhang, X.-F., Tao, Y., and Yang, Z. (2005). Recruitment of Total Phytoplankton, Chlorophytes and Cyanobacteria from Lake Sediments Recorded by Photosynthetic Pigments in a Large, Shallow Lake (Lake Taihu, China). Int. Rev. Hydrobiol. 90, 347–357.
6. Cao, H.-S., Tao, Y., Kong, F.-X., and Yang, Z. (2008). Relationship between Temperature and Cyanobacterial Recruitment from Sediments in Laboratory and Field Studies. J. Freshw. Ecol. 23, 405–412.
7. McFEETERS, S.K. (1996). The use of the Normalized Difference Water Index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 17, 1425–1432.
8. Hu, C. (2009). A novel ocean color index to detect floating algae in the global oceans. Remote Sens. Environ. 113, 2118–2129.
9. Paerl, H.W., and Otten, T.G. (2013). Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 65, 995–1010.
10. Chase, M.J., Schlossberg, S., Griffin, C.R., Bouché, P.J.C., Djene, S.W., Elkan, P.W., Ferreira, S., Grossman, F., Kohi, E.M., Landen, K., et al. (2016). Continent-wide survey reveals massive decline in African savannah elephants. PeerJ 4, e2354.
11. Huisman, J., Codd, G.A., Paerl, H.W., Ibelings, B.W., Verspagen, J.M.H., and Visser, P.M. (2018). Cyanobacterial blooms. Nat. Rev. Microbiol. 16, 471–483.
12. Xie, P. (2006). Microcystins in aquatic animals with potential risk to human health (Scientific Press, Beijing).
13. Tamele, I.J., and Vasconcelos, V. (2020). Microcystin Incidence in the Drinking Water of Mozambique: Challenges for Public Health Protection. Toxins 12, 368.
14. Ndlela, L.L., Oberholster, P.J., Van Wyk, J.H., and Cheng, P.H. (2016). An overview of cyanobacterial bloom occurrences and research in Africa over the last decade. Harmful Algae 60, 11–26.
15. Falconer, I.R. (1999). An Overview of problems caused by toxic blue–green algae (cyanobacteria) in drinking and recreational water. Environ. Toxicol. 14, 5–12.
16. Qi, Y., Rosso, L., Sedan, D., Giannuzzi, L., Andrinolo, D., and Volmer, D.A. (2015). Seven new microcystin variants discovered from a native Microcystis aeruginosa strain – unambiguous assignment of product ions by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 29, 220–224.
17. Chen, J., Xie, P., Li, L., and Xu, J. (2009). First Identification of the Hepatotoxic Microcystins in the Serum of a Chronically Exposed Human Population Together with Indication of Hepatocellular Damage. Toxicol. Sci. 108, 81–89.
18. Falconer, I.R. (1991). Tumor promotion and liver injury caused by oral consumption of cyanobacteria. Environ. Toxicol. Water Qual. 6, 177–184.
19. Liu, Y., Xie, P., Qiu, T., Li, H.-Y., Li, G.-Y., Hao, L., and Xiong, Q. (2010). Microcystin extracts induce ultrastructural damage and biochemical disturbance in male rabbit testis. Environ. Toxicol. 25, 9–17.
20. Zhao, Y., Xie, P., and Fan, H. (2012). Genomic Profiling of MicroRNAs and Proteomics Reveals an Early Molecular Alteration Associated with Tumorigenesis Induced by MC-LR in Mice. Environ. Sci. Technol. 46, 34–41.
21. Chen, L., Chen, J., Zhang, X., and Xie, P. (2016). A review of reproductive toxicity of microcystins. J. Hazard. Mater. 301, 381–399.
22. Hu, Y., Chen, J., Fan, H., Xie, P., and He, J. (2016). A review of neurotoxicity of microcystins. Environ. Sci. Pollut. Res. 23, 7211–7219.
23. World Health Organization ed. (2011). Guidelines for drinking-water quality 4th ed. (World Health Organization).
24. Walls, J.T., Wyatt, K.H., Doll, J.C., Rubenstein, E.M., and Rober, A.R. (2018). Hot and toxic: Temperature regulates microcystin release from cyanobacteria. Sci. Total Environ. 610–611, 786–795.
25. Mantzouki, E., Lürling, M., Fastner, J., De Senerpont Domis, L., Wilk-Woźniak, E., Koreivienė, J., Seelen, L., Teurlincx, S., Verstijnen, Y., Krztoń, W., et al. (2018). Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins 10, 156.
26. Scheffer, M., and Jeppesen, E. (2007). Regime Shifts in Shallow Lakes. Ecosystems 10, 1–3.
27. Wang, H.-J., Wang, H.-Z., Liang, X.-M., and Wu, S.-K. (2014). Total phosphorus thresholds for regime shifts are nearly equal in subtropical and temperate shallow lakes with moderate depths and areas. Freshw. Biol. 59, 1659–1671.
28. Moss, B. (2015). Mammals, freshwater reference states, and the mitigation of climate change. Freshw. Biol. 60, 1964–1976.
29. Fawell, J.K., James, C.P., and James, H.A. (1994). Toxins from blue-green algae: toxicological assessment of microcystin-LR and a method for its determination in water. Water Res. Cent. Medmenham UK, 1–46.
30. Sach, F., Dierenfeld, E.S., Langley-Evans, S.C., Hamilton, E., Murray Lark, R., Yon, L., and Watts, M.J. (2020). Potential bio-indicators for assessment of mineral status in elephants. Sci. Rep. 10, 8032.
31. Oberholster, P.J., Botha, A.-M., and Cloete, T.E. (2005). An overview of toxic freshwater cyanobacteria in South Africa with special reference to risk, impact and detection by molecular marker tools. Biokemistri 17, 57–71.
32. Stephens, E.L. (1949). Microcystis Toxica Sp. Nov.: A Poisonous Alga from the Transvaal and Orange Free State. Trans. R. Soc. South Afr. 32, 105–112.
33. Steyn, D.G. (1945). Poisoning of animals by algae (scum or waterbloom) in dams and pans. Trans Dept Agr Union Afr, 1–9.
34. Toerien, D.F., Scott, W.E., and Pitout, M.J. (1976). Microcystis toxins: isolation, identification, implications. Water Sa 2, 160–162.
35. Soll, M.D. & W., M.C. (1985). Mortality of a white rhinoceros (Ceratotherium simum) suspected to be associated with the blue-green alga Microcystis aeruginosa. J. S. Afr. Vet. Assoc. 56, 49–51.
36. Scott, W.E. (1991). Occurrence and Significance of Toxic Cyanobacteria in Southern Africa. Water Sci. Technol. 23, 175–180.
37. Theron, C.P. (1990). Kameelperdvrektes en die teenwoordigheid van Microcystis in Bloemhof Dam. DWAF-HRI Rep. N 4.
38. Theron, C.P. (1990). ‘n Ondersoek na die voorkoms van Microcystis in Klipdrif Dam. Inst. Water Qual. Stud. Rep. N4841Department Water. Recite from: Oberholster, P.J., Botha, A.-M., and Cloete, T.E. (2005). An overview of toxic freshwater cyanobacteria in South Africa with special reference to risk, impact and detection by molecular marker tools. Biokemistri 17, 57–71.
39. Van Halderen, A., Harding, W.R., Wessels, J.C., Schneider, D.J., Heine, E.W., Van der Merwe, J., and Fourie, J.M. (1995). Cyanobacterial (blue-green algae) poisoning of livestock in the western Cape Province of South Africa. J. S. Afr. Vet. Assoc. 66, 260–264.
40. G. A. Harding, W.R.* (1995). Death of a dog attributed to the cyanobacterial (blue-green algal) hepatotoxin nodularin in South Africa. J. S. Afr. Vet. Assoc. 66, 256–259.
41. Harding, W.R., and Paxton, B.R. (2001). Cyanobacteria in South Africa: a review (Water Research Commission Pretoria).
42. Van Ginkel, C.E., and Hohls, B.C. (1999). Toxic algae in Erfenis and Allemanskraal dams. Occasional report by the Institute for Quality Studies, Department of Water Affairs and Forestry (N).
43. Oberholster, P.J., Botha, A.-M., and Myburgh, J.G. (2009). Linking climate change and progressive eutrophication to incidents of clustered animal mortalities in different geographical regions of South Africa. Afr. J. Biotechnol. 8.
44. Krienitz, L., Ballot, A., Kotut, K., Wiegand, C., Pütz, S., Metcalf, J.S., Codd, G.A., and Stephan, P. (2003). Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 43, 141–148.
45. Bengis, R., Govender, D., Lane, E., Myburgh, J., Oberholster, P., Buss, P., Prozesky, L., and Keet, D. (2016). Eco-epidemiological and pathological features of wildlife mortality events related to cyanobacterial bio-intoxication in the Kruger National Park, South Africa. J. S. Afr. Vet. Assoc. 87, 1–9.
46. Masango, M.G., Myburgh, J.G., Labuschagne, L., Govender, D., Bengis, R.G., and Naicker, D. (2010). Assessment of Microcystis bloom toxicity associated with wildlife mortality in the Kruger national park, South Africa. J. Wildl. Dis. 46, 95–102.
47. Oberholster, P.J., Myburgh, J.G., Govender, D., Bengis, R., and Botha, A.-M. (2009). Identification of toxigenic Microcystis strains after incidents of wild animal mortalities in the Kruger National Park, South Africa. Ecotoxicol. Environ. Saf. 72, 1177–1182.
48. Saoudi, A., Brient, L., Boucetta, S., Ouzrout, R., Bormans, M., and Bensouilah, M. (2017). Management of toxic cyanobacteria for drinking water production of Ain Zada Dam. Environ. Monit. Assess. 189, 361.
49. Nasri, H., Bouaïcha, N., and Harche, M.K. (2007). A new morphospecies of Microcystis sp. forming bloom in the Cheffia dam (Algeria): Seasonal variation of microcystin concentrations in raw water and their removal in a full-scale treatment plant. Environ. Toxicol. 22, 347–356.
50. Mohamed, Z.A., and Hussein, A.A. (2006). Depuration of microcystins in tilapia fish exposed to natural populations of toxic cyanobacteria: A laboratory study. Ecotoxicol. Environ. Saf. 63, 424–429.
51. Addico, G., Hardege, J., Komarek, J., Babica, P., and Graft-Johnson, K. de (2006). Cyanobacteria species identified in the Weija and Kpong reservoirs, Ghana, and their implications for drinking water quality with respect to microcystin. Afr. J. Mar. Sci. 28, 451–456.
52. Willén, E., Ahlgren, G., Tilahun, G., Spoof, L., Neffling, M.-R., and Meriluoto, J. (2011). Cyanotoxin production in seven Ethiopian Rift Valley lakes. Inland Waters 1, 81–91.
53. Ballot, A., Krienitz, L., Kotut, K., Wiegand, C., and Pflugmacher, S. (2005). Cyanobacteria and cyanobacterial toxins in the alkaline crater lakes Sonachi and Simbi, Kenya. Harmful Algae 4, 139–150.
54. Simiyu, B.M., Oduor, S.O., Rohrlack, T., Sitoki, L., and Kurmayer, R. (2018). Microcystin Content in Phytoplankton and in Small Fish from Eutrophic Nyanza Gulf, Lake Victoria, Kenya. Toxins 10, 275.
55. Olokotum, M., Mitroi, V., Troussellier, M., Semyalo, R., Bernard, C., Montuelle, B., Okello, W., Quiblier, C., and Humbert, J.-F. (2020). A review of the socioecological causes and consequences of cyanobacterial blooms in Lake Victoria. Harmful Algae 96, 101829.
56. Pindihama, G.K., and Gitari, M.W. (2020). Cyanobacterial toxins: an emerging threat in South African irrigation water. Water Environ. J. 34, 506–516.
57. Mokoena, M.M., Mukhola, M.S., and Okonkwo, O.J. (2016). Hazard Assessment of Microcystins from the Household’s Drinking Water. Appl. Ecol. Environ. Res. 14, 695–710.
58. Oberholster, P.J., Cloete, T.E., van Ginkel, C., Botha, A.M., and Ashton, P.J. (2008). The use of remote sensing and molecular markers as early warning indicators of the development of cyanobacterial hyperscum crust and microcystinproducing genotypes in the hypertrophic Lake Hartebeespoort, South Africa. Pretoria Counc. Sci. Ind. Res. CSIR.
59. Masango, M., Myburgh, J., Botha, C., Labuschagne, L., and Naicker, D. (2008). A comparison of in vivo and in vitro assays to assess the toxicity of algal blooms. Water Res. 42, 3241–3248.
60. Nchabeleng, T., Cheng, P., Oberholster, P.J., Botha, A.-M., Smit, W.J., and Luus-Powell, W.J. (2014). Microcystin-LR equivalent concentrations in fish tissue during a postbloom Microcystis exposure in Loskop Dam, South Africa. Afr. J. Aquat. Sci. 39, 459–466.
61. Ndebele, M.R., and Magadza, C.H.D. (2006). The occurrence of microcystin-LR in Lake Chivero, Zimbabwe. Lakes Reserv. Sci. Policy Manag. Sustain. Use 11, 57–62.
https://blog.sciencenet.cn/blog-1475614-1272647.html
上一篇:视觉系统的演化之旅——视觉器官、光感受器及视觉分子
下一篇:新理论探索│谁是蛋白质和核酸失踪的“媒人”?